Orthotrichumcamanchacanum, a remarkable new moss species from Chile (Bryopsida, Orthotrichaceae)

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
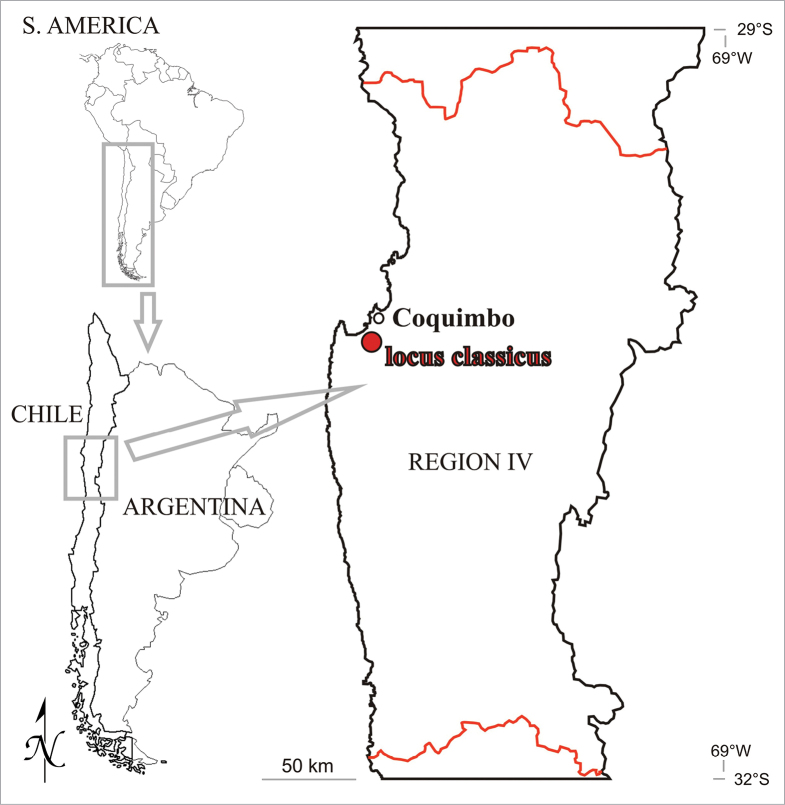 Figure 1
Figure 1 Figure 2
Figure 2 Figure 3
Figure 3 Figure 4
Figure 4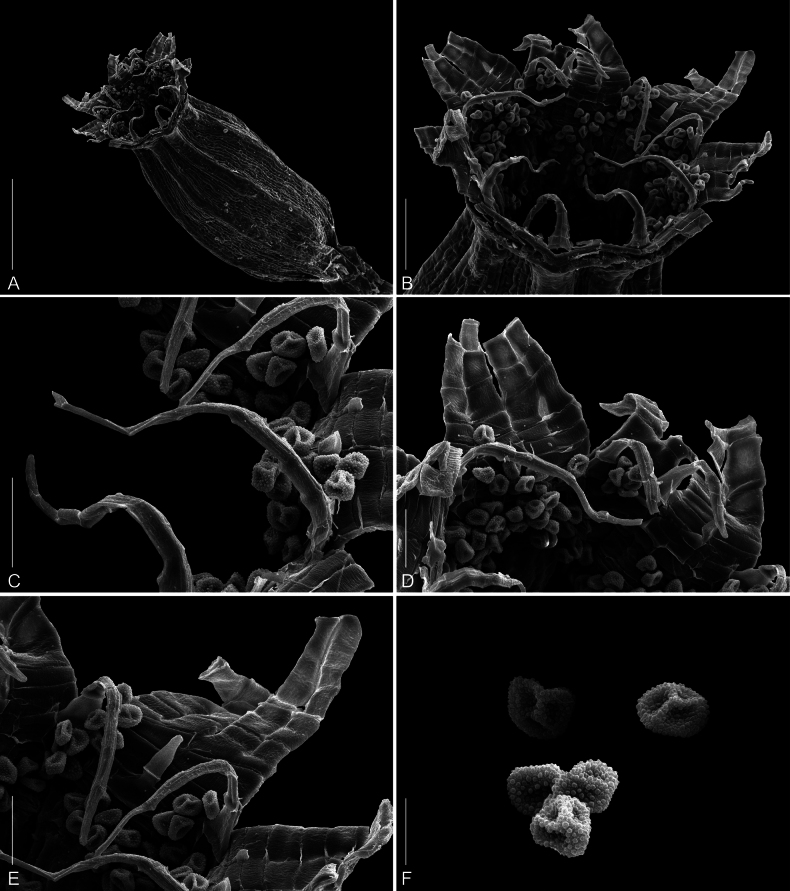 Figure 5
Figure 5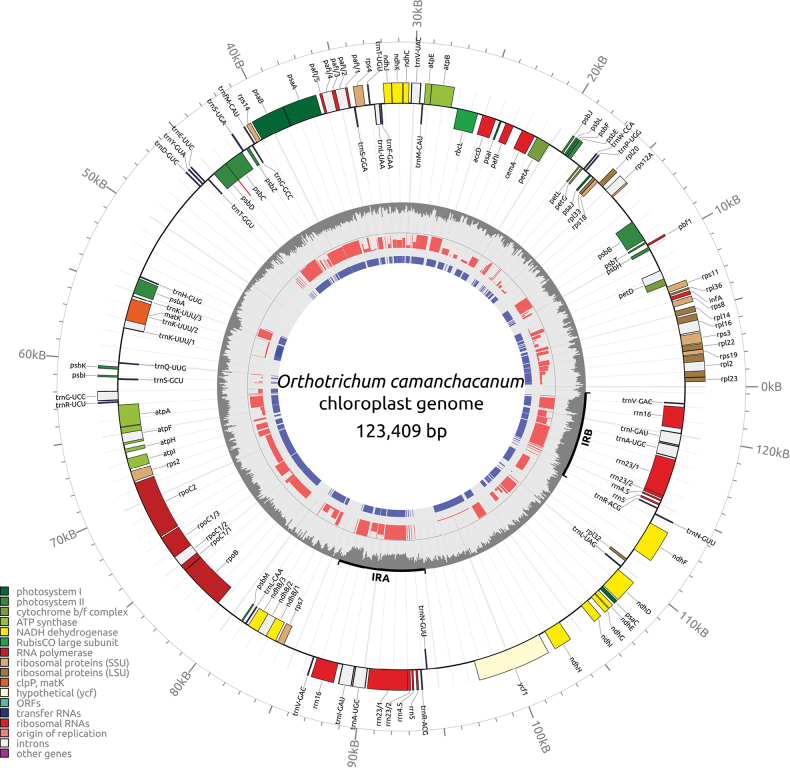 Figure 6
Figure 6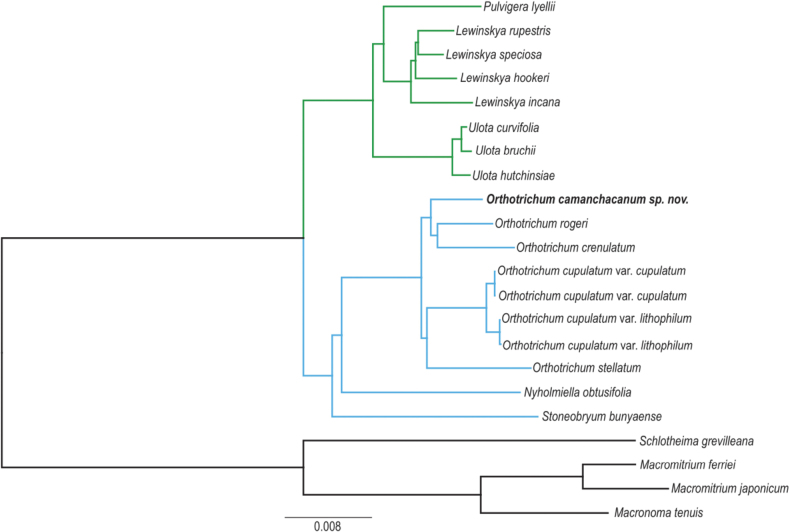 Figure 7
Figure 7|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| ♢ | ♢ | ♢ | |||||||||||
|
| ♢ | ♢ | ♢ | ♢ | ♢ | |||||||||
|
| ♢ | ♢ | ♢ | ♢ | ||||||||||
|
| ♢ | ♢ | ||||||||||||
|
| ♢ | ♢ | ♢ | |||||||||||
|
| ♢ | ♢ | ♢ | |||||||||||
|
| ♢ | ♢ | ♢ | |||||||||||
|
| ♢ | ♢ | ♢ | |||||||||||
|
| ♢ | ♢ | ♢ | ♢ | ||||||||||
|
| ♢ | ♢ | ♢ |
| Species / Diagnostic characters | leaf shape | leaf margin | lamina | perichaetial leaves | vaginula | capsule position | exostome teeth | endostome segments | preperistome | spore size (µm) | calyptra | asexual repr. |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
| lanceolate | revolute | unistratose | not differentiated | occasionally hairy | long exserted | 8 pairs | absent | present | 14–18 | hairy | not observed |
|
| ovate-lanceolate | revolute | unistratose | not differentiated | naked | emergent | 8 pairs | 8 (or 16) | absent | 15–20 | naked or sparsely hairy | gemmae (uncommon) |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| ovate-lanceolate | recurved | unistratose | not differentiated | naked | shortly emergent | 16 teeth | mostly absent | present | 15.5–20 | hairy | not observed |
|
