Acid-treated Staphylococcus aureus induces acute silkworm hemolymph melanization
Yasuhiko Matsumoto, Eri Sato, Takashi Sugita, Jian Xu, Jian Xu, Jian Xu, Jian Xu

TL;DR
This study shows that acid-treated Staphylococcus aureus activates the innate immune response in silkworms, as indicated by increased hemolymph melanization.
Contribution
The novel finding is that acid treatment of S. aureus enhances its ability to induce innate immunity in silkworms, possibly through bacterial proteins.
Findings
Acidification of Staphylococcus aureus increases silkworm hemolymph melanization.
Protease treatment inhibits the melanization activity of acid-treated S. aureus.
C. acnes culture supernatant lowers pH and enhances S. aureus-induced immune activation.
Abstract
The skin microbiome maintains healthy human skin, and disruption of the microbiome balance leads to inflammatory skin diseases such as folliculitis and atopic dermatitis. Staphylococcus aureus and Cutibacterium acnes are pathogenic bacteria that simultaneously inhabit the skin and cause inflammatory diseases of the skin through the activation of innate immune responses. Silkworms are useful invertebrate animal models for evaluating innate immune responses. In silkworms, phenoloxidase generates melanin as an indicator of innate immune activation upon the recognition of bacterial or fungal components. We hypothesized that S. aureus and C. acnes interact to increase the innate immunity-activating properties of S. aureus. In the present study, we showed that acidification is involved in the activation of silkworm hemolymph melanization by S. aureus. Autoclaved-killed S. aureus (S. aureus…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
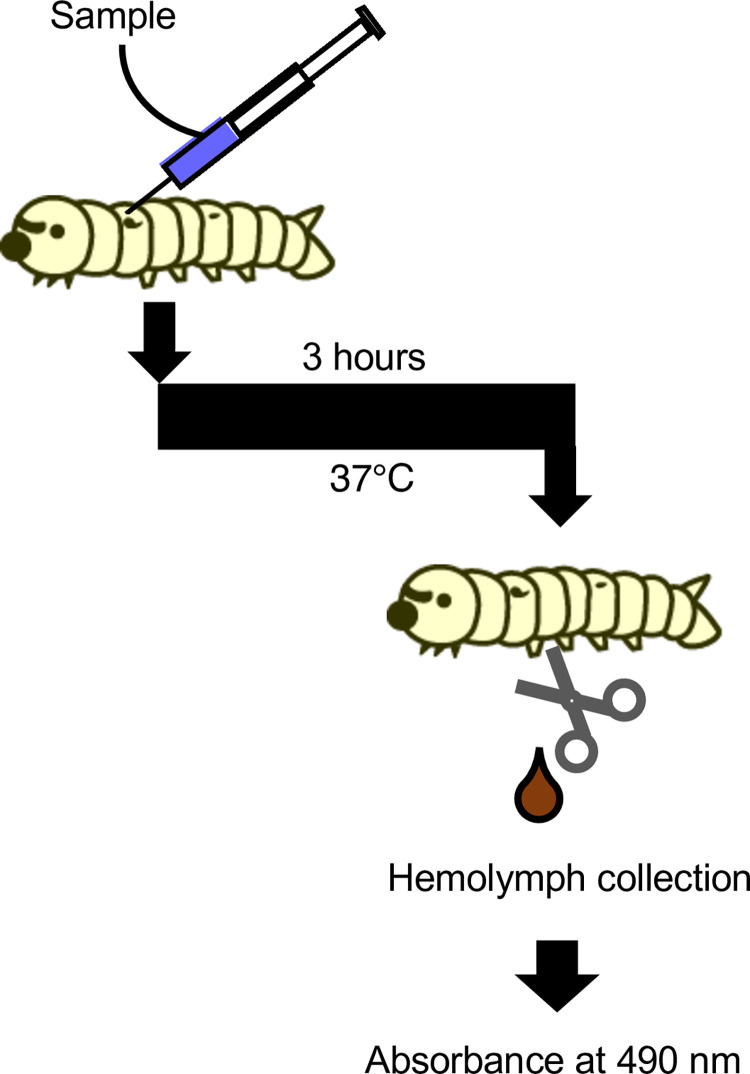 Fig 1
Fig 1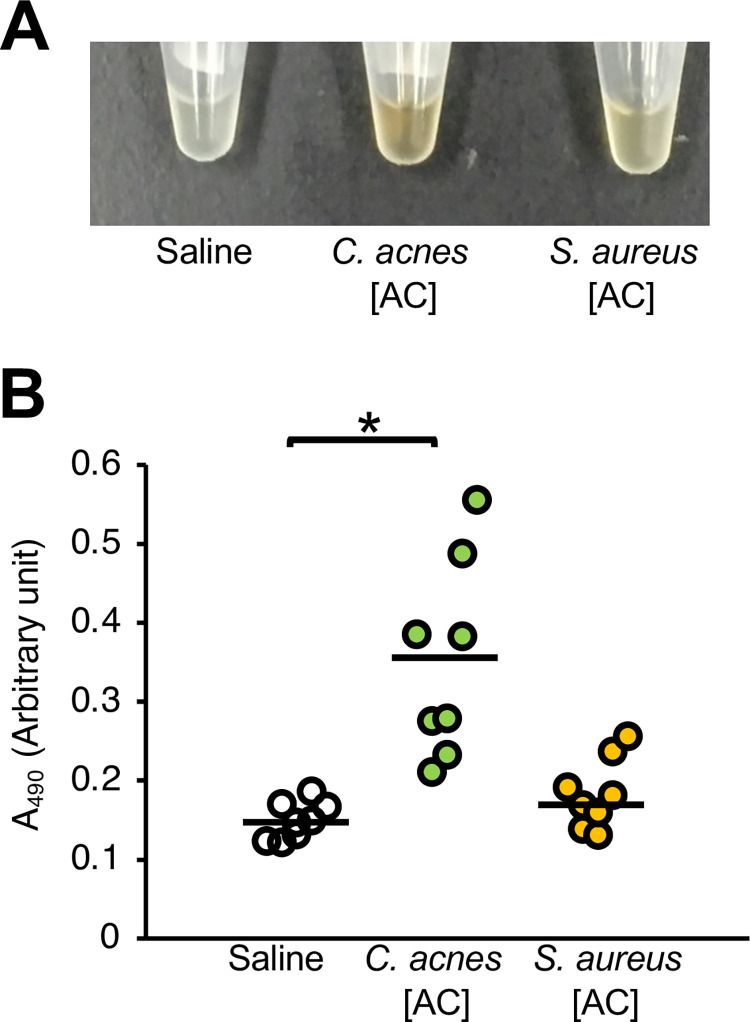 Fig 2
Fig 2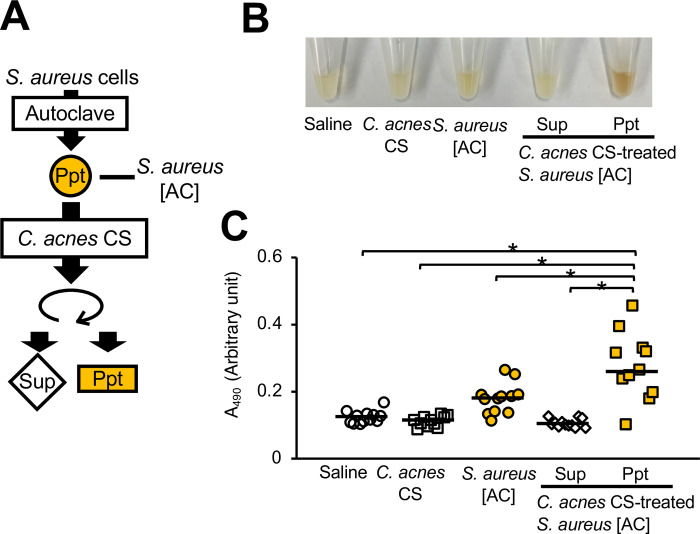 Fig 3
Fig 3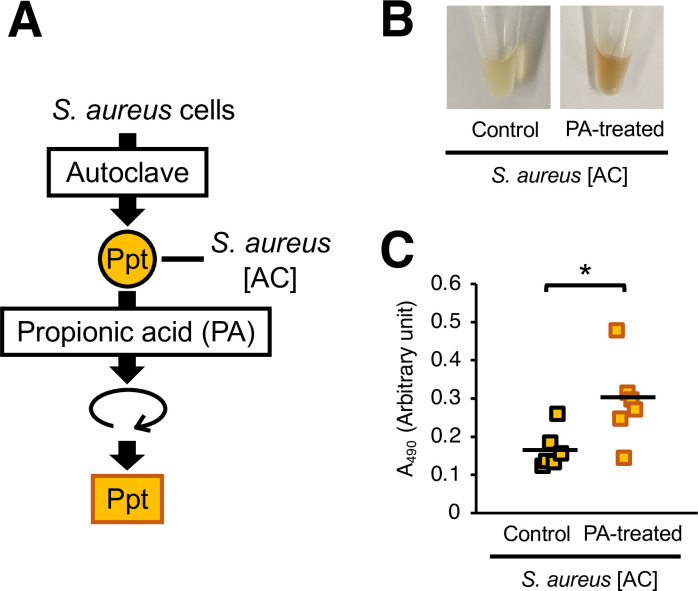 Fig 4
Fig 4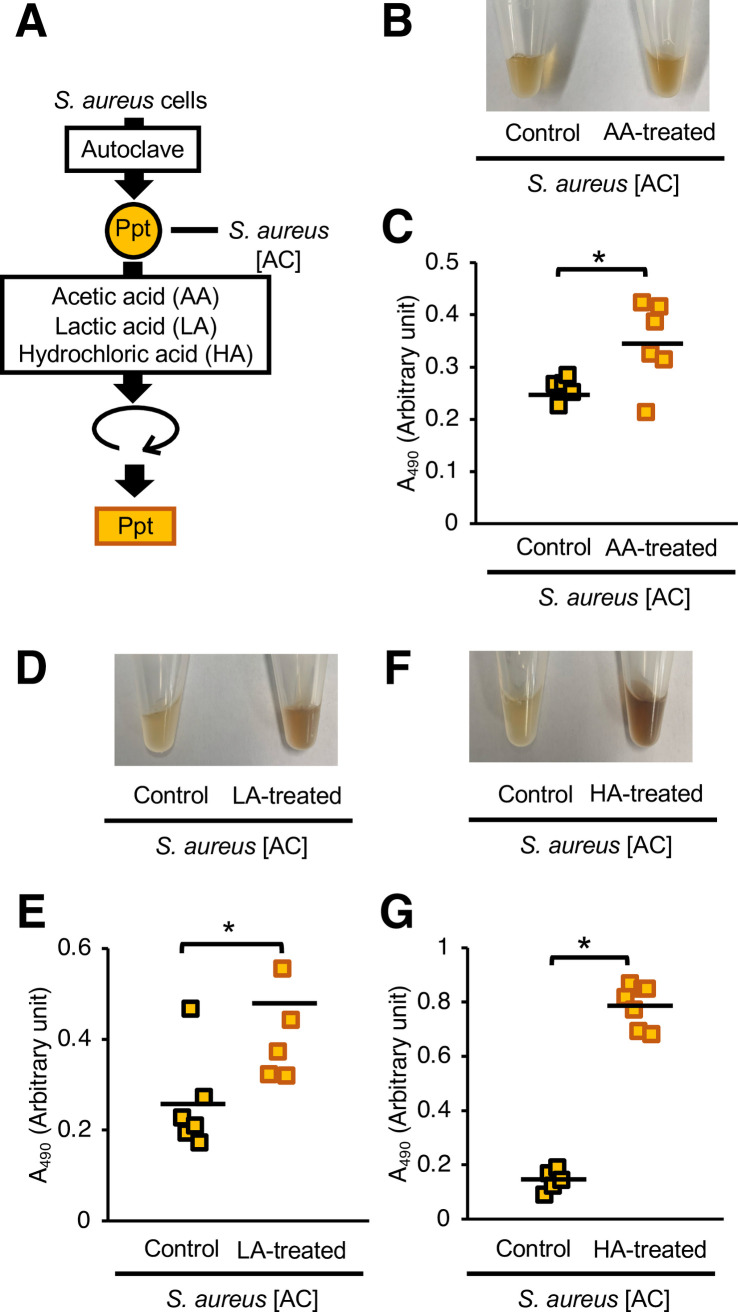 Fig 5
Fig 5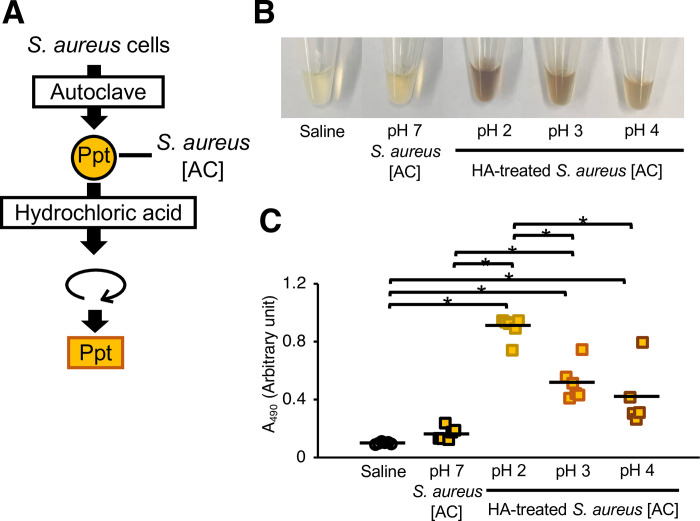 Fig 6
Fig 6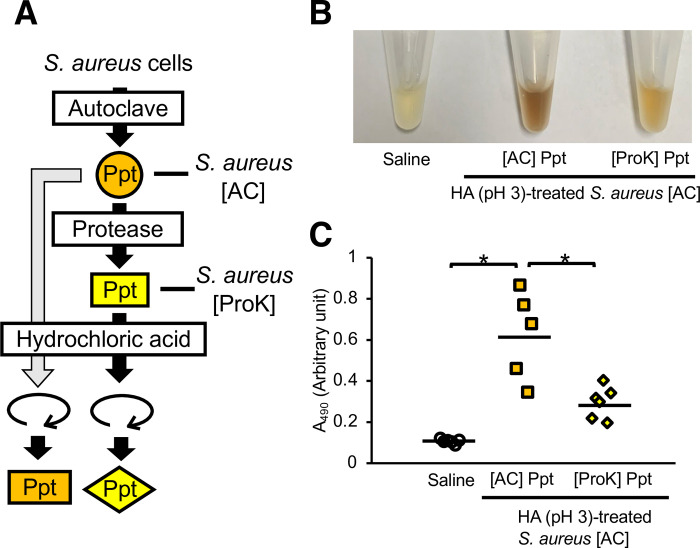 Fig 7
