The Segregation of p.Arg68Ter-CLDN14 Mutation in a Syrian Deaf Family, Phenotypic Variations, and Comparative Analysis with the GJB2 Gene
Abdelaziz Tlili, Abdullah Al Mutery, Jihen Chouchen

TL;DR
This study identifies a genetic mutation in a Syrian family with hearing loss and compares it to another gene involved in deafness.
Contribution
The study identifies a novel nonsense mutation in the CLDN14 gene and compares it with GJB2 to explain phenotypic differences.
Findings
A nonsense mutation in the CLDN14 gene was found to segregate with hearing loss in a Syrian family.
The CLDN14 mutation was not present in the control population, supporting its pathogenicity.
A comparative analysis between CLDN14 and GJB2 was conducted to explain differences in mutation frequency.
Abstract
Hearing impairment, a rare inherited condition, is notably prevalent in populations with high rates of consanguinity. The most common form observed globally is autosomal recessive non-syndromic hearing loss. Despite its prevalence, this genetic disorder is characterized by a substantial genetic diversity, making diagnosis and screening challenging. The emergence of advanced next-generation sequencing (NGS) technologies has significantly advanced the discovery of genes and variants linked to various conditions, such as hearing loss. In this study, our objective was to identify the specific variant causing hearing loss in a family from Syria using clinical exome sequencing. The proband in the family exhibited profound deafness as shown by pure-tone audiometry results. The analysis of the different variants obtained by NGS revealed the presence of a nonsense mutation within the CLDN14…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
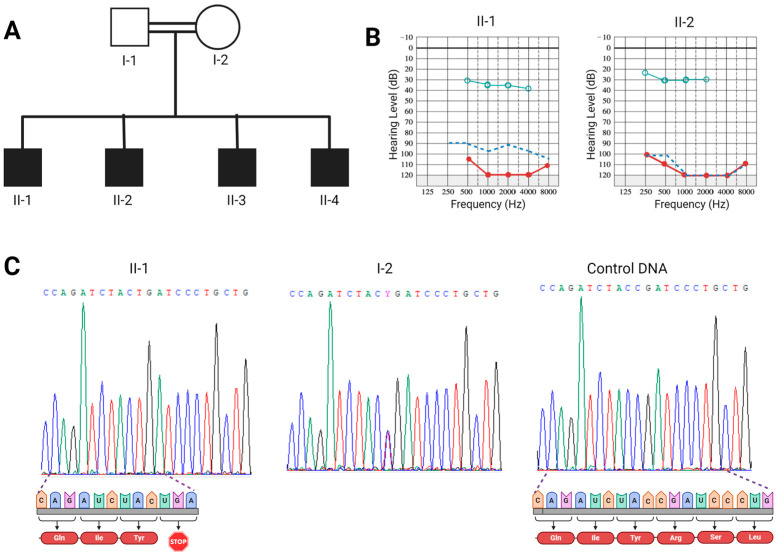 Figure 1
Figure 1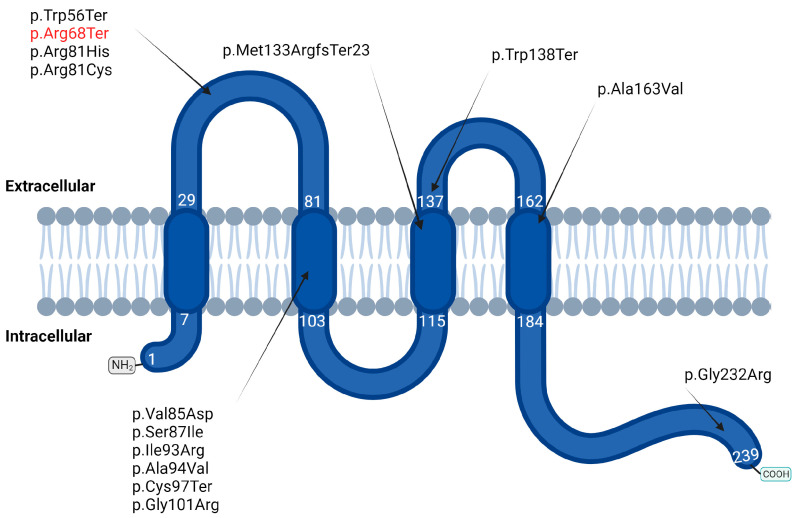 Figure 2
Figure 2- —University of Sharjah
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsHearing, Cochlea, Tinnitus, Genetics · Ear Surgery and Otitis Media · Vestibular and auditory disorders
1. Introduction
Hearing loss (HL) is the most commonly occurring neurosensorial defect, affecting around 1/1000 children [1]. Genetic factors are responsible for a minimum of 60% of nonsyndromic sensorineural hearing loss cases [2]. Different inheritance patterns associated with this type of hearing loss were reported. Autosomal recessive nonsyndromic hereditary hearing loss (ARNSHL) is the most predominant form representing approximately 70% of all nonsyndromic hereditary hearing loss cases [1]. To date, mutations in over 75 genes have been identified as causes of ARNSHL “https://hereditaryhearingloss.org/ (accessed on 1 February 2024)”. Among these genes is CLDN14, which is associated with a specific form of ARNSHL known as DFNB29 [3].
