Intraspecific variation in Gyrodactylus mediotorus and G. crysoleucas (Gyrodactylidae) from Nearctic shiners (Leuciscidae): evidence for ongoing speciation, host-switching, and parasite translocation
Chahrazed Rahmouni, Mária Seifertová, Megan G. Bean, Andrea Šimková

TL;DR
This study explores the variation in two Gyrodactylus parasite species infecting fish in Texas, revealing evidence of speciation, host-switching, and parasite translocation.
Contribution
The study provides new evidence for ongoing speciation and host-switching in Gyrodactylus parasites associated with Nearctic fish.
Findings
Gyrodactylus crysoleucas and G. mediotorus were found on new fish hosts in Texas, indicating host-switching.
Genetic divergence in G. mediotorus suggests geographical subgrouping linked to speciation.
Hydrographic barriers and Pleistocene glaciations may have driven the speciation of Gyrodactylus parasites.
Abstract
A parasitological investigation of Cyprinella venusta and Notropis cf. stramineus sampled in Texas, USA, in the Guadalupe River, revealed the presence of Gyrodactylus crysoleucas Mizelle and Kritsky, 1967 on C. venusta, and Gyrodactylus mediotorus King, Marcogliese, Forest, McLaughlin & Bentzen, 2013 on both fish species. This represents new leuscicid fish hosts and locality records for these two gyrodactylids. Gyrodactylus crysoleucas previously identified from both non-native Californian Notemigonus crysoleucas and from farmed stocks in Minnesota demonstrated intraspecific variability in terms of morphology and genetics as a local adaptation associated with isolation by distance. Results further confirmed G. crysoleucas as alien in the western USA and suggested host-switching involving C. venusta and N. crysoleucas. Conservative morphology and genetics on the part of G. mediotorus…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
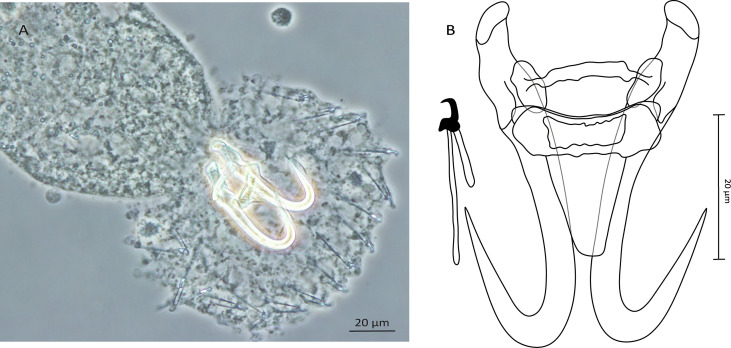 Figure 1
Figure 1 Figure 1
Figure 1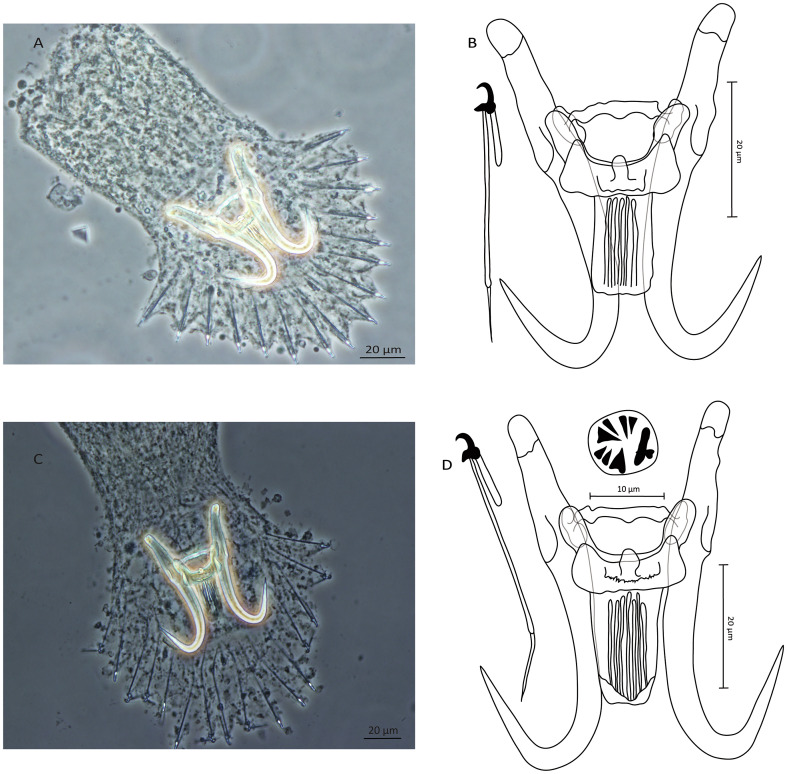 Figure 2
Figure 2 Figure 2
Figure 2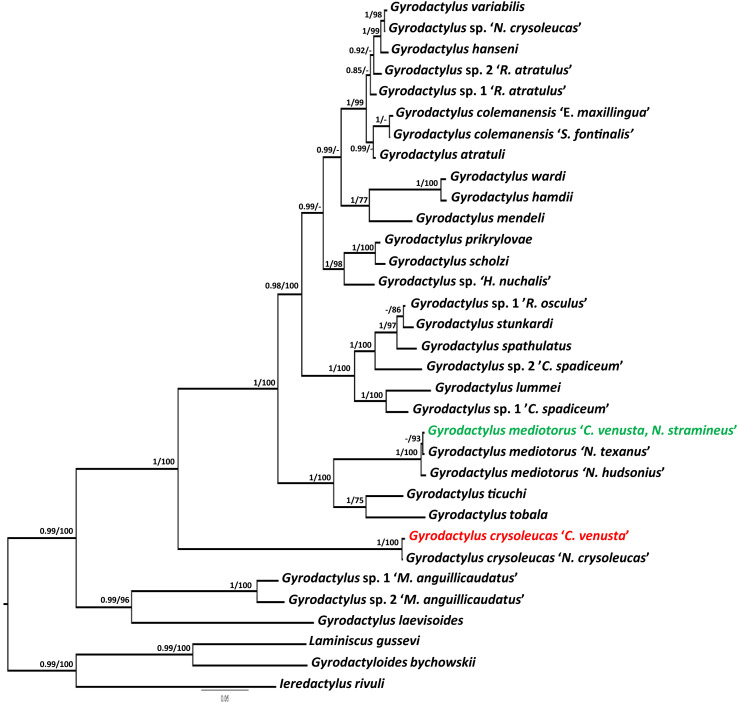 Figure 3
Figure 3 Figure 3
Figure 3- —Ministry of Education, Youth and Sports of the Czech Republic
- —U.S. Fish and Wildlife Service10.13039/100000202
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsParasite Biology and Host Interactions · Bird parasitology and diseases · Genetic diversity and population structure
Introduction
The taxonomic identification of parasitic organisms is an essential task in biodiversity assessment, restoration ecology, and conservation biology, and is crucial for a better understanding of host biogeography and parasite evolution. Generally speaking, nominal species are distinct morphological and genetic clusters separated from other taxa by speciation or separately evolving metapopulation lineages [80, 94]. The evolutionary processes in monogeneans, a highly host-specific ectoparasitic group [85, 105], are known to be driven by new host colonization, ecological adaptation, and host speciation [46, 106, 111]. However, the taxonomy of monogeneans is challenging due to the occurrence of cryptic species characterized by only a few distinguishing traits. Such species, however, exhibit divergent genetic characteristics. The integration of DNA sequences into classification has enabled the recognition of more cryptic monogenean species [7, 34, 83].