Fig 7- —http://dx.doi.org/10.13039/501100001691Japan Society for the Promotion of Science
- —http://dx.doi.org/10.13039/100009619Japan Agency for Medical Research and Development
- —Kose Cosmetology Research Foundation
- —http://dx.doi.org/10.13039/501100007173Bio-oriented Technology Research Advancement Institution
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsInvertebrate Immune Response Mechanisms · Insect Utilization and Effects · Insect Pest Control Strategies
Introduction
The human skin microbiome maintains the skin environment to prevent the onset of inflammatory skin diseases such as folliculitis and atopic dermatitis [1–3]. Staphylococcus aureus, a gram-positive bacterium on human skin and in the nasal cavity, causes severe systemic infections such as sepsis and inflammatory skin infections such as folliculitis and atopic dermatitis [4–6]. S. aureus enters the hair follicles and proliferates, which induces immune responses and inflammation resulting in folliculitis [2, 5]. S. aureus peptidoglycan, a cell wall component, and proteins such as lipoproteins induce innate immune activation and inflammation [6, 7]. Understanding the mechanism underlying the induction of host immunity by S. aureus may contribute to the prevention and treatment of inflammatory skin diseases.
Cutibacterium acnes, a gram-positive bacterium on the human skin, is a causative agent of inflammatory skin diseases, such as acne vulgaris [1, 8, 9]. Acne vulgaris is an inflammatory skin disease that affects hair follicles and sebaceous glands [9]. C. acnes interacts with S. aureus in hair follicles [5]. Various factors such as lipase and propionic acid, which slightly acidify the skin surface, are produced by C. acnes, [10, 11]. Lipase released by C. acnes produces free fatty acids from sebum [12]. C. acnes also secretes short-chain fatty acids such as propionic acid to lower the pH of the skin environment [13]. The effects of these acidic substances associated with C. acnes on the induction of innate immunity by S. aureus, however, remain unknown.