CLDN14, that encodes for claudin proteins (~26 KDa), is involved in the regulation of epithelial barrier function, particularly at bicellular junctions [4]. This gene is expressed in various locations within the inner ear. The expression profile of CLDN14 in the inner ear suggests its involvement in preserving the integrity of various compartments, most notably, the organ of Corti [5]. Murine studies showed a rapid deterioration of hair cells in the knockout cldn14−/− mouse, probably due to the basolateral parts of these cells being subjected to high levels of K+-rich endolymph, as a consequence of the absence/malfunction of Claudin 14 protein in the ion barrier [6].
So far, fourteen mutations in the CLDN14 gene have been associated with ARSNHL [3,7,8,9,10,11,12,13,14,15,16,17]. The highest contribution and number of mutations have been reported in the Pakistani population, where the prevalence of CLDN14 pathogenic variants was estimated to be 2.25% of deaf individuals [7]. It is worth noting that even though there is phenotypic variability associated with certain mutations, the impact of CLDN14 mutations tends to be more severe at higher frequencies, with moderate to severe phenotypes reported at low frequencies, while severe to profound phenotypes often exhibited at higher frequencies [3,7,8,10,11,12,13,14].
Although the CLDN14 gene is relatively small, it consists of a single coding exon, has a coding region of 720 nucleotides, and is associated with a limited number of mutations linked to hearing loss; another gene, GJB2, with a similar structure (one single coding exon) is also involved in deafness [18]. However, the GJB2 gene exhibits a high number (approximately 135) of pathogenic variants associated with hearing loss [19]. This gene encodes the gap-junction connexin 26 (Cx26) protein, which serves as a subunit of gap junctions, facilitating communication between neighboring cells. Within the inner ear, Cx26 plays a crucial role in the maintenance of potassium homeostasis [20]. Within the spectrum of GJB2 mutations, many variants have been reported as prevalent in specific populations. These include 35delG, 235delC, V37I, W24X, 167delT8, and R143W, which are common in Mediterranean, East Asian, Southeast Asian, Indian subcontinent, Ashkenazi, and Ghanaian populations, respectively [18,21,22,23,24,25].
In our current investigation, we have identified the p.Arg68Ter-CLDN14 mutation responsible for ARNSHL in a consanguineous Syrian family. Additionally, we conducted a comprehensive review of prior CLDN14 mutations and their corresponding phenotypes. Furthermore, we carried out a comparative analysis between the CLDN14 and GJB2 genes, aiming to elucidate potential factors contributing to the higher mutability of GJB2 when compared to CLDN14.
2. Materials and Methods
2.1. Study Family
A Syrian consanguineous family with four affected children was recruited, and genomic DNA was isolated from saliva samples using the Oragene-DNA (OG-500) Kit (DNA Genotek, Stittsville, ON, Canada). To ensure confidentiality, the DNA samples were appropriately labeled with codes. Additionally, 100 individuals with deafness and 80 hearing participants were incorporated in this study. The parents of affected individuals provided written informed consent after audiological and clinical assessments. Moreover, the Ethics Committee at the University of Sharjah approved all the experimental methods used in this study.
2.2. GJB2 Screening and Whole Exome Sequencing
Individuals with the hearing loss underwent screening for GJB2 mutations through Sanger sequencing and were analyzed using whole exome sequencing (WES) as previously described [26]. Finally, the potential functional impact of the identified variants was estimated using a variety of bioinformatics tools including Variant Effect Predictor (VEP), Mutation Taster, VarSome, PROVEAN, PolyPhen-2, SIFT, and Human Splicing Finder.