| ovate-lanceolate | revolute | unistratose | not differentiated | naked | immersed to emergent | 8 pairs (splitting) | 16 segments | absent | 18–22 | hairy | gemmae |
|
| ovate-lanceolate | revolute | unistratose | not differentiated | naked | emergent to shortly exserted | 8 pairs | 8 segments | absent | 12–15 | hairy | gemmae |
|
| ovate-lanceolate | plane or slightly recurved | unistratose | differentiated | hairly | long exserted | 16 teeth | 16 segments | absent | 31–34 (–40) | naked | not observed |
|
| linear-lanceolate | plane or revolute | unistratose | not differentiated | naked | long exserted | 8 pairs | 8 segments | absent | 13–19 | with few scattered hairs | gemmae (occasionally) |
|
| narrow lanceolate | plane | bistratose | differentiated | naked | emergent to just exserted | 8 pairs | 8 segments | absent | 14–16 | naked | not observed |
|
| ovate-lanceolate | broadly revolute | unistratose | differentiated | naked | emergent to shortly exserted | 8 pairs | 8 segments | absent | 14–18 | with few scattered hairs | not observed |
|
| ovate-lanceolate | reflexed | unistratose | not differentiated | hairly | shortly exserted | 8 pairs | 8 segments | mostly absent | 20–23 | with few scattered hairs | not observed |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBryophyte Studies and Records · European Cultural and National Identity · Botany and Plant Ecology Studies
Introduction
Orthotrichum Hedw. is a cosmopolitan moss genus, mainly distributed in temperate regions of both northern and southern hemispheres. Similarly, as is the case with most of such moss genera, Orthotrichum has been recognized as a heterogeneous taxon (Sawicki et al. 2017). Extensive taxonomic and molecular investigations have confirmed its polyphyly, resulting in the separation of three distinct segregates from the genus: Nyholmiella Holmen & E. Warncke; Pulvigera Plášek, Sawicki & Ochyra and Lewinskya F. Lara, Garilleti & Goffinet (Sawicki et al. 2009, 2010, 2017; Plášek et al. 2015; Lara et al. 2016). In South America, the genus Orthotrichum s. str. displays a widespread presence, primarily inhabiting the forests or bushes of temperate regions. These mosses predominantly thrive as epiphytes on trees and shrubs, occasionally as epilithic species on boulders and rocks (Lewinsky 1984).
Orthotrichum in Chile was comprehensively treated by Lewinsky (1984). Subsequent taxonomic findings and new distribution data were further documented by Lewinsky and Deguchi (1989), Müller (2002, 2009), Buck (2005), Ireland et al. (2006, 2017), Goffinet et al. (2007), Medina et al. (2013), Larraín (2016), Larraín and Bahamonde (2017), Drapela and Larraín (2020), Lara et al. (2021), and Ellis et al. (2023). According to these studies, Orthotrichum s. str. is there represented by eight species and two varieties (see also Table 1): Orthotrichumanomalum Hedw., O.assimile Müll. Hal., O.cupulatumvar.austroamericanum Lewinsky, O.diaphanumvar.podocarpi (Müll. Hal.) Lewinsky, O.freyanum Goffinet, W.R. Buck & M.A. Wall, O.gigantosporum Lewinsky, O.inclinatum Müll. Hal., O.perexiguum Dusén ex Lewinsky, O.tristriatum Lewinsky, and O.truncatum Lewinsky & Deguchi. This paper describes a new species from Chile, bringing the total number of Orthotrichum taxa in the country to eleven.