Innate immune activation occurs in multiple immune cells through several pattern-recognition proteins [14, 15]. Therefore, the use of individual animals is desirable for evaluating the activation of immunity. Experiments using a large number of mammals such as mice trigger ethical issues from the perspective of animal welfare [16, 17]. The silkworm, an invertebrate, has benefits as a model animal to overcome these ethical issues [16, 18, 19]. Moreover, experimental systems using silkworms have been established to evaluate the innate immune activation by pathogenic microorganisms [20–22]. The melanization response is an innate immune mechanism of insects, including silkworms [23–25]. When pathogens enter the silkworm body, the silkworms produce melanin in the hemolymph to coagulate the pathogens and repair the wound [20, 25, 26]. Melanization and immune responses via the Toll pathway are mediated by the same signaling cascades [23–25, 27]. Therefore, silkworm hemolymph melanization is a useful indicator for evaluating the induction of innate immunity by pathogens such as Cutibacterium acnes, Porphyromonas gingivalis, and Candida albicans [20–22].
In this study, using silkworm hemolymph melanization as an indicator of innate immune activation, we found that treatment with C. acnes culture supernatant enhanced the immune-inducing activity of S. aureus. Moreover, treatment with acidic substances such as propionic acid, acetic acid, lactic acid, and hydrochloric acid enhanced the immune-inducing activity of S. aureus. These findings suggest that acidification is involved in immune induction by S. aureus.
Materials & methods
Reagents
Gifu anaerobic medium agar was purchased from Nissui Pharmaceutical Co., Ltd. (Tokyo, Japan). Tryptic soy broth was purchased from Becton Dickinson (Franklin Lakes, NJ, USA). Protease K was purchased from QIAGEN (Hilden, Germany). Propionic acid, acetic acid, lactic acid, and hydrochloric acid were purchased from FUJIFILM Wako Pure Chemical Corporation (Osaka, Japan).
Culture of bacteria
C. acnes ATCC6919 and S. aureus Newman strains were used in this study. The C. acnes ATCC6919 strain was spread on Gifu anaerobic medium agar and incubated under anaerobic conditions at 37°C for 3 days [22]. The S. aureus Newman strain was spread on tryptic soy broth agar and incubated under aerobic conditions at 37°C for 1 day [22]. To prepare the C. acnes culture supernatant (CS), C. acnes ATCC6919 strain (8 x 10^9^ cells) was added to 10 mL of tryptic soy broth + 2% glucose medium and incubated at 37°C for 6 days under anaerobic conditions.
Silkworm rearing
The silkworm rearing procedures were previously described [28]. Silkworm eggs (Hu Yo × Tukuba Ne) were purchased from Ehime-Sanshu Co. Ltd. (Ehime, Japan), disinfected, and hatched at 25°C –27°C. Silkworms were fed an artificial diet, Silkmate 2S, containing antibiotics purchased from Ehime-Sanshu Co., Ltd. Fifth-instar larvae were used for the infection experiments.
In vivo melanization assay
An in vivo melanization assay was performed as previously described [22], with slight modifications. The silkworm injection experiments were performed as previously described [29]. Fifth instar silkworm larvae were fed an artificial diet (1.5 g Silkmate 2S; Ehime-Sanshu Co., Ltd) overnight. A 50-μL suspension of bacterial samples was injected into the silkworm hemolymph with a 1-ml tuberculin syringe (Terumo Medical Corporation, Tokyo, Japan). The silkworms were maintained at 37°C for 3 h. Hemolymph was collected from the larvae through a cut on the first proleg as described previously [30]. The silkworm hemolymph (50 μL) was mixed with 50 μL of physiologic saline solution (0.9% NaCl: PSS). Absorbance at 490 nm was measured using a microplate reader (iMark™ microplate reader; Bio-Rad Laboratories Inc., Hercules, CA, USA). Each experiment was performed at least twice to check reproducibility.