2.3. CLDN14 Mutational Screening
To confirm the WES results and the cosegregation of the c.202C>T (p.Arg68Ter) nonsense variant with deafness in the affected family, Sanger sequencing of the CLDN14 coding exon was performed as described previously [12]. To summarize, after PCR amplification using CLDN14-F: ACCACCATCCTGCCGCACTG and CLDN14-R: TGTTTGCAGTGGTCGTGGTG primers, the products were purified and underwent cycle sequencing. The resulting sequences were then aligned with the publicly available sequence of the CLDN14 gene (NM_144492).
2.4. In Silico Analysis
In order to ascertain the characteristics of GJB2 and CLDN14 genes, the following databases were used: QGRS Mapper “https://bioinformatics.ramapo.edu/QGRS/index.php” (accessed on 1 February 2024) for determining putative quadruplex forming G-rich sequences (QGRS) in nucleotide sequences, VectorBuilder “https://en.vectorbuilder.com/” (accessed on 1 February 2024) for the purpose of calculating the GC content, and GnomeAD “https://gnomad.broadinstitute.org/” (accessed on 1 February 2024) for determining the reported variants classified as pathogenic and likely pathogenic in both genes.
3. Results
3.1. Molecular Analysis
A consanguineous Syrian family with four affected deaf children was recruited in this study (Figure 1A). Analysis of the family pedigree suggested that the inheritance pattern is likely autosomal recessive; moreover, clinical investigation did not reveal any other abnormality and confirmed the profound hearing loss phenotype in all siblings. All affected children received cochlear implants in their right ears, resulting in an enhancement of sound field threshold levels. These levels shifted from profound hearing loss (100 or more decibels) to a mild hearing impairment (25–40 decibels) (Figure 1B). Sanger sequencing of the GJB2 gene, the most common gene related to deafness, showed an absence of mutations in the affected individuals. Thus, we undertook whole exome sequencing for the affected individual I-1 to determine the causative mutation. Our investigation unveiled a total of 32,808 DNA variations, of which 11,726 were found to be homozygous. By choosing unknown variants or those with a frequency of less than 0.01 in the G1000 and gnomAD databases, a total of 49 DNA variations in 45 genes were obtained. After considering genes responsible for hearing loss, we narrowed down the list of candidate variants to a lone known variant located within the CLDN14 gene. It consists of a transition C to T (c.202C>T) that substitutes the residue Arginine 68 by a stop codon (p.Arg68Ter). To validate this finding and confirm the segregation of this nonsense variant with deafness in the affected family, Sanger sequencing was carried out for the affected children and their parents. Our results revealed that all affected individuals were homozygous for this nonsense variant, whereas their parents were heterozygous (Figure 1C). This observation thereby confirms the co-segregation of c.202C>T with deafness in the investigated family. Additionally, screening for this mutation in 100 unrelated individuals with deafness and 80 hearing participants revealed that it was absent in all cases.
3.2. CLDN14 Mutations and Phenotypic Variations
An overview of reported CLDN14 mutations shows that, so far, 14 mutations are associated with ARNSHL (Table 1). The majority of these mutations occur in the second transmembrane domain, and most of them are associated with a severe to profound phenotype (p.Arg81His, p.Ser87Ile, and p.Gly232Arg), with the notable exception of p.Ala94Val and p.Val85Asp which have been associated with moderate to severe hearing loss (Figure 2). It is worth mentioning that five mutations are truncated with four nonsense variants (p.Trp56Ter, p.Arg68Ter, p.Cys97Ter and p.Trp138Ter) and one frameshift (p.Met133ArgfsTer23). Phenotypic details are available for p.Trp138Ter and p.Met133ArgfsTer23, showing a hearing loss phenotype of moderate to profound severity in both cases (Table 1). Although the impact of CLDN14 mutations tends to increase in severity at higher frequencies, the less severe phenotype was observed at higher frequencies for two mutations that involve the replacement of an Alanine residue by a Valine amino acid (p.Ala94Val and p.Ala163Val).
Finally, our review analysis revealed the existence of four variants reported as founder mutations across various populations. These mutations, p.Met133ArgfsX23 and p.Val85Asp, were both identified as founder variants within the Pakistani population, whereas p.Trp138Ter was identified as a founder in the Yemeni population, and p.Ala163Val was reported in the Newfoundland population.