Cryptic diversity in monogeneans was previously detected using nuclear markers in gill-specific Dactylogyrus spp. (Dactylogyridae) from native northwest African Luciobarbus spp. (Cyprinidae) [83] despite the generally known presence of relevant morphological characters to discriminate among Dactylogyrus spp. The genus Gyrodactylus von Nordmann, 1832 (Gyrodactylidae) is certainly more taxonomically challenging than Dactylogyrus and its systematics represents a demanding task [58, 78]. As one of the most diverse and widespread taxa within Monogenea [1, 2, 15, 36, 78], Gyrodactylus spp. are known for their highly conservative morphology, making the use of molecular data essential for accurate identification [42, 60, 81, 82]. In contrast to Dactylogyrus, the morphology of each of the sclerotized marginal hooks, anchors (hamuli), transverse bars, and the male copulatory organ (MCO), alone, can be insufficient for recognising a species of Gyrodactylus. Thus, a pairwise comparison of the DNA sequences of various nuclear genes, mainly those of the most informative internal transcribed spacer (ITS rDNA), is required [75,113]. Cryptic Gyrodactylus lineages were reported in several freshwater systems on the basis of sequences of the ITS rDNA, including, for instance, Gyrodactylus rugiensis Gläser, 1974 and Gyrodactylus rugiensoides Huyse and Volckaert, 2002 from European gobiids with a genetic divergence varying between 1.5% and 1.8% [42], and the widespread Gyrodactylus cichlidarum Paperna, 1968 and Gyrodactylus ulinganisus García-Vásquez, Hansen, Christison, Bron and Shinn, 2011, both parasitizing African cichlids and a range of other teleost hosts worldwide, with a genetic divergence reaching 5.2% [31]. Distinct genetic markers like the cytochrome oxidase subunit II (COII) revealed up to 17.2% intraspecific genetic divergence in the case of Gyrodactylus bullatarudis Turnbull, 1956, Gyrodactylus poeciliae Harris and Cable, 2000 and Gyrodactylus turnbulli Harris, 1986, all known to parasitize South American and Caribbean Poecilia guppies [107]. Moreover, cryptic diversity in Gyrodactylus was also observed in non-native (introduced) fish populations of European catfishes [73]. In the Nearctic region, Rahmouni et al. [82] identified complex speciation and diversification processes in Gyrodactylus communities of native Leuciscidae (Cypriniformes) and hypothesized recent/ongoing gene flow. In this case, however, Gyrodactylus huyseae Rahmouni, Seifertová and Šimková, 2023 parasitized two evolutionarily closely-related leuciscid hosts: the striped and spottail shiners, Luxilus chrysocephalus Rafinesque, 1820 and Notropis hudsonius (Clinton, 1824), respectively (the latter species being previously a member of Luxilus (Mayden 1989)), both potentially occurring in an overlapped distributional range [74]. Most recently, the Nearctic Gyrodactylus mediotorus King, Marcogliese, Forest, McLaughlin and Bentzen, 2013, a species originally described from the spottail shiner, N. hudsonius, from Canada, was identified on the weed shiner, Notropis texanus (Girard, 1856), in Wisconsin – however, with only a slight genetic divergence [56]. Molecular phylogeny has revealed host-switching in Gyrodactylus, which is now considered a common speciation scenario in these parasites due to hyperviviparity and a monoxenous life cycle involving transmission by fertile adults [5, 9]. Host-switching scenarios in Gyrodactylus were thought to be less likely due to the phylogenetic dissimilarity between native and new (exotic) fish hosts [27]. Hypothetically, once a gyrodactylid has colonized a suitable natural host population, one would expect a high probability of speciation. From an evolutionary perspective, such sympatric speciation can occur even on phylogenetically distant hosts [6]. While sympatric speciation by host-switch on closely related hosts was evidenced in Gyrodactylus spp. parasitizing European marine gobiids [41, 42], speciation by geographic isolation (allopatric speciation), host-switch, and instant isolation by host specificity were all revealed in Gyrodactylus from European salmonids [63]. At a larger scale, Boeger et al. [3] emphasized the importance of adaptive radiation and nonvicariant processes in the historical diversification of viviparous gyrodactylids. With respect to the Nearctic region, Rahmouni et al. [82], using both 18S rDNA and ITS markers and morphological features of the attachment organ (haptor), reported potential host-switching for the generalist Gyrodactylus hanseni Rahmouni, Seifertová and Šimková, 2023 found on unrelated hosts: L. chrysocephalus and the creek chub Semotilus atromaculatus (Mitchill, 1818). In their study [82], Gyrodactylus colemanensis Mizelle and Kritsky, 1967 was reported for the first time on a wild-living leuciscid, the cutlip minnow Exoglossum maxillingua (Lesueur, 1817), whereas all previous records mostly involved non-native salmonids introduced to North America mainly for farming purposes [12, 14, 18, 33, 65, 104, 109]. To a lesser degree, Nearctic Gyrodactylus that have been introduced to Europe have been shown to accidently infect native freshwater fish [72]. Further, the ornamental (aquarium) fish trade in exotic hosts is one of the main pathways for the co-introduction of non-native monogeneans, leading to parasite spillover towards native fish fauna. This type of invasive scenario was observed in exotic poeciliid fishes (Cyprinodontiformes) native to southeast Asia [47] and to the neotropics [43, 53].
During our recent survey of fish parasites in Texas, we studied a set of monogeneans collected from two shiner species naturally occurring in southcentral USA watersheds, the blacktail and sand shiners, Cyprinella venusta Girard, 1856 and Notropis cf. stramineus (Cope, 1865), respectively. Nearctic minnows of the shiner clade represent one of the most taxonomically complex clades of Cypriniformes due to conserved morphologies in numerous taxa [96]. Taxonomically, shiners and related minnows have been classified as members of the subfamily Pogonichthyinae since the taxonomic revision by Schönhuth et al. [93] and Tan and Armbruster [100], who elevated subfamilies to family rank in cyprinoids. Notropis and many taxa formally placed in this genus have long had an ambiguous status and this genus was considered a taxonomic repository for small, silvery shiners of uncertain placement (see [30]). Still, over 320 nominal species of Nearctic members of Notropis are listed in the Catalog of Fishes [29]. Cyprinella, like Luxilus, Lythrurus, and Pteronotropis, were considered subgenera within Notropis until Mayden [61] elevated them into different genera and relocated species. Cyprinella venusta Girard, 1856 is a small freshwater shiner native to the USA with a wide distributional range across the country [30, 74]. The species has also been introduced into the Pecos River near Pandale, Texas [39]. The blacktail shiner prefers flowing waters, creeks, and rocky pools and runs, and areas with little vegetation and gravelly bottoms [30]. Likewise, Notropis stramineus (Cope, 1865) is a small shiner native to the central part of the USA, southern Canada, and Mexico [74]. In the USA, it is found sporadically on the Edward Plateau, in the Big Bend region of the Rio Grande, and along the Red River [39], and inhabits sand and gravel runs, pools of creeks, and small to large watersheds [30].