Protease treatment
Autoclaved S. aureus cells [AC] were diluted with phosphate-buffered saline (PBS) to an absorbance at 600 nm (A_600_) = 3 in 1 mL, and 50 μL protease K (0.75 AU/ml) was added. After incubation for 1 h at 50°C, the samples were centrifuged at 15,000 rpm for 10 min at room temperature. The precipitate was suspended in PSS (1 mL), and the remaining enzymes were inactivated by incubation at 80°C for 30 min. The samples were centrifuged at 15,000 rpm for 10 min at room temperature and the precipitate was diluted with PSS to A_600_ = 1 to obtain the precipitate sample.
Statistical analysis
Statistical differences between groups were analyzed using the Student’s t-test, the Tukey’s test or the Tukey-Kramer test. The Student’s t-test was used to assess whether the two groups were statistically significantly different. The Tukey’s test and the Tukey-Kramer test were used to assess whether the multiple groups were statistically significantly different. Each experiment was performed at least twice. A P value of less than 0.05 was considered statistically significant.
Results
Comparison of melanization-inducing activity between heat-killed C. acnes and S. aureus
The injection of heat-killed C. acnes cells obtained by autoclaving causes silkworm hemolymph melanization [22]. We examined whether heat-killed C. acnes and S. aureus cells differed in their ability to induce silkworm hemolymph melanization. The experimental scheme for silkworm hemolymph melanization is shown in Fig 1. Melanization was induced by injection of autoclave-treated C. acnes, but not by that of autoclave-treated S. aureus (Fig 2A and 2B). The result suggests that the silkworm hemolymph melanization activity induced by heat-killed S. aureus is lower than that by heat-killed C. acnes.
Evaluation system for inducing silkworm hemolymph melanization using bacterial samples.Illustration of an experimental method to determine silkworm hemolymph melanization. Sample solution was injected into the silkworm hemolymph. The silkworms were maintained at 37°C for 3 h. The hemolymph was collected from the larvae through a cut on the first proleg. Absorbance was measured at 490 nm.
*Comparison of silkworm hemolymph melanization induced by heat-killed C. acnes and S. aureus.(A, B) Sample solution was injected to silkworms and hemolymph was collected at 3 hours after injection. Samples were saline (Saline), heat-killed C. acnes cell suspension (C. acnes; 1 x 108 cells/larva), or heat-killed S. aureus cell suspension (S. aureus; 1 x 108 cells/larva). (A) Photograph. (B) Absorbance at 490 nm (A490). n = 8/group. Statistically significant differences between groups were evaluated using the Tukey’s test. P < 0.05.
Increased melanization-inducing activity of S. aureus after treatment with C. acnes culture supernatant
Silkworm hemolymph melanization was increased by injecting silkworms with S. aureus [AC] treated with C. acnes CS (Fig 3). We confirmed that C. acnes CS alone did not induce silkworm hemolymph melanization (Fig 3). These results suggest that the substances produced by C. acnes induced the silkworm hemolymph melanization activity of S. aureus.
*Silkworm hemolymph melanization was induced by injecting heat-killed S. aureus treated with C. acnes culture supernatant.(A) Preparation of C. acnes culture supernatant (CS)-treated S. aureus [AC]. The S. aureus [AC] fraction was treated with C. acnes CS at 37°C for 24 h. (B, C) Sample solution was injected to silkworms and hemolymph was collected at 3 hours after injection. Samples were saline (Saline), C. acnes CS, S. aureus [AC], C. acnes CS-treated S. aureus [AC] supernatant (Sup), or precipitate (Ppt). (B) Photograph. (C) Absorbance at 490 nm (A490). n = 11-12/group. Statistically significant differences between groups were evaluated using the Tukey-Kramer test. P < 0.05.
Silkworm hemolymph melanization by acid-treated S. aureus cells
C. acnes produces short-chain fatty acids such as propionic acid [13], which affect the pH in the environment [3, 31]. The pH of the C. acnes CS used in this study was lower than that of the preculture medium (Table 1). Therefore, we tested whether silkworm hemolymph melanization by S. aureus [AC] was induced by pretreatment with propionic acid. Propionic acid-treated S. aureus [AC] induced silkworm hemolymph melanization (Fig 4). Next, we examined whether acids other than propionic acid increased the silkworm hemolymph melanization activity of S. aureus [AC]. Compared with untreated S. aureus [AC], S. aureus [AC] treated with acetic acid, lactic acid, or hydrochloric acid also exhibited higher silkworm hemolymph melanization activity (Fig 5). On the other hand, administration of these short-chain fatty acid solutions did not affect melanization of the silkworm hemolymph (S1 Fig in S1 File). Furthermore, injections of S. aureus [AC] treated with low-pH solutions adjusted by hydrochloric acid at 37°C for 24 h also enhanced silkworm hemolymph melanization in a pH-dependent manner, with more melanization occurring at a lower pH (Fig 6). These results suggest that acid treatment induces silkworm hemolymph melanization activity by S. aureus.