Previously, Orthotrichumaequatoreum was also reported from the territory of Chile (Ireland et al. 2006). However, a subsequent review of the material revealed that this was a misidentification (Ireland et al. 2017).
Material and methods
Plant material
During a bryofloristic survey in the Andes Mountains in 2021, a remarkable epiphytic moss from the genus Orthotrichum was collected. Specimens were carefully gathered, air-dried, and sent to the University of Ostrava for identification and inclusion in the herbarium collections (herbarium OSTR). The material proved to be the first record of this taxon for Chile and, upon closer examination, also to represent a species new to science. Plants were subsequently documented using an Olympus SZ61 trinocular microscope for macrophotographs and Olympus BX53 and IPOS-810 microscopes for microphotographs. Detailed SEM photo-documentation of peristome structures and spores was carried out using a Jeol SEM microscope. All photographs were captured from the holotype (OSTR #8123).
Molecular analyses
Total genomic DNA from a single individual was extracted using the Qiagen Mini Spin Plant Kit (Qiagen, Germany). Details concerning DNA quantification and nanopore sequencing are identical to those in the previous studies (Plášek et al. 2023; Sawicki et al. 2023), but due to lower than required for native DNA sequencing amount of extracted DNA, the low input protocol for library preparation was used. The PCR amplification of total DNA was performed using EXP-PCA001 (Oxford Nanoporetech, UK, hereafter ONT) expansion module followed by SQK-LSK114 kit (ONT) protocol. The beads-based post PCR cleaning stage was replaced by column based method using Clean-Up kit (A&A Biotechnology, Poland) and remaining steps and reagents were as recommended in Lingation sequencing V14 - low input by PCR protocol (ONT). Prepared library was sequenced using FLO-MIN114 (ONT) flow cell and sequenced using Minion Mk1C device. Raw reads were basecalled using Dorado 0.5.1 (ONT) using SUP model with enabled duplex read calling. For downstream analyses, reads containing duplex flags were extracted using Samtools software (Danecek et al. 2021). Application duplex high quality reads (Q>30) allow assembling error-free plastomes using exclusively nanopore sequencing technology (Sawicki et al. 2024).
Obtained raw reads were trimmed using porechop 0.2.4 and assembled using Flye 2.91 assembler (Kolmogorov et al. 2019), which produced complete, circularized plastome contigs. The complete chloroplast genome was annotated using previously published Orthotrichum Hedw. sequences (Mizia et al. 2019; Frangedakis et al. 2021; Plášek et al. 2023) as references in Geneious Prime 2023.2.1 software (Biomatters, Auckland, New Zealand) and deposited in GenBank under PP274123 accession number. The newly sequenced genome was aligned with previously analyzed dataset of Orthotrichaceae plastomes (Plášek et al. 2023) using MAFFT 7.52 (Katoh and Standley 2013). The second copy of IR was removed from subsequent analyses and ambiguously aligned regions were trimmed by Gblocks 0.91 (Talavera and Castresana 2007). The plastome map was drawn using Chloe web server (https://chloe.plastid.org/).
Chloroplast sequences of 22 specimens of Orthotrichaceae, including seven from Orthotrichum were used for phylogenetic analysis. Phylogenetic analysis was carried out using the Bayesian inference (BI) according to model and parameters used in previous study (Plášek et al. 2023). The discrete molecular diagnostic characters (MDCs) for each species of Orthotrichum, were calculated according to the Jörger and Schrödl (2013) approach using FASTACHAR 0.2.4 (Merckelbach and Borges 2020).
Results
Taxonomic treatment
Orthotrichum
camanchacanum
Taxon classificationPlantaeOrthotrichalesOrthotrichaceae
Plášek, Sawicki & Osorio sp. nov.
2E6E3165-D7E8-5322-8988-A04F04ED3CEC
Diagnosis.
Plantae olivacea, obscure viridis, usque ad 1-cm altae. Folia erecta, lineari-lanceolata, carinata, apicibus longis acuminatis. Capsulae emergentes, cylindricae. Stomata cryptopora. Vaginula dense pilosa cum capilli longi. Peristomium duplex, exostoma 8 paribus dentium siccitate erectum, endostoma 16 segmentis. Calyptra dense pilosa. Sporae 19–24 µm, leniter papillosae.
Type.