From a species-richness perspective, parasitological investigations of Notropis spp. and Cyprinella spp., including all previous fish members of these genera, have revealed a total of 14 Gyrodactylus spp. [48, 49, 56, 62, 82, 86–89]. While most of the known Gyrodactylus spp. parasitizing representatives of the genera Cyprinella, Luxilus, Lythrurus, and Notropis exhibit strict host specificity by infecting a single host species, there are three Gyrodactylus spp. with more than a single recorded host. They are Gyrodactylus baeacanthus Wellborn and Rogers, 1967 from C. venusta [103] and from the comely shiner Notropis amoenus (Abbott, 1874) [52], Gyrodactylus lythruri Rogers, 1975 from the blacktip shiner Lythrurus bellus (Snelson, 1972), and from the pretty shiner congener Lythrurus bellus (Hay, 1881) [88], and finally G. mediotorus found on N. hudsonius and N. texanus [48, 56]. Furthermore, Gyrodactylus atratuli Putz and Hoffman, 1963, known from a range of leuciscid dace species of the genera Margariscus and Rhinichthys [35, 79, 82], was also found to parasitize the spotfin shiner Cyprinella spiloptera (Cope, 1867) [49].
The goal of the present study was first to perform a morphological and genetic study of Gyrodactylus specimens collected from C. venusta and N. stramineus in Texas. Previously, Gyrodactylus spp. had never been documented for N. stramineus. We investigated the degree of similarity/dissimilarity to known Gyrodactylus spp. according to relevant morphological characters of the haptor, mainly, and genetic markers. Further, we summarized the data on the host specificity of the Gyrodactylus spp. studied and tried to infer their historical transmissions in the Nearctic region.
Material and methods
Ethics and permits
Fish were collected under Texas Parks and Wildlife’s state employee biologist permit for Megan Bean.
Shiner fish sampling and genetic identification
A total of 34 shiner specimens of C. venusta and 14 specimens N. stramineus were collected in Texas (USA) on June 5, 2023, in West Mud Creek, Neches River, and on May 30, 2023, in the Guadalupe River (30.073490, −90.140196), near Kerrville, Texas, respectively. Fish specimens were identified in situ by local co-workers (listed in acknowledgements). We further performed molecular analysis on the samples and obtained sequence data from the partial cytochrome b (cyt-b) mitochondrial gene to confirm the identity of the investigated shiners. Mitochondrial DNA of host species was isolated from fin clips preserved in 96% ethanol using a DNeasy^®^ Blood and Tissue Kit (QIAGEN, Hilden, Germany), following the manufacturer’s instructions. The Cyt-b gene was amplified using the forward primer GluF (5′-AACCACCGTTGTATTCAACTACAA-3′) and reverse primer ThrR (5′-ACCTCCGATCTTCGGATTACAAGACCG-3′) [57]. PCR reactions consisted of 1 U of Taq DNA polymerase (Thermo Fisher Scientific, Waltham, MA, USA), 1 × PCR buffer, 1.5 mM MgCl_2_, 0.4 mM of each dNTP, 0.4 μM of each primer, and an aliquot of 30 ng (1 μL) of genomic DNA in a total volume of 25 μL. PCR was carried out in a Mastercycler ep gradient S (Eppendorf AG, Hamburg, Germany) with the following steps: 2 min at 94 °C followed by 39 cycles of 45 s at 92 °C, 90 s at 48 °C, and 105 s at 72 °C, and 7 min of final elongation at 72 °C. The PCR product was purified by EPPiC Fast (A&A Biotechnology, Gdynia, Poland) and was sequenced directly in both directions using the same primers as in the amplification reaction. The initial amplification was carried out using a BigDye^®^ Terminator v3.1 Cycle Sequencing Kit (Applied Biosystems by Thermo Fisher Scientific) and an Applied Biosystems 3130 Genetic Analyzer (Applied Biosystems). Raw nucleotide sequences were edited using Sequencher software v. 5.0 (Gene Codes, Ann Arbor, MI, USA) and aligned using ClustalW [101] as implemented in MEGA v. 11 [99]. The identification of shiners based on a sequence similarity approach was carried out using the Basic Local Alignment Search Tool (https://blast.ncbi.nlm.nih.gov/Blast.cgi: blastn, default settings). Newly generated sequences for the cypriniform species were deposited in GenBank under the accession numbers (accession numbers indicated in Table 1). In this study, shiner fish host nomenclature follows Froese and Pauly [30]. Below, we refer to the sand shiner as N. cf. stramineus (Guadalupe River) as the species is part of a species complex in southcentral USA currently being described (Kevin W. Conway, unpublished data).
Table 1. List of gyrodactylid species included in the phylogenetic analyses performed based on sequences of the ITS region. Monogenean species are grouped by host species and family, and geographical locality. Species used to root the phylogenetic tree are marked by “”. Newly generated sequences are indicated in bold. Fish host nomenclature follows Fricke et al. [29] and FishBase [30].Gyrodactylid speciesHost speciesHost FamilyLocalityITS regions**Gyrodactyloides bychowskii Salmo salar SalmonidaeScotland AJ249348
Gyrodactylus atratuli
Rhinichthys cataractae LeuciscidaeCanada OR270008
Gyrodactylus colemanensis
Exoglossum maxillingua LeuciscidaeUSA OR270009
Gyrodactylus colemanensis
Salvelinus fontinalis SalmonidaeCanada JF836142
** Gyrodactylus crysoleucas **
** Cyprinella venusta **
Leuciscidae
USA
[ PP309998 ](http://www.ncbi.nlm.nih.gov/nuccore/PP309998)
Gyrodactylus crysoleucas
Notemigonus crysoleucas LeuciscidaeUSA KT149287
Gyrodactylus hamdii
Catostomus commersonii CatostomidaeUSA OR269996
Gyrodactylus hanseni
Luxilus chrysocephalus LeuciscidaeUSA OR269998
Gyrodactylus laevisoides
Phoxinus eos LeuciscidaeCanada KF263527
Gyrodactylus lummei
Campostoma spadiceum LeuciscidaeUSA OR270003
Gyrodactylus mediotorus ‘C. venusta, N. stramineus’
** Cyprinella venusta Notropis stramineus **
Leuciscidae
USA
[ PP309999 ](http://www.ncbi.nlm.nih.gov/nuccore/PP309999) Gyrodactylus mediotorus ‘N. hudsonius’ Notropis hudsonius LeuciscidaeCanada KF178301 Gyrodactylus mediotorus ‘Notropis texanus’ Notropis texanus LeuciscidaeUSA MW666182
Gyrodactylus mendeli
Nocomis biguttatus LeuciscidaeUSA OR270004
Gyrodactylus prikrylovae
Pimephales promelas LeuciscidaeUSA OR270005
Gyrodactylus scholzi