*Silkworm hemolymph melanization induced by the injection of heat-killed S. aureus treated with propionic acid.(A) Preparation of propionic acid (PA)-treated S. aureus [AC]. The S. aureus [AC] fraction was treated with propionic acid (1%: 130 mM) at 37°C for 24 h. (B, C) Sample solution was injected to silkworms and hemolymph was collected at 3 hours after injection. Samples were S. aureus [AC] or propionic acid (130 mM)-treated S. aureus [AC] precipitates (Ppt). (B) Photograph. (C) Absorbance at 490 nm (A490). n = 6/group. Statistically significant differences between groups were evaluated using the Tukey-Kramer test. P < 0.05.
*Effects of acid treatment of heat-killed S. aureus on silkworm hemolymph melanization.(A) Preparation of the acid-treated S. aureus [AC]. The S. aureus [AC] fraction was treated with acetic acid (AA) (130 mM), lactic acid (LA) (130 mM), or hydrochloric acid (HA) (130 mM) at 37°C for 24 h. (B, C) Sample solution was injected to silkworms and hemolymph was collected at 3 hours after injection. Samples were S. aureus [AC], AA, LA, or HA-treated S. aureus [AC] precipitates (Ppt). (B) Photograph. (C) Absorbance at 490 nm (A490). n = 5-7/group. Statistically significant differences between groups were evaluated using the Student’s t-test. P < 0.05.
*Effects of low-pH treatments of heat-killed S. aureus on silkworm hemolymph melanization.(A) Preparation of the acid-treated S. aureus [AC]. The S. aureus [AC] fraction was treated with saline or hydrochloric acid solution adjusted to pH 2–7 at 37°C for 24 h. (B, C) Sample solution was injected to silkworms and hemolymph was collected at 3 hours after injection. Samples were S. aureus [AC] or low pH-treated S. aureus [AC] precipitate (Ppt). (B) Photograph. (C) Absorbance at 490 nm (A490). n = 5-6/group. Statistically significant differences between groups were evaluated using the Tukey-Kramer test. P < 0.05.
Table 1: Decrease in the pH of medium by the addition of C. acnes culture.
Effect of protease treatment on the induction of silkworm hemolymph melanization by S. aureus
S. aureus lipoproteins bind to mammalian TLR2 and activate innate immunity [6, 7]. Therefore, we hypothesized that a protein in S. aureus [AC] was responsible for this activity. Melanization of the silkworm hemolymph by S. aureus [AC] treated with low pH was decreased by pretreating the S. aureus [AC] with a protease (Fig 7). On the other hand, administration of protease solution did not affect melanization of the silkworm hemolymph (S1 Fig in S1 File). These results suggest that S. aureus proteins are involved in the induction of silkworm hemolymph melanization by acid-treated S. aureus.
*Effect of protease treatment of heat-killed S. aureus on silkworm hemolymph melanization.(A) Preparation of the acid-treated S. aureus [AC] with protease treatment. S. aureus [AC] was treated with protease K. Protease K-treated S. aureus [AC] was further treated with hydrochloric acid solution adjusted to pH 3 at 37°C for 7 days. (B, C) Sample solution was injected to silkworms and hemolymph was collected at 3 hours after injection. Samples were saline (Saline), acid-treated S. aureus [AC] precipitate ([AC] Ppt), or acid-treated protease K-treated S. aureus [AC] precipitate ([ProK] Ppt). (B) Photograph. (C) Absorbance at 490 nm (A490). n = 5-6/group. Statistically significant differences between groups were evaluated using the Tukey-Kramer test. P < 0.05.
Discussion
Acid treatment of S. aureus stimulates innate immune activity in silkworms, and the innate immune activating substances of S. aureus may be proteins. Our findings suggest that acid-treated S. aureus induces excess host immunity, which may cause immune-active diseases, such as folliculitis.
