Functional Analysis of the Major Pilin Proteins of Type IV Pili in Streptococcus sanguinis CGMH010
Yi-Ywan M. Chen, Yuan-Chen Yang, Hui-Ru Shieh, Yu-Juan Lin, Wan-Ju Ke, Cheng-Hsun Chiu

TL;DR
This study explores how different pilin proteins affect the function of Type IV pili in Streptococcus sanguinis, revealing their roles in motility and host cell invasion.
Contribution
The study identifies PilA1 as critical for twitching motility and structured biofilm formation in S. sanguinis.
Findings
PilA1 is essential for the assembly of twitching-active Type IV pili in S. sanguinis CGMH010.
Recombinant strains expressing motility form more structured biofilms under shearing forces.
Tfp retraction, not twitching, is essential for host cell invasion in S. sanguinis.
Abstract
The pil gene cluster for Type IV pilus (Tfp) biosynthesis is commonly present and highly conserved in Streptococcus sanguinis. Nevertheless, Tfp-mediated twitching motility is less common among strains, and the factors determining twitching activity are not fully understood. Here, we analyzed the functions of three major pilin proteins (PilA1, PilA2, and PilA3) in the assembly and activity of Tfp in motile S. sanguinis CGMH010. Using various recombinant pilA deletion strains, we found that Tfp composed of different PilA proteins varied morphologically and functionally. Among the three PilA proteins, PilA1 was most critical in the assembly of twitching-active Tfp, and recombinant strains expressing motility generated more structured biofilms under constant shearing forces compared to the non-motile recombinant strains. Although PilA1 and PilA3 shared 94% identity, PilA3 could not…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
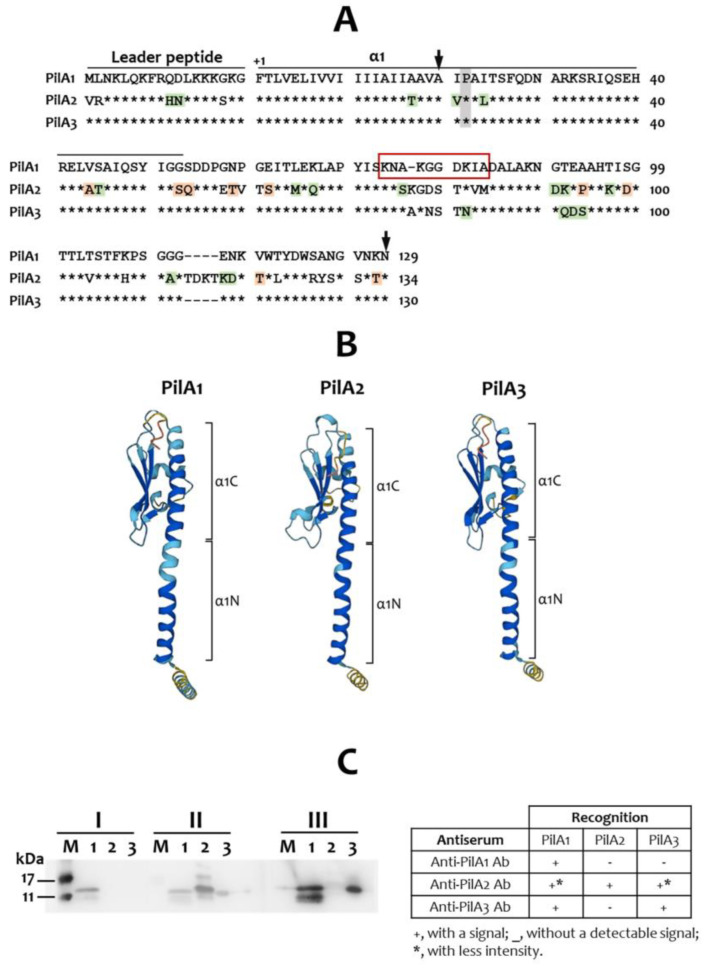 Figure 1
Figure 1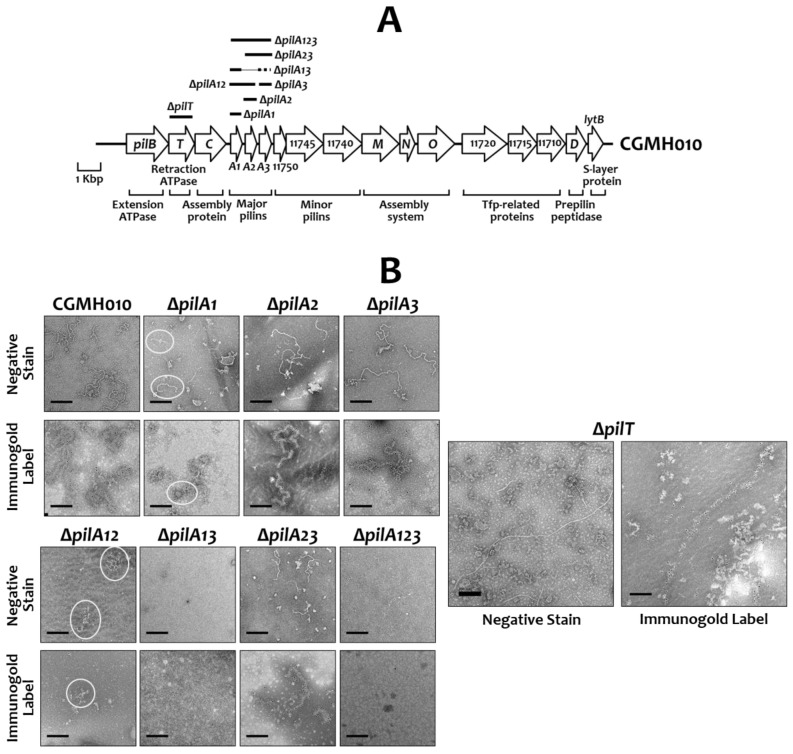 Figure 2
Figure 2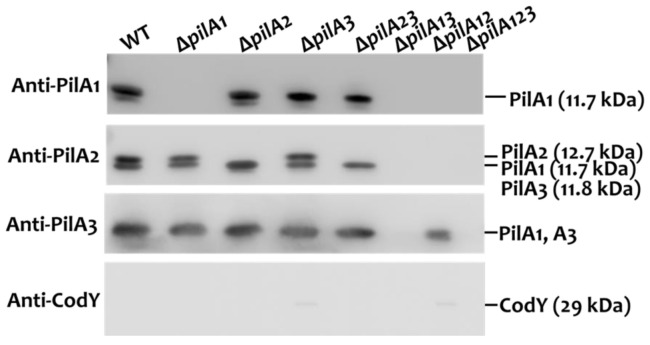 Figure 3
Figure 3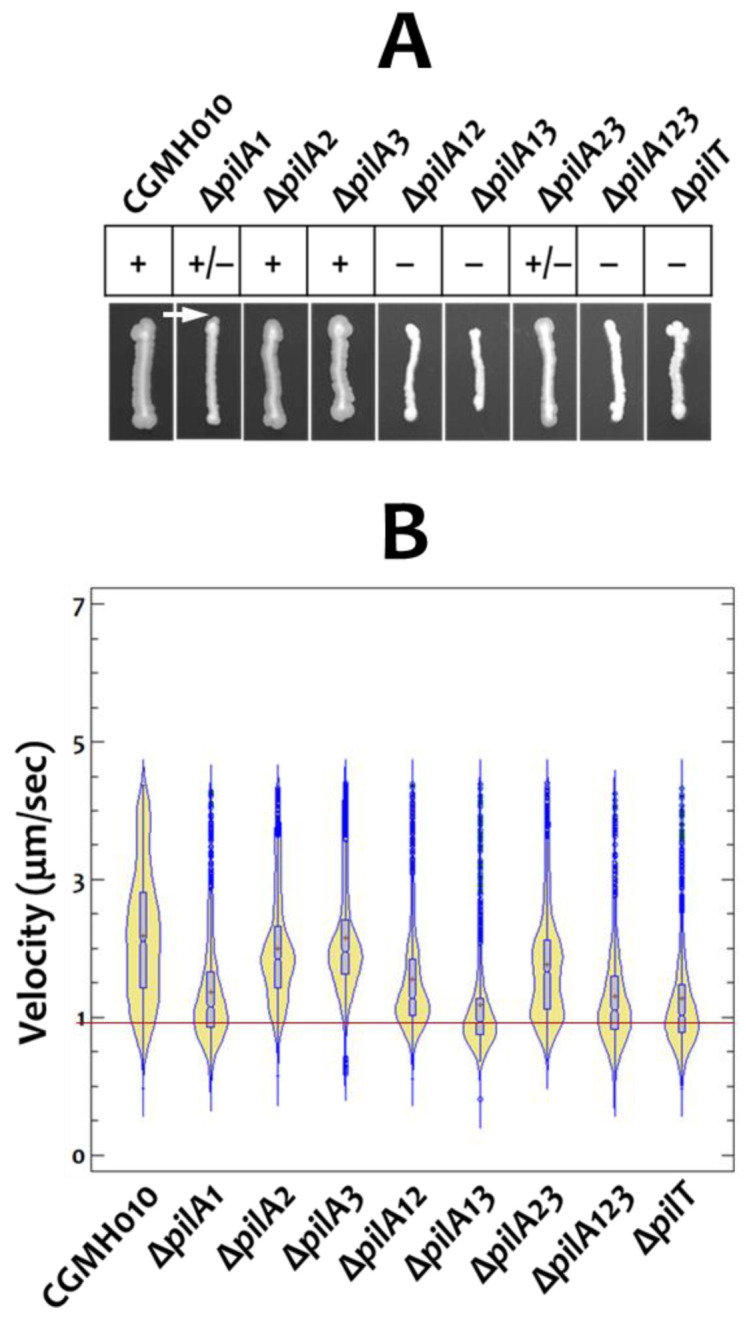 Figure 4
Figure 4