Chile, Región de Coquimbo (Region IV), Provincia del Elqui, Comuna de Coquimbo, 2 km southeast of Totoralillo town, GPS: 30°04'26"S, 71°21'13"W (-30.073972, -71.353583), on hills profoundly influenced by the humidity generated by camanchaca, vegetation formed mainly by shrubs (Adesmiaargyrophylla Phil. and Echinopsischiloensissubsp.chiloensis (Colla) H.Friedrich & G.D.Rowley), moss was found epiphytically on bark of shrubs, leg. F. Osorio 4378, 10 Aug 2021, holotype (OSTR #8123); isotype (VALD s.n.).
Description.
Plants in dense tufts to 1 cm tall, olive green above, dark green to brown below (Fig. 2). Stem moderately branched, branches up to 5 mm long. Rhizoids well developed, mainly at base of stems. Stem leaves erect to slightly recurved when dry, spreading to slightly recurved when moist; in upper third linear-lanceolate, long acuminate, 3.0–4.1 × 0.3–0.5 mm, carinate; costa ending just below apex or more often excurrent in hyaline long arista. Leaves in the middle and lower part of the stem wider and shorter, ovate-lanceolate, acuminate, 3.0–3.6 × 0.4–0.7 mm, carinate; costa ending just below apex or rarely excurrent in short aristae. Lamina of stem leaves unistratose, margins entire, recurved from the base to two-thirds of the leaf. Hyaline aristae rectangular, formed by (1–)2–3 gradually elongating cells, from 50–65 to 90–115 × 8–10 µm. Upper laminal cells isodiametric to short elongate, (9–)10–15 × 8–11 µm, fairly thick-walled, with one low papillae on both side; basal laminal cells elongate rectangular to rhomboidal, thick-walled, (18–)20–45 × 10–12 µm, smooth. Alar cells slightly differentiated. Sexual condition goniautoicous. Perichaetial leaves differentiated, ovate-lanceolate, acuminate, significantly shorter than stem leaves, only 1.7–2.2 × 0.5–0.8 mm; upper cells forming conspicuous hyaline (sometimes denticulate) apex (Fig. 3). Seta 1.1–1.5 mm long, ochrea up to 1/5 of the seta, vaginula densely hairy with 0.8–1.9 mm long, single-rowed or sometimes double-rowed, smooth hairs, which usually reach the base of the urn (Fig. 4). Capsule emergent; cylindric to oblong-ovoid, about 1.8–2.2 mm long, yellowish brown, slightly constricted below the mouth when dry. Exothecial cells differentiated mainly in the upper half of capsule, urn strongly furrowed when dry. Stomata cryptopore, scattered in the lower part of the urn, more than half covered by subsidiary cells. Peristome double (Figs 4, 5), preperistome absent. Exostome of 8 pairs of teeth, yellow to light brown, erect-spreading when dry. The outer peristome layer (OPL) ornamentation formed by dense papillae below and a mixture of papillae and distinct striae above. The primary peristome layer (PPL) finely and evenly ornamented by vermiculous lines, slightly also with low papillae. Endostome segments 16, almost as tall as exostome, reflexed when dry; 8 main segments double-rowed and 8 intermediate thinner, single-rowed, somewhat shorter or completely broken in matured capsules. The inner peristome layer (IPL) smooth or ornamented indistinctly by vermiculous lines in the lower part. Calyptra conic-oblong, more or less plicate, yellowish with longitudinal brown stripes, apex red-brown, densely hairy with long, yellowish, smooth hairs. Lid conic, apiculate. Spores light brown, 19–24 µm, densely papillose. Asexual reproduction not observed.
Distribution and ecology.
Moss Orthotrichumcamanchacanum was discovered growing epiphytically on the bark of shrubs within vegetation predominantly composed of shrubs such as Adesmiaargyrophylla and Echinopsischiloensissubsp.chiloensis. This epiphytic growth pattern suggests a specific ecological niche for this moss within the ecosystem. See map (Fig. 1) for a visual representation of the study area. The territory under investigation lies within the Coquimbo region, characterized by numerous transverse valleys. Notably, the Elqui valley, where Orthotrichumcamanchacanum was observed, is situated within this region. The environmental conditions in the study area are influenced by the Andes mountain range, contributing to a steppe-like climate. This climate is typified by sparse vegetation, consisting primarily of shrubs and scrubby vegetation. Precipitation, the heaviest of which occurs during the winter months, further shapes the ecological dynamics of the region.