3.3. Comparative Analysis of CLDN14 and GJB2 Genes
Since CLDN14 and GJB2 genes both possess coding regions of roughly equivalent sizes, albeit with distinct mutational rates, we tried to identify potential parameters that might explain the low frequency and number of mutations in CLDN14 compared to GJB2. In fact, using the gnomAD database, a brief review of pathogenic and/or likely pathogenic variants in these two genes demonstrated the presence of 74 variants in GJB2, while the CLDN14 gene showed only 4 (Table 2). To provide an explanatory insight, we investigated their chromosomal locations, their nucleotide sequences, and the number of putative QGRS. Our analysis showed that the GJB2 gene is very close to the centromeric region with a distance of approximately 1.29 Mb from the centromere, while the CLDN14 gene was located at a distance of 23.46 Mb from the centromeric region. The investigation into the coding regions of these two genes revealed that the GJB2 nucleotide sequence has a significant cytosine content, accounting for 37.5% (A (17.5% 126)|C (37.5% 270)|G (26.81% 193)|T (18.19% 131)). In contrast, the cytosine content of the CLDN14 gene amounts to 25.55% (A (23.2% 158)|C (25.55% 174)|G (26.58% 181)|T (24, 67% 168)), which represents a perceptible difference between the two. Finally, the investigation of both nucleotide sequences unveiled the presence of 17 potential QGRS within the GJB2 gene, while the CLDN14 gene revealed a more limited number, with only 4 identified QGRS.
4. Discussion
In the current study, we investigated a Syrian family with ARNSHL. Affected individuals showed profound hearing loss. The analysis of GJB2, which is the primary gene contributing to ARNSHL [27], revealed the absence of pathogenic variants. To identify the responsible DNA variation, WES, which represents an effective method in identifying HL causative mutations [28], was performed for the affected individuals. This approach unveiled the presence of the nonsense DNA variant c.202C>T (p.Arg68Ter) in the CLDN14 gene, which segregates with HL in the studied family. This mutation was previously identified in a simplex Chinese family, but details regarding its segregation and associated phenotype were not provided [17].
Claudin 14, belonging to the claudin protein family, is an essential membrane protein that plays a key role in regulating calcium levels and maintaining ion balance. Various variants of the CLDN14 gene have been linked to hearing loss and the formation of kidney stones [29]. In the knockout mouse model (Cldn14−/−), a degeneration of outer hair cells starts during the first postnatal week, followed by the deterioration of the inner hair cells in the subsequent week [6]. The deterioration of hair cells likely results from the exposure of the hair cell’s basolateral areas to a potassium-rich endolymph, which is facilitated by a defect in the ion barrier impairment due to the absence of the tight junction protein Claudin 14 [6]. To date, 14 pathogenic mutations have been associated with ARNSHL in humans, of which 9 are missense mutations, 3 are nonsense mutations and 1 is a frameshift deletion (Table 1). The phenotype associated with 9 of these variants has been reported, showing a greater degree of hearing loss severity at higher frequencies compared to lower frequencies, which correlates with the degeneration pattern reported in the knockout mouse where the apical region showed more pronounced hair cell degeneration and a more significant loss of presynaptic ribbons compared to the basal region [30]. In this report, it is noteworthy that all affected siblings display profound hearing loss across all frequencies. This stands in contrast to previous mutations reported and the knockout mouse model, where a clear severity gradient was observed. This variation could potentially be attributed to the complete absence or significantly reduced size of the polypeptide resulting from the c.202C>T (p.Arg68Ter) mutation, which lacks both extracellular loops (ECL1 and ECL2). These domains are very important in the regular function of claudins, as ECL1 is vital for controlling the tightness of paracellular junctions and selective ion permeability, whereas ECL2 can lead to a reduction in the width of the paracellular cleft [31].
Moreover, the Cldn14 knockout mice have shown the loss of ribbon synapses in inner hair cells and regression of auditory nerve fibers [30]. Given the important role of the auditory nerve in cochlear-implant-based auditory rehabilitation [32], Claußen et al. proposed exploring the performance of cochlear implantation in Cldn14−/− mice in the future [30]. In the present study, the four deaf children with the c.202C>T (p.Arg68Ter) nonsense variant do not appear to exhibit defects in their auditory nerves, as their cochlear implants have successfully improved their hearing levels. This situation was also reported in a prior investigation, where a cochlear implant was carried out on a patient with the missense mutation p.Arg81Cys, leading to an increase in threshold levels from 77.5 decibels to 37.5 [16].