Pimephales promelas LeuciscidaeUSA OR270007 Gyrodactylus sp. 1 ‘Misgurnus anguillicaudatus’ Misgurnus anguillicaudatus CobitidaeUSA MH667465 Gyrodactylus sp. 2 ‘Misgurnus anguillicaudatus’ Misgurnus anguillicaudatus CobitidaeUSA MH667466 Gyrodactylus sp. ‘R. osculus’ Rhinichthys osculus LeuciscidaeUSA AY099508 Gyrodactylus sp. ‘H. nuchalis’ Hybognathus nuchalis LeuciscidaeUSA OR270019 Gyrodactylus sp. ‘N. crysoleucas’ Notemigonus crysoleucas LeuciscidaeUSA KT149288 Gyrodactylus sp. 1 ‘R. atratulus’ Rhinichthys atratulus LeuciscidaeUSA OR270020 Gyrodactylus sp. 1 ‘C. spadiceum’ Campostoma spadiceum LeuciscidaeUSA OR270016 Gyrodactylus sp. 2 ‘R. atratulus’ Rhinichthys atratulus LeuciscidaeUSA OR270021 Gyrodactylus sp. 2 ‘C. spadiceum’ Campostoma spadiceum LeuciscidaeUSA OR270017
Gyrodactylus spathulatus
Catostomus commersonii CatostomidaeUSA OR270011
Gyrodactylus stunkardi
Rhinichthys cataractae LeuciscidaeUSA OR270012
Gyrodactylus ticuchi
Notropis moralesi LeuciscidaeMexico MT879676
Gyrodactylus tobala
Notropis imeldae LeuciscidaeMexico MT879671
Gyrodactylus variabilis
Notemigonus crysoleucas LeuciscidaeCanada OR270014
Gyrodactylus wardi
Catostomus catostomus CatostomidaeCanada OR270015 *Ieredactylus rivuli Rivulus hartii RivulidaeTrinidad HQ738514 *Laminiscus gussevi Mallotus villosus OsmeridaeIceland HF548678
Collection and morphological characterization of Gyrodactylus
Once at the laboratory, the external body surfaces, fins, and gills of cyprinid hosts were checked for the presence of gyrodactylid parasites using an MST130 stereoscopic microscope. When present, parasite specimens were removed using surgical needles and mounted on slides with a mixture of glycerine and ammonium picrate (GAP) [59]. Monogenean specimens were studied morphologically considering a total of 23 morphological characters, of which four, seven, and one corresponded to the hamuli (also termed anchors), and the ventral and dorsal bars, respectively, and eight and two characters corresponded to the marginal hooks and male copulatory organ (MCO), respectively. The terminology for the haptoral sclerites and the method of measurements follow those of Malmberg [58], Pugachev et al. [78], and Kritsky and Thatcher [50]. Gyrodactylus species were identified using original descriptions available thus far (see the result sections for references). Measurements (given in micrometers as the mean followed by the range and the number of measurements (n) in parentheses) and photographs were taken using an Olympus BX51 phase-contrast microscope and Olympus Stream Image Analysis v. 1.9.3 software (Olympus, Tokyo, Japan). Drawings of the haptoral sclerotized parts were made on flattened specimens using an Olympus BX51 microscope equipped with a drawing tube and edited with a graphic tablet compatible with Adobe Illustrator CS6 v. 16.0.0 and Adobe Photoshop v. 13.0 (Adobe Systems Inc., San Jose, CA, USA). Prevalence and intensity of infection were calculated according to Bush et al. [8]. Voucher material was deposited in the Helminthological Collection of the Institute of Parasitology, the Biology Centre of the Academy of Sciences of the Czech Republic, České Budějovice (IPCAS) under the accession numbers (see Results section).
DNA amplification, sequencing, and phylogeny of Gyrodactylus spp.
Gyrodactylus specimens were subjected to DNA amplification and sequencing. Specimens stored in 96% ethanol were dried using an Eppendorf 5301 Concentrator. Total genomic DNA was extracted using the DNeasy^®^ Blood and Tissue Kit (QIAGEN) following the protocol for the purification of total DNA from animal tissues. Two nuclear ribosomal DNA markers suitable for the differentiation of Gyrodactylus spp. were used (for instance, see [10,33,60,75,113]). A fragment spanning ITS1, 5.8S and ITS2 (ITS regions) was amplified using forward primer ITS1F (5′-GTTTCCGTAGGTGAACCT-3′) [90], complementary to the sequence at the 3′ end of the 18S rRNA gene, and reverse primer ITS2 (5′-TCCTCCGCTTAGTGATA- 3′) [17], complementary to the sequence at the 5′ end of the 28S rRNA gene [17]. A partial fragment of 18S rDNA containing the V4 region, which exhibits intraspecific variation in Gyrodactylus [16, 60], was amplified using the primer pairs PBS18SF (5′-CGCGCAACTTACCCACTCTC-3′) and PBS18SR (5′-ATTCCATGCAAGACTTTTCAGGC-3′) [13]. Polymerase chain reactions (PCRs) for the 18S rDNA gene and ITS region were performed in a final volume of 30 μL, containing 1xPCR buffer, 1.5 mM MgCl_2_, 200 μM of each dNTP, 0.5 μM of each primer, 1 U of Taq DNA Polymerase (Thermo Scientific) and 5 μL of template DNA. The PCRs were carried out in the Mastercycler ep gradient S (Eppendorf) using the following steps: i) ITS regions: an initial denaturation at 96 °C for 3 min, followed by 39 cycles of denaturation at 95 °C for 50 s, annealing at 52 °C for 50 s, and an extension at 72 °C for 50 s, and a final elongation at 72 °C for 7 min; and ii) 18S region: an initial denaturation at 95 °C for 3 min, followed by 39 cycles of denaturation at 94 °C for 1 min, annealing at 54 °C for 45 s, and an extension at 72 °C for 1 min 30 s, and a final elongation at 72 °C for 7 min. PCR products were electrophoresed on 1.5% agarose gels stained with Good View (SBS Genetech, Bratislava, Slovakia) and then purified using EPPiC Fast (A&A Biotechnology, Gdynia, Poland), following the manufacturer’s protocol. The purified PCR products were sequenced directly in both directions using the PCR primers. Sanger sequencing was carried out using a BigDye^®^ Terminator v3.1 Cycle Sequencing Kit (Applied Biosystems) and an Applied Biosystems 3130 Genetic Analyzer. Newly-generated DNA sequences were assembled and edited using Sequencher software v. 5.0 (Gene Codes, Ann Arbor, MI, USA) and aligned using ClustalW [101] as implemented in MEGA v. 11 [99]. Sequences were further checked by the nBLAST Search Tool (https://blast.ncbi.nlm.nih.gov/Blast.cgi: blastn, default settings, access date: 11/09/2023) to assess any similarity to available congeners, then deposited in GenBank under accession numbers indicated in the Results section. Genetic divergences were estimated using uncorrected p-distances in MEGA v. 11 [99].