The map illustrates the geographical location where Orthotrichumcamanchacanum was discovered.
Macro photographs of Orthotrichumcamanchacanum. View on fertile plants: A plants with capsules at various stages of development: young immature capsule (left), capsule closed by lid (middle), and mature capsule with open peristome (right) B dried furrowed capsules emergent from long leaves. Scale bars: 1 mm (A, B). Photographs were taken from the holotype (OSTR #8123).
Micro photographs of Orthotrichumcamanchacanum leaves A–G leaves from upper part of a stem (linear-lanceolate with remarkable narrow hyaline apex) H–N leaves from middle and lower part of a stem (many of them are damaged and without apical part) O–S detail views of laminar cells, abaxial views (O apical part P upper part Q middle part R lower part and S base of leaf) T–V perichaetial leaves W leaf sections (from apical to basal part). Scale bars: 1 mm (A–N); 100 µm (O–S); 0.5 mm (T–V); 50 µm (W). Photographs were taken from the holotype (OSTR #8123).
Macro and micro photographs of Orthotrichumcamanchacanum sporophyte characters A, B view on mature capsules C, D mature capsule with peristome E detail of peristome F stoma (immersed) on capsule urn G lid H, I spores J, K calyptra covered by long hairs L, M hairy vaginula. Scale bars: 1 mm (A, B); 0.5 mm (C, D); 100 µm (E); 50 µm (F); 0.1 mm (G); 20 µm (H, I); 0.5 mm (J–M). Photographs were taken from the holotype (OSTR #8123).
SEM photographs of OrthotrichumcamanchacanumA capsule B double peristome C–E detailed view on exostome teeth and endostome segments F spores. Scale bars: 500 µm (A); 100 µm (B); 50 µm (C–E); 20 µm (F). Photographs were taken from the holotype (OSTR #8123).
Etymology.
The authors are delighted to name the species (camanchacanum) after “camanchaca”, a term derived from Aymara native language, signifying marine cloud (fog). Forming along the Chilean coast as a cloud, the camanchaca transforms into a dense fog as it moves inland towards the mountains. This fog provides the humidity essential for plant survival.
Molecular survey
Chloroplast genome of Orthotrichumcamanchacanum sp. nov. was 123,409 bp long and had a typical quadripartite structure with one small single-copy (SSC), one large single-copy (LSC), and two inverted repeats (IR). The use of third-generation sequencing did not identify any structural heteroplasmy associated with inversions in the SSC region. The newly sequenced plastome comprises 82 gene encoding proteins (including hypothetical chloroplast reading frames like ycf1, 2, 3, 4, 12, and 66), 32 transfer RNA (tRNA), and four ribosomal RNA (rRNA) genes. Notably, the rps12 gene is split into two separate transcription units, namely 5’-rps12 and 3’-rps12, and their transcripts undergo trans-splicing (Fig. 6).
Chloroplast genome of newly described Orthotrichumcamanchacanum. Gray inner circle indicates GC content. Red bars indicate normalized score form reference genome (Orthotrichumrogeri) while blue bars incidence percentage of reference genome features.
Analysis of molecular diagnostics characters (MDCs) revealed 641 SNPs characteristic for Orthotrichumcamanchacanum followed by 646 for O.rogeri and 890 in the case of O.cupulatum. Higher numbers of MDCs were detected for O.crenulatum and O.stellatum, 932 and 1304 respectively.
The phylogenetic relationships based on complete plastomes sequences results in a tree (Fig. 7) with all nodes maximally supported by Bayesian posterior probabilities (1.0).The Orthotrichaceae species were found to split into two separate groups, or clades. The first clade includes genera from the Lewinskyinae subgroup, namely Lewinskya, Pulvigera, and Ulota. The second clade consists of genera from the Orthotrichinae subgroup, which are Nyholmiella, Stoneobryum D.H. Norris & H. Rob., and Orthotrichum s. str. Upon analysis, each of these genera was confirmed to be monophyletic. Within these groupings, Ulota was identified as the closest relative to the combined Pulvigera/Lewinskya clade. Similarly, Nyholmiella was found to be most closely related to Orthotrichum. The newly described Orthotrichumcamanchacanum resolved as sister to O.crenulatum/O.rogeri clade.
The Bayesian inference tree based on complete chloroplast genomes. All clades have maximum PP values (1.0).