Finally, the comparison between CLDN14 and GJB2 genes allowed us to identify some potential factors that can explain the low rate of mutations in the CLDN14 gene compared to GJB2. In fact, it appears that chromosomal location seems to play a role in this difference. Many studies have demonstrated that centromeric regions, particularly the heterochromatin ones, tend to undergo late replication [33]. This pattern of late replication leads to a two-fold increase in transition DNA variations and a six-fold rise in transversion DNA variations, in comparison to loci that replicate early [34].
Additionally, the nucleotide composition can be a contributing factor to the lower mutation rate observed in the CLDN14 gene when compared to GJB2. The CLDN14 gene has a C content of 25.55%, whereas the GJB2 gene has a C content of 37.5%. This elevated C content in the GJB2 gene may explain its increased mutation rate as previous studies have showed that Cytosines exhibit a germline mutation rate roughly tenfold greater than that observed in the other nucleotides [35]. Finally, the analysis of both nucleotide sequences showed 17 potential QGRS within the GJB2 gene versus 4 in the CLDN14 gene. The high number of QGRS in the GJB2 gene can be also a contributor to the high mutational rate observed in this gene compared to the CLDN14 one, as many studies showed an association between QGRS and high mutation rates [36].
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Morton N.E. Genetic epidemiology of hearing impairment Ann. N. Y. Acad. Sci.1991630163110.1111/j.1749-6632.1991.tb 19572.x 1952587 · doi ↗ · pubmed ↗
- 2Smith R.J. Bale J.F.Jr. White K.R. Sensorineural hearing loss in children Lancet 200536587989010.1016/S 0140-6736(05)71047-315752533 · doi ↗ · pubmed ↗
- 3Wilcox E.R. Burton Q.L. Naz S. Riazuddin S. Smith T.N. Ploplis B. Belyantseva I. Ben-Yosef T. Liburd N.A. Morell R.J. Mutations in the gene encoding tight junction claudin-14 cause autosomal recessive deafness DFNB 29Cell 200110416517210.1016/s 0092-8674(01)00200-811163249 · doi ↗ · pubmed ↗
- 4Riazuddin S. Ahmed Z.M. Fanning A.S. Lagziel A. Kitajiri S. Ramzan K. Khan S.N. Chattaraj P. Friedman P.L. Anderson J.M. Tricellulin is a tight-junction protein necessary for hearing Am. J. Hum. Genet.2006791040105110.1086/51002217186462 PMC 1698716 · doi ↗ · pubmed ↗
- 5Kitajiri S. Furuse M. Morita K. Saishin-Kiuchi Y. Kido H. Ito J. Tsukita S. Expression patterns of claudins, tight junction adhesion molecules, in the inner ear Hear Res.2004187253410.1016/s 0378-5955(03)00338-114698084 · doi ↗ · pubmed ↗
- 6Ben-Yosef T. Belyantseva I.A. Saunders T.L. Hughes E.D. Kawamoto K. Van Itallie C.M. Beyer L.A. Halsey K. Gardner D.J. Wilcox E.R. Claudin 14 knockout mice, a model for autosomal recessive deafness DFNB 29, are deaf due to cochlear hair cell degeneration Hum. Mol. Genet.2003122049206110.1093/hmg/ddg 21012913076 · doi ↗ · pubmed ↗
- 7Bashir Z. Latief N. Belyantseva I.A. Iqbal F. Riazuddin S.A. Khan S.N. Friedman T.B. Riazuddin S. Riazuddin S. Phenotypic variability of CLDN 14 mutations causing DFNB 29 hearing loss in the Pakistani population J. Hum. Genet.20135810210810.1038/jhg.2012.14323235333 PMC 3596117 · doi ↗ · pubmed ↗
- 8Charif M. Boulouiz R. Bakhechane A. Benrahma H. Nahili H. Eloualid A. Rouba H. Kandil M. Abidi O. Lenaers G. Genetic and molecular analysis of the CLDN 14 gene in Moroccan family with non-syndromic hearing loss Indian J. Hum. Genet.20131933133610.4103/0971-6866.12082824339547 PMC 3841559 · doi ↗ · pubmed ↗