A phylogenetic tree was reconstructed based on the newly generated ITS sequences of G. crysoleucas and G. mediotorus, together with 31 DNA sequences representing 29 Gyrodactylus spp. retrieved from the GenBank database. The dataset included Gyrodactyloides bychowskii Albova, 1948, Ieredactylus rivuli Schelkle et al., 2011, and Laminiscus gussevi (Bychowsky and Polyansky, 1953) Pálsson and Beverley-Burton, 1983 as the outgroup following Rahmouni et al. [81] (Table 1). Nucleotide sequences were aligned by multiple alignments using MAFFT v7.505 [45] and trimmed using trimAl v1.2rev57 [11] through plugins installed in PhyloSuite v1.2.3 [108, 110]. The plugin of ModelFinder v2.2.0 [44] in PhyloSuite v1.2.3 [108, 110] was used to determine the best-fit substitution model for the dataset. A final alignment of 33 ITS sequences composed of 927 bp was used to infer the phylogenetic relationships using Maximum Likelihood (ML) and Bayesian Inference (BI). ModelFinder v2.2.0 [44] indicated GTR+F+G4 as the best-fitting evolutionary models for ML analysis based on the corrected Akaike Information Criterion (AICc) [40, 97]. ML trees were inferred using IQ-TREE v1.5.5 [69] based on the selected model employing a sub-tree pruning and re-grafting (SPR) branch-swapping algorithm. The branch support (bootstrap support, BS) was estimated using ultrafast bootstrap approximation [64] with 1,000 replicates. BI analysis was performed using MrBayes v3.2.1 [91] and applying the GTR+I+G evolutionary model with two independent Markov Chain Monte Carlo (MCMC) simulations (six chains, 10^6^ generations, sampling frequency 100, 25% burn-in to obtain the consensus tree and posterior probability values (PP)). Chain stationarity and parameter convergence were assessed in TRACER v1.7.1 [84], with effective samples sizes (ESS) always >200 for all parameters, and via the average standard deviation of split frequencies (always well below 0.01), and post burn-in trees were summarized in a 50% majority rule consensus tree. The resulting ML and BI trees were visualized in FigTree v1.4.4 (http://tree.bio.ed.ac.uk) and manually edited on Photoshop v13.0.
Results
Morphological characterization considering the haptoral hard parts indicated Gyrodactylus specimens studied herein to be highly reminiscent of already known species. Based on this statement together with DNA sequences, we confirmed the presence of G. crysoleucas Mizelle and Kritsky, 1967, restricted so far to either non-native [65] or captive golden shiner, Notemigonus crysoleucas (Mitchill, 1817) [55], and the well-known G. mediotorus of N. hudsonius [48] and N. texanus [56].
Morphological characterization
Gyrodactylidae Cobbold, 1864
Gyrodactylus Nordmann, 1832
Gyrodactylus crysoleucas Mizelle and Kritsky, 1967 (Figures 1A–1B)
Previous records: golden shiner, N. crysoleucas, Rooney Pond, 17 miles east of Sacramento, California, USA [65]; captive N. crysoleucas, private baitfish, Minnesota [55], both in the USA.
Figure 1. Photomicrographs (A) and line drawings (B) of Gyrodactylus crysoleucas from the blacktail shiner, Cyprinella venusta.
Present study: blacktail shiner, C. venusta, West Mud Creek, Neches River, Texas, USA
Site of infection: fins
Prevalence and intensity of infection: 2.9%, a single infected host out of 34 investigated, two Gyrodactylus specimen on a single infected host.
Voucher: IPCAS M-795
Host GenBank accession number: cyt-b: PP314044–PP314045
Parasite GenBank accession numbers: 18S rDNA: PP309996; ITS: PP309998
Morphology: Haptor subcircular, anchor base with folds, tips curved inward, total length 51.6 (51.2–51.9; n = 2); shaft slightly bowed, length 39.6 (39–40.3; n = 2); point curved and elongate, length 22.9 (22.5–23.4; n = 2); root relatively short, tapered, length 14.3 (13.1–15.5; n = 2). Ventral bar with short, blunt lateral processes extending out of bar, total length 23.1 (23–23.2; n = 2), total width 22.9 (22.7–23.1; n = 2), lateral processes length 2 (1.5–2.5; n = 2), distance between tips 22.4 (21.5–23.3; n = 2), median width 5.3 (5.2–5.3; n = 2), membrane (shield) almost trapezoid tapering posteriorly and extending almost 1/2 of length of anchor shaft, no striations or ridges were observed, length 15 (13.8–16.2; n = 2), width at the insertion 12.8 (12.3–13.4; n = 4). Dorsal bar straight with projections near each end, attenuated ends inserted into terminal plates, total length 21.9 (19.4–24.4; n = 2), width at midpoint 2.5 (2.2–2.7; n = 2). Marginal hooks total length 25.6 (25.1–26.1; n = 2); sickle foot noticeable with downward globose heel, prominent triangular toe, conspicuous shelf; sickle proper almost as thick as toe base, shaft length 5.2 (4.9–5.5; n = 2); sickle length to shaft attachment 3.5 (3.4–3.5; n = 2); sickle proximal width 2.9 (2.6–3.2; n = 2); sickle distal width 3.7 (3.6–3.8; n = 2); point relatively thin and slightly curved, length 1.5 (1.3–1.8; n = 2); filament loop extending about 1/2 of handle length, length 8.9 (8.3–9.5; n = 2); handle length 20.8 (19.9–21.6; n = 2). MCO not found.
Considering the original study by Mizelle and Kritsky [65], our specimens presented relatively (i) shorter anchors (51.2–51.9 μm in this study vs. 55–61 μm in the original description), and (ii) a shorter ventral bar (23–23.2 μm in this study vs. 25–32 μm in the original description). The shape of the marginal hooks was similar with a downward heel and prominent finger-like toe and shelf, but with a slightly thinner shaft of the sickle proper in the case of our specimens. Photographs published by Leis et al. [55] provided more details about the morphology of the haptoral sclerotized structures of G. crysoleucas and indicated that our specimens parasitizing C. venusta sampled in its natural southcentral range and those found on cultured N. crysoleucas in Minnesota were of a similar shape, and that measurements of the hard parts mostly overlapped.
Gyrodactylus mediotorus King, Marcogliese, Forest, McLaughlin & Bentzen, 2013 (Figures 2A–2D)
Previous records: spottail shiner, N. hudsonius (type-host), Îles de la Paix, St. Louis Lake (type-locality) and Îles Vert, St. Lawrence Lake, both in Quebec, Canada [48]; weed shiner, N. texanus, Lake Onalaska, Upper Mississippi River, Wisconsin, USA [56].
Figure 2. Photomicrographs (A–C) and line drawings (B–D) of Gyrodactylus mediotorus King, Marcogliese, Forest, McLaughlin and Bentzen, 2013 from the blacktail shiner, Cyprinella venusta (A–B) and from the sand shiner, Notropis cf. stramineus (Guadalupe River) (C–D).
Present study: blacktail shiner, C. venusta, West Mud Creek, Neches River (Figure 2A–2B), and sand shiner, N. cf. stramineus (Guadalupe River), Guadalupe River (Figure 2C–2D), both in Texas, USA.
Site of infection: fins
Prevalence and intensity of infection: for C. venusta, 5.9%, two infected hosts out of 34 investigated, a single Gyrodactylus specimen on each infected host. For N. cf. stramineus (Guadalupe River), 14.3%, two infected hosts out of 14 investigated, from 1 to 3 Gyrodactylus specimen per infected host.
Voucher: IPCAS M-794/1-2
Host GenBank accession numbers: cyt-b: PP314044–PP314045 for C. venusta, PP314046 for N. stramineus
Parasite GenBank accession numbers: 18S rDNA: PP309997; ITS: PP309999
Morphology: Haptor subcircular, anchor base with folds, tips curved outward, total length 59.4 (55.9–62.6; n = 8); shaft slightly bowed, length 42.9 (40–45; n = 8); point curved and elongate, length 21.5 (20.3–22.6; n = 8); root long, length 20.3 (18.3–21.6; n = 8). Ventral bar with prominent blunt lateral processes extending out of bar, total length 33.5 (30.7–36.9; n = 8), total width 22.4 (20.7–24.8; n = 8), lateral processes length 7.8 (5.5–9.5; n = 8), distance between tips 26.1 (23–29.5; n = 8), median part with a noticeable knob, width 5.5 (4.3–6.9; n = 8), membrane (shield) rectangular with fine longitudinal striations, extending almost 1/2 of length of anchor shaft, length 18.1 (15.6–20.4; n = 8), width at the insertion 15.3 (12.3–17.6; n = 8). Dorsal bar straight with projections near each end, attenuated ends inserted into terminal plates, total length 23.7 (21–26.2; n = 8), width at midpoint 2.6 (1.8–2.9; n = 8). Marginal hooks total length 35.6 (31–38.8; n = 8); sickle foot moderate with downward globose heel, prominent triangular straightforward toe, conspicuous shelf; sickle proper almost as thick as toe base, shaft length 4.9 (4.4–5.4; n = 8); sickle length to shaft attachment 3.2 (2.8–3.7; n = 8); sickle proximal width 3.1 (2.6–3.5; n = 8); sickle distal width 3.4 (2.8–4; n = 8); point relatively thin and slightly curved, length 1.3 (1.1–1.7; n = 8); filament loop extending about 1/3 of handle length, length 8.5 (7.3–9.7; n = 8); handle ending in a noticeable posterior filament, length 30.1 (26.2–34; n = 8). MCO observed in a single Gyrodactylus specimen from N. stramineus cf. (Guadalupe River) with a single apical prominent spine and a single row of seven spinelets.
Size and shape of the sclerotized structures of G. mediotorus specimens from southeast populations of each of C. venusta and N. cf. stramineus (Guadalupe River) overlapped. Compared to the type-material [48], noticeable intraspecific variability was observed, mainly in terms of the (i) shorter hamuli (55.9–62.6 μm in this study vs. 65.7–69.7 μm in the original description), and (ii) the shorter ventral bar (30.7–36.9 μm in this study vs. 36.4–41.3 μm in the original description). Although well visible on the photographs, King et al. [48] did not mention the presence of a knob in the median part of the ventral bar or a prominent filament attachment posteriorly to the handle of the marginal hooks. The median knob of the ventral bar was later emphasized by Leis et al. [56] when reporting a variant of G. mediotorus on N. texanus, whereas the additional filament marking the posterior end of the marginal hooks was not highlighted. The specimens of G. mediotorus studied herein can be compared to the so-far-unknown Gyrodactylus sp. “C. venusta” collected recently in Mississippi [82] in having a similar haptoral morphotype, but mainly because of the presence of the ventral bar knob and the filament of the marginal hooks. Yet, considerable variation in the size of the ventral bar is observed (30.7–36.9 μm in this study vs. 20.1 μm in [82]). Likewise, our specimens of G. mediotorus possessed a longer ventral bar than G. mediotorus from N. texanus (30.7–36.9 μm in this study vs. 22 μm in [56]).
Genetics and phylogenetic reconstruction
Gyrodactylus crysoleucas
Two fragments, the first covering the 18S rDNA (442 bp) and the second covering the ITS regions (1,207 bp), were successfully sequenced for two gyrodactylid specimens from Texas, and the newly obtained sequences were found to be identical. nBLAST search indicated G. crysoleucas KT149283 [55] from the farmed golden shiner N. crysoleucas in Minnesota (USA) as an identical match to our specimens according to sequences of 18S rDNA (100% similarity, 100% coverage), while G. crysoleucas KT149287 [55] was recovered as the closest match to the specimens studied herein according to the sequences of the ITS regions (99.17% similarity, 99% coverage, p-distance = 0.3%; 3 bp). In accordance with the morphological identification, our specimens were genetically assigned to G. crysoleucas following the delimitation by Ziȩtara and Lumme [112] and the recent findings of Rahmouni et al. [82]. Sequences of 18S rDNA further indicated Gyrodactylus salmonis (Yin and Sproston, 1948) from the non-native rainbow trout Oncorhynchus mykiss (Walbaum, 1792) from Veracruz (Mexico) (JN230350,[92]) and Washington (USA) (JF836097, [33]) as the closest match to our G. crysoleucas specimens from Texas (for both G. salmonis genetic variants; 99.55% similarity, 100% coverage, p-distance = 0.5%; 2 bp). Phylogenetically, G. crysoleucas parasitizing herein C. venusta from Texas together with its congener parasitizing N. crysoleucas from Minnesota formed a highly supported basal clade (PP = 1, BS = 100) in relation to a large clade of Nearctic Gyrodactylus spp. (Figure 3).
Figure 3. Bayesian inference (BI) phylogram of Gyrodactylus spp. parasitizing Nearctic Cypriniformes based on sequences of the ITS regions (927 bp). Values above branches indicate posterior probabilities (PP) from BI analyses and bootstrap support (BS) from ML. Values below 0.80 (BI) and 70 (ML) are shown as dashes.
Gyrodactylus mediotorus
Two fragments, the first covering the 18S rDNA region (436 bp) and the second covering the ITS region (1,010 bp), were successfully sequenced for two gyrodactylid specimens from each of C. venusta and N. cf. stramineus (Guadalupe River) from Texas. For each gene, no intraspecific variability associated with respective fish hosts was found. Based on sequences of the 18S rDNA region, nBLAST search recovered two variants of G. mediotorus from the farmed spottail shiner N. hudsonius from Quebec (Canada) (KF178302, [48]) and the native weed shiner N. texanus from the Upper Mississippi in Wisconsin (USA) (MW666777, [56]) as identical matches to our specimens (respectively, 100% similarity, 100% coverage; 100% similarity, 94% coverage). Likewise, nBLAST search using the ITS sequences indicated the same variants of G. mediotorus from the USA (MW666182, [56]) and Canada (KF178301, [48]) as the closest matches to our specimens (respectively, 98.91% similarity, 99% coverage; 98.42% similarity, 99% coverage). In accordance with the morphological characterization, the parasite specimens we collected were genetically recognized as G. mediotorus following the delimitation by Ziȩtara and Lumme [116]. Yet, genetic intraspecific variation was found when comparing newly obtained and published sequences of G. mediotorus. P-distances using sequences of the ITS regions approached the limit value (1%) [116] with our specimens of G. mediotorus from C. venusta and N. cf. stramineus (Guadalupe River) from the southcentral basin (Texas) genetically closer to those found on N. texanus from the Upper Mississippi (Wisconsin, USA) (p-distance = 0.2%; 2 bp) than to Gyrodactylus specimens parasitizing N. hudsonius from the northeastern locality (Canada) (p-distance = 0.9%; 9 bp). With high support values (PP = 1, BS = 100), the phylogenetic reconstruction (Figure 3) indicated the monophyly of three variants of G. mediotorus with sister position to the clade of Gyrodactylus ticuchi Pinacho-Pinacho et al., 2021 (MT879676) and Gyrodactylus tobala Pinacho-Pinacho et al., 2021 (MT879671) parasitizing Notropis moralesi de Buen, 1955 and Notropis imeldae Cortés, 1968, respectively, from Mexico [75].
Discussion
The present survey focused on Gyrodactylus spp. hosted by two Nearctic leuciscid shiners, C. venusta and N. cf. stramineus (Guadalupe River) occurring in their native watersheds in Texas [30, 74] Morphological study based on taxonomically important haptoral features combined with DNA sequences of the 18S rDNA and ITS regions revealed the presence of two already-known species – namely, G. crysoleucas and G. mediotorus. We recovered the former species from C. venusta (Figures 1A and 1B), while G. mediotorus was found to parasitize both C. venusta (Figures 2A and 2B) and N. cf. stramineus (Guadalupe River) (Figures 2C and 2D).
Gyrodactylus crysoleucas: a successfully introduced parasite in the western USA
Considering the morphometric data provided by Mizelle and Kritsky [65] when describing G. crysoleucas from non-native N. crysoleucas, intraspecific variability was found in which specimens collected herein from the southcentral location exhibited shorter haptoral parts, mainly the hamuli and ventral bar. Genetically, while published and newly obtained sequences of the 18S rDNA regions of G. crysoleucas were conserved, sequences of the ITS regions demonstrated weak intraspecific variability on the geographical scale. Although genetic data on G. crysoleucas in western freshwaters are missing, the morphological variability between our specimens and the types could be explained by phenotypic plasticity and/or local adaptation in the newly invaded host (habitat) or possibly allopatric isolation. In the Nearctic region, variability in haptoral morphology across distant localities was already documented in G. atratuli Putz and Hoffman, 1963, a species parasitizing a wide range of leuciscid fish hosts [35, 49, 79, 82]. nBLAST search recovered G. crysoleucas and G. salmonis from non-native salmonids with highly conservative 18S rDNA sequences, which is in accordance with the morphotype of their haptoral parts, marked by poorly developed lateral processes of the ventral bar, a short posteriorly-tapering membrane, and marginal hooks with a downward heel and finger-like toe with a prominent shelf [92]. Conservative 18S rDNA sequences were reported in the recently described G. hanseni Rahmouni, Seifertová and Šimková, 2023 parasitizing both the striped shiner Luxilus chrysocephalus Rafinesque, 1820, and the creek chub, Semotilus atromaculatus (Mitchill, 1818), and in other Gyrodactylus species from native leuciscids and cultured salmonids [33, 82].
The present study adds two shiner species to the known hosts of G. crysoleucas across the USA; N. crysoleucas from California [65] and Minnesota [55], and C. venusta studied herein from Texas. This study presents, thus, new host and locality records for G. crysoleucas. To understand the geographical range of distribution of G. crysoleucas, it is necessary to track the historical origin of N. crysoleucas and C. venusta in the collected areas. According to the USGS database [71], both N. crysoleucas and C. venusta are naturally present in Texas and Minnesota, but not in the western part of the USA, including California. In contrast, N. crysoleucas was previously (late 1890s) distributed to multiple water bodies in California as a major bait and forage fish species by the California Fish Commission [26] and, since then, it has quickly spread in the western USA [98]. This is similar to the red shiner Cyprinella lutrensis (Baird and Girard, 1853), native to central North America west of the Mississippi River drainage [30, 74], which was successfully introduced into California’s inland waters [67]. According to Moyle [67], the golden shiner, N. crysoleucas, was introduced to California from more eastern watersheds, which makes G. crysoleucas first described by Mizelle and Kritsky [65] from N. crysoleucas sampled in California an alien parasite in the western part of the USA, co-introduced with golden shiner hosts. This scenario is supported by the fact that G. crysoleucas was found herein on wild C. venusta native to Texas and previously on cultured golden shiners in the far North in Minnesota [55], where they are often harvested from wild sources, which makes the possibility that G. crysoleucas is of western origin less likely. Nevertheless, this statement requires further investigation since the native parasite fauna in freshwater fish is still underexplored in this region. From native C. lutrensis in Midwestern USA (Nebraska), a single species, G. callawayensis Mayes, 1977, was described [62]. This was interestingly reminiscent of G. crysoleucas regarding the morphotype of the ventral bar characterized by poorly developed lateral processes and a short, posteriorly tapering membrane. The twisted anterior part (tips) to the hamuli is present in G. callawayensis [62] but not in G. crysoleucas which discriminates these two species. Since C. lutrensis is non-native in western inland waters, it would be worthwhile to investigate whether the red shiner has co-introduced its native gyrodactylids to Californian freshwaters.
Only a single parasite species, namely, G. baeacanthus, was formally described from C. venusta [103], whereas Rahmouni et al. [82] recently reported the presence of an undescribed species, Gyrodactylus sp. “C. venusta”, highly reminiscent of G. mediotorus isolated herein from C. venusta but also from N. cf. stramineus (Guadalupe River) (see below). In contrast, two species were recognized on N. crysoleucas additionally to G. crysoleucas [55, 65]; they are G. rachelae Price and McMahon, 1967 from the southeast (Tennessee) [76], and G. variabilis Mizelle and Kritsky, 1967 from western [65], northeast, and southcentral USA, as well as from northeast Canada [82]. Interestingly, previous [103] and current records of gyrodactylids from C. venusta were made from southern populations occurring in Georgia, Alabama, and Louisiana watersheds and in Texas, all representing the native distributional range of C. venusta [30, 70, 74]. In this study, morphological and genetic characterizations indicated that C. venusta hosted G. crysoleucas rather than G. baeacanthus. Two hypotheses may explain this pattern. The first one is that southcentral C. venusta is a native host of G. crysoleucas, making further widescale parasitological investigations of shiner hosts necessary to identify the gyrodactylid fauna in Texas. The second hypothesis is that G. crysoleucas is native to N. crysoleucas and infected a non-congeneric host, C. venusta in this case, by host-switching in overlapped habitats. Host-switching scenarios are common in Gyrodactylus as one major mechanism of speciation [72, 111] and have been documented in Nearctic freshwaters [82]. Further sampling of N. crysoleucas for the investigation of Gyrodactylus in Texas as well as across the whole range of its current distribution would provide more support for this hypothesis. Further, the natural host-switching of Gyrodactylus is known to be favorable under conditions of high parasite abundance and population growth [66] or continuous transmission ability [4]. More specific information on the infection rates of G. crysoleucas and on the population density of C. venusta would provide support for the host-switching scenario in southcentral USA.
Gyrodactylus mediotorus illustrating ongoing speciation in the Nearctic
In the present study, G. mediotorus was isolated from the blacktail and sand shiners, C. venusta and N. cf. stramineus (Guadalupe River), respectively, both collected in their natural distribution range in Texas. Gyrodactylus mediotorus was originally described on the spottail shiner, N. hudsonius in Canada [48], and recently identified from the weed shiner, N. texanus, from the Upper Mississippi River in Wisconsin [56]. Therefore, this study presents two new shiner hosts for G. mediotorus and a new locality in the southeast USA. No morphological intraspecific variability of the haptoral hard parts was found in G. mediotorus across the studied shiner hosts. Inversely, the sclerotized structures of G. mediotorus collected from Texas in this study were shorter compared to those in the type-material [48]. Moreover, G. mediotorus appeared to possess typical features previously overlooked – specifically, the knob in the median part of the ventral bar and the prominent filament attachment posterior to the handle of marginal hooks. The filament of the marginal hooks is already known to be present in G. spathulatus Mueller, 1936 restricted to catostomid hosts so far [20–24, 37, 54, 68, 77, 82, 102], and in the generalist G. stunkardi Kritsky and Mizelle, 1968 infecting a range of Nearctic cypriniforms [21, 22, 51, 52, 68, 82, 102]. It was also reported in the new but unidentified Gyrodactylus spp. from the blacknose dace, Rhinichthis atratulus, and C. venusta [82]. Although morphologically similar to each other and occurring in the southcentral part of the USA, the G. mediotorus we studied herein and Gyrodactylus sp. “C. venusta” collected previously in Mississippi most likely belong to two distinct species due to the considerable size variation in the ventral bar [82]. DNA sequences of the ITS regions will certainly clarify the taxonomic status of Gyrodactylus sp. “C. venusta” in the future. Furthermore, our specimens and those of G. mediotorus from N. texanus [56]presented intraspecific variability. This morphological discrepancy can be related to a specific host and/or geographical locality or phenotypic plasticity as previously evidenced in Gyrodactylus communities [19, 72, 82]. Sequences of the 18S rDNA and ITS regions of G. mediotorus were successfully obtained in this study and were fully conserved at the host species level. This could be linked to the common evolutionary history of shiners in the Nearctic region [96]. On the one hand, in terms of host specificity and similar to remarks by Šimková et al. [95], G. mediotorus appears to be an intermediate specialist parasitizing congeneric as well as phylogenetically closely related non-congeneric shiner hosts across the Nearctic region. Alternatively, the presence of G. mediotorus on southcentral populations of C. venusta and N. cf. stramineus (Guadalupe River) could simply be an inheritance from a common ancestor or has resulted from host-switching. The former scenario seems plausible given the evolutionary relatedness between these shiners [96]. The overall Notropis host range associated with G. mediotorus and its phylogenetically closely related G. ticuchi and G. tobala may indicate that host-switching occurred from Notropis spp. to C. venusta rather than the opposite pattern (from C. venusta to Notropis spp.). Host-switching of Gyrodactylus also seems possible given the close phylogenetic relationship between C. venusta and N. cf. stramineus (Guadalupe River) and their occurrence in overlapping ecological niches in Texas. In monogeneans of the genus Lamellodiscus (Diplectanidae) parasitizing sympatric marine sparid fish hosts, for instance, host-parasite associations have been shown to be mostly driven by ecological factors that considerably facilitated host-switching processes [25].
In accordance with the morphological variability observed among all currently available G. mediotorus, i.e., specimens from Canadian N. hudsonius (type-material), from N. texanus of the Upper Mississippi River in Wisconsin, and from both C. venusta and N. cf. stramineus (Guadalupe River) from Texas, genetic divergence was found, with G. mediotorus from Wisconsin and Texas (both USA) being genetically closer to each other than to the Canadian variant. Values of *p-*distances (0.9%) even approached the upper limit value (1%) of intraspecific genetic variation in the ITS regions usually considered to discriminate among Gyrodactylus spp. [41, 112]. Rahmouni et al. [82] questioned the cryptic status of G. huyseae found to parasitize two historically-connected hosts, L. chrysocephalus and N. hudsonius, occurring in overlapping ranges, when the genetic variation in the ITS sequences slightly exceeded the limit value and a single mutation was found in the 18S rDNA sequences. The morphological and genetic divergence of G. mediotorus on the geographical scale evidenced in this study may be explained by the evolutionary history of shiners – particularly, their evolution during the Pleistocene glaciations, which considerably shaped the current distribution of freshwater biotas in North America [28, 38]. Hydrographic barriers favoring separate evolutionary pathways in Nearctic freshwaters could also have been involved in creating the morphological and genetic patterns observed in G. mediotorus populations. Ongoing speciation in G. mediotorus is, thus, most likely given the complex combinations of dispersal and vicariance events that shiner hosts have experienced. The geological, climatic, and biotic factors and circumstances that have promoted such speciation remain unknown, but deeper investigations involving powerful genetic approaches using various markers, ideally both nuclear and mitochondrial, will certainly help illuminate how gyrodactylid communities are evolving and adapting to distinct Nearctic fish hosts.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bakke TA, Cable J, Harris PD. 2007. The biology of gyrodactylid monogeneans: the “Russian-doll killers”. Advances in Parasitology, 64, 161–376.17499102 10.1016/S 0065-308X(06)64003-7 · doi ↗ · pubmed ↗
- 2Bakke TA, Harris PD, Cable J. 2002. Host specificity dynamics: observations on gyrodactylid monogeneans. International Journal for Parasitology, 32, 281–308.11835970 10.1016/s 0020-7519(01)00331-9 · doi ↗ · pubmed ↗
- 3Boeger WA, Kritsky DC, Pie MR. 2003. Context of diversification of the viviparous Gyrodactylidae (Platyhelminthes, Monogenoidea). Zoologica Scripta, 32, 437–448.32336863 10.1046/j.1463-6409.2003.00130.x PMC 7165889 · doi ↗ · pubmed ↗
- 4Boeger WA, Kritsky DC, Pie MR, Engers KB. 2005. Mode of transmission, host switching, and escape from the Red Queen by viviparous gyrodactylids (Monogenoidea). Journal of Parasitology, 91, 1000–1007.16419740 10.1645/GE-515R.1 · doi ↗ · pubmed ↗
- 5Brooks DR, Mc Lennan DA. 1993. Comparative study of adaptive radiations with an example using parasitic flatworms (Platyhelminthes: Cercomeria). American Naturalist, 142, 755–778.10.1086/28557119425955 · doi ↗ · pubmed ↗
- 6Brooks DR. 1979. Testing the context and extent of host-parasite coevolution. Systematic Zoology, 28, 299–307.
- 7Bueno-Silva M, Boeger WA, Pie MR. 2011. Choice matters: incipient speciation in Gyrodactylus corydori (Monogenoidea: Gyrodactylidae). International Journal for Parasitology, 41, 657–667.21310159 10.1016/j.ijpara.2011.01.002 · doi ↗ · pubmed ↗
- 8Bush AO, Lafferty KD, Lotz JM, Shostak AW. 1997. Parasitology meets ecology on its own terms: Margolis et al. revisited. Journal of Parasitology, 83, 575–583.9267395 · pubmed ↗