Discussion
The combination of characters of the newly described species, Orthotrichumcamanchacanum, is distinctive, making it easily recognizable. Upon initial observation, the most notable feature of the gametophyte is the shape of the stem leaves, particularly those in the upper third of the stem. They exhibit a linear-lanceolate form with a narrow apex that terminates in a long, hyaline arista (Fig. 3A–G, O). When comparing the shape of the stem leaves, the majority of the Chilean species within the Orthotrichum genus generally have ovate-lanceolate leaves (Table 2) with shorter apices. Only O.inclinatum (in Lewinsky 1984 as O.compactum Dusén) shares a similarly linear-lanceolate leaf shape, but it differs from O.camanchacanum in other gametophyte characters, such as the absence of a long aristate hyaline apex in the stem leaves, a naked vaginula, and the absence of differentiated perichaetial leaves. Adding sporophyte characters, then, O.inclinatum has long exserted capsule, naked vaginula and different combination of peristome (8+8).
Although Orthotrichumperexiguum also exhibits a narrow and lanceolate leaf shape (Lewinsky 1984), this species is notably small, reaching only up to 0.5 cm. Moreover, it can be distinguished by a bistratose laminal in its marginal parts, a naked vaginula, and a different peristome combination (8+8).
The perichaetial leaves produced by Orthotrichumcamanchacanum are distinctive and markedly different when compared with all South American species of this genus. Notably, they possess a unique apex formed by a group of conspicuous hyaline cells (Fig. 3T–V). In contrast, the perichaetial leaves of other Chilean Orthotrichum species that produce them (such as O.gigantosporum, O.perexiguum, and O.tristriatum), are similar in shape to the stem leaves, differing mostly in their smaller size (Lewinsky 1984).
The hairiness of the vaginula is considered a distinctive taxonomic character for identifying Orthotrichum species (Lewinsky 1993). However, in some European species, this character has not always proven suitable due to its considerable variability (Plášek and Sawicki 2010). Among the Chilean species, three exhibit a distinctly hairy vaginula (O.camanchacanum, O.gigantosporum, and O.truncatum), while in one other species (O.anomalum), its hairiness was only occasionally noted (Lewinsky 1984; Lewinsky and Deguchi 1989). In O.camanchacanum, the hairs in vaginula are dense and long, often reaching the base of the urn or clearly visible among upper stem leaves.
When comparing the sporophyte characters of the Chilean species within the genus Orthotrichum, most of them, including the newly described species, have emergent or shortly exserted capsules. However, three species (O.anomalum, O.gigantosporum, and O.inclinatum) produce significantly exserted capsules on a long seta (Lewinsky 1984; Lewinsky and Deguchi 1989). In Orthotrichum species, the peristome is of the arthrodontous type and primally diplolepidous. Typically, it consists of eight pairs of exostome teeth, occasionally splitting into 16, and eight or sixteen endostome segments. The combination of the number of teeth and segments, along with their surface ornamentation, is a crucial taxonomic feature for both the genus and the entire family Orthotrichaceae (Lewinsky 1993). In the case of O.camanchacanum, its peristome is formed by eight pairs of exostome teeth with the outer peristome layer densely papillose below and a mixture of papillae and striae in the upper part. A similar OPL surface is observed in O.alpestre Bruch & Schimp., a European species not found in South America (cf. Lara et al. 2009; Plášek and Ochyra 2020). Regarding the endostome of the newly described species, it consists of 16 thin segments, which are delicate and often partially broken. However, in well-developed capsules, all the segments are significantly long, almost reaching the length of the exostome teeth. Only two other Chilean species, O.assimile and O.tristriatum, have similarly long and thin endostome segments, but both of them generally have a total of eight segments.
It is widely acknowledged that species within the genus Orthotrichum have immersed stomata, distinguishing them from representatives of the related genus Lewinskya, whose urns possess superficial stomata (Lewinsky 1993). Despite this, some species across both genera exhibit macroscopic similarities. Paradoxically, in the case of Orthotrichumcamanchacanum, certain South American Lewinskya species share a closer resemblance than Orthotrichum species, especially those with distinctly long and narrow leaves culminating in a markedly narrow apex, e.g. L.elongata (Taylor) F. Lara, Garilleti & Goffinet, L.johnstonii (E.B. Bartram) F. Lara, Garilleti & Goffinet or L.mandonii (Schimp. ex Hampe) F. Lara, Garilleti & Goffinet (cf. Lewinsky 1984). However, aside from the already mentioned distinct positions of the stomata, these Lewinskya species also differ in a clear combination of peristome parts. This marked difference in characters prevents any confusion between the newly described species and representatives of Lewinskya species.