S. aureus and C. acnes are gram-positive bacteria present in the human skin that cause inflammatory diseases such as folliculitis [2]. We hypothesized that an interaction between S. aureus and C. acnes within the hair follicle may influence the induction of innate immune responses by S. aureus. In the present study, we found that acidic substances released by C. acnes affected the immune-inducing activity of S. aureus. Further studies should be conducted in mammalian models to confirm and develop these findings.
On the human skin, various bacteria, including C. acnes and S. aureus, produce short-chain fatty acids such as lactic acid [32]. Moreover, S. aureus and C. acnes produce lipases that produce free fatty acids from sebum [13], which can decrease the pH of the skin. On the other hand, bacteria such as Delftia acidovorans, a gram-negative bacterium on human skin, produce ammonia that increases the pH [33]. Therefore, the balance of the skin microbiome may affect changes in skin pH. Weak acidity caused by alterations in the skin microbiome may induce an innate immune response by S. aureus. Proteins of S. aureus may be involved in the immune activity induced by acid-treated S. aureus in the silkworm evaluation system. The conformational changes of proteins responsible for inducing innate immunity by acid treatment, however, remain unknown. We assumed that S. aureus lipoproteins treated with acids lead to conformational changes, and that acid-treated lipoproteins are easily recognized by Toll receptors, which are involved in innate immunity in silkworms. Preventing the acid-enhanced induction of innate immune activation by S. aureus may contribute to inhibiting the onset of inflammatory skin diseases such as folliculitis.
As a limitation of this study, the effects of acid-treated S. aureus on inflammation in individual mammals were not evaluated. It is also unclear whether the effects of an acidic pH on the skin microbiome are related to immune induction. Furthermore, the responsible proteins of S. aureus have not yet been identified. The effects of acid treatment and the identification of the responsible proteins are important issues for future research.
Conclusion
Acid-treated S. aureus induces innate immunity in silkworms, and S. aureus proteins may be the responsible factors.
Supporting information
S1 File(DOCX)
S1 DatasetDatasets included in this study.(XLSX)
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Byrd AL, Belkaid Y, Segre JA. The human skin microbiome. Nat Rev Microbiol. 2018;16: 143–155. doi: 10.1038/nrmicro.2017.157 29332945 · doi ↗ · pubmed ↗
- 2Lousada MB, Lachnit T, Edelkamp J, RouilléT, Ajdic D, Uchida Y, et al. Exploring the human hair follicle microbiome. Br J Dermatol. 2021;184: 802–815. doi: 10.1111/bjd.19461 32762039 · doi ↗ · pubmed ↗
- 3Smythe P, Wilkinson HN. The Skin Microbiome: Current Landscape and Future Opportunities. Int J Mol Sci. 2023;24: 3950. doi: 10.3390/ijms 24043950 36835363 PMC 9963692 · doi ↗ · pubmed ↗
- 4Leung DYM. Atopic dermatitis: New insights and opportunities for therapeutic intervention. J Allergy Clin Immunol. 2000;105: 860–876. doi: 10.1067/mai.2000.106484 10808164 · doi ↗ · pubmed ↗
- 5Nowicka D, Grywalska E. Staphylococcus aureus and Host Immunity in Recurrent Furunculosis. Dermatology. 2019;235: 295–305. doi: 10.1159/000499184 30995649 · doi ↗ · pubmed ↗
- 6Hulme J. Staphylococcus Infection: Relapsing Atopic Dermatitis and Microbial Restoration. Antibiotics. 2023;12: 222. doi: 10.3390/antibiotics 12020222 36830133 PMC 9952585 · doi ↗ · pubmed ↗
- 7Iwamoto K, Moriwaki M, Miyake R, Hide M. Staphylococcus aureus in atopic dermatitis: Strain-specific cell wall proteins and skin immunity. Allergol Int. 2019;68: 309–315. doi: 10.1016/j.alit.2019.02.006 30878567 · doi ↗ · pubmed ↗
- 8Stirling A, Worthington T, Rafiq M, Lambert PA, Elliott TS. Association between sciatica and Propionibacterium acnes. Lancet. 2001;357: 2024–2025. doi: 10.1016/s 0140-6736(00)05109-6 11438138 · doi ↗ · pubmed ↗