The territory where the new species was collected possesses distinctive geographical and climatic conditions. The entire Coquimbo region features several transverse valleys that cut across perpendicular to the main Andes mountain chain, carving through the country horizontally (Alaniz and Carvajal 2019). Among these valleys is situated the Elqui interion region, where Orthotrichumcamanchacanum was collected. The Andes contribute the Región de Coquimbo, providing a steppe-like climate characterized by bushy, scrubby vegetation and heaviest precipitation in the winter (Moreira-Muñoz 2011). On the coast, especially on the coastal mountain range influenced by the moderating effect of the ocean, an abundance of clouds and coastal fog, known as camanchaca, fosters the growth of Chile’s northernmost forests (Moreira-Muñoz 2011). Inland, the climate becomes warm and typically dry (Alaniz and Carvajal 2019). This varied landscape relief combined with a unique climate may have significantly contributed to the speciation of this new species. The moss O.camanchacanum exhibits morphological characters that distinguish it from related species. The fact that it has not been found despite intensive research carried out in Chile since 2011 by the first author may be attributed to its geographical isolation and consequently a very limited area of occurrence. It seems to be endemic to this region of Chile. This situation is not unique; similar examples exist, such as the moss Lewinskyajohnstonii, which also possesses unique morphological features and is limited to the National Park Fray Jorge and its close surroundings (Lewinsky 1984). This area similarly exhibits a specific and characteristic climate.
Supplementary Material
XML Treatment for Orthotrichum camanchacanum
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Alaniz AJ Carvajal MA (2019) Chile: Environmental History, Perspectives and Challenges. NOVA Science Publishers, 1–306.
- 2Buck WR (2005) Exploración briológica en Cabo de Hornos. 3a parte.Briolatina 53: 1–5.
- 3Buck WR Goffinet B (2010) Preliminary key to the mosses of Isia Navarino, Chile (Prov. Antartica Chilena). Nova Hedwigia.Beiheft 138: 215–229.
- 4Danecek P Bonfield JK Liddle J Marshall J Ohan V Pollard MO Whitwham A Keane T Mc Carthy SA Davies RM Li H (2021) Twelve years of SA Mtools and BC Ftools. Giga Science 10(2): giab 008. 10.1093/gigascience/giab 008PMC 793181933590861 · doi ↗ · pubmed ↗
- 5Drapela P Larraín J (2020) The bryophytes of Madre de Dios Archipelago, Magallanes Region, Chile.Phytotaxa 428(1): 7–29. 10.11646/phytotaxa.428.1.2 · doi ↗
- 6Ellis LT Aceñolaza PG Alvarez D Bednarek-Ochyra H Brzęczek D Cabezudo B Cedrés-Perdomo RD Cubas B Sde Faria Lopes S Eckstein J Eiroa D Escolà-Lamora N Fedosov VE Guerra J Jiménez-Alfaro B JukonienėI Kiebacher T Koponen T Strgulc Krajšek S Kutnar L Lavrinenko KV Losada-Lima A Mamontov Yu S Muñoz J Nejfeld P Norhazrina NO’Leary SV Papp BPérez-Haase A Plášek V Porley RD SabovljevićM Schnyder N Shkurko AV Silva JBŠirka P Stebel A Suárez GM Syazwana N Uotila P Virchenko VM (2023) New national and regional bryophyte records, 74.Journal of Bryology 45(3): 249–262. 10.1080/037366 · doi ↗
- 7Frangedakis E Guzman-Chavez F Rebmann M Markel K Yu Y Perraki A Tse SW Liu Y Rever J Sauret-Gueto S Goffinet B Schneider H Haseloff J (2021) Construction of DNA tools for hyperexpression in Marchantia chloroplasts.ACS Synthetic Biology 10(7): 1651–1666. 10.1021/acssynbio.0c 0063734097383 PMC 8296666 · doi ↗ · pubmed ↗
- 8Goffinet BA Buck WR Wall MA (2007) Orthotrichumfreyanum (Orthotrichaceae), a new epiphytic moss from Chile.Nova Hedwigia 131: 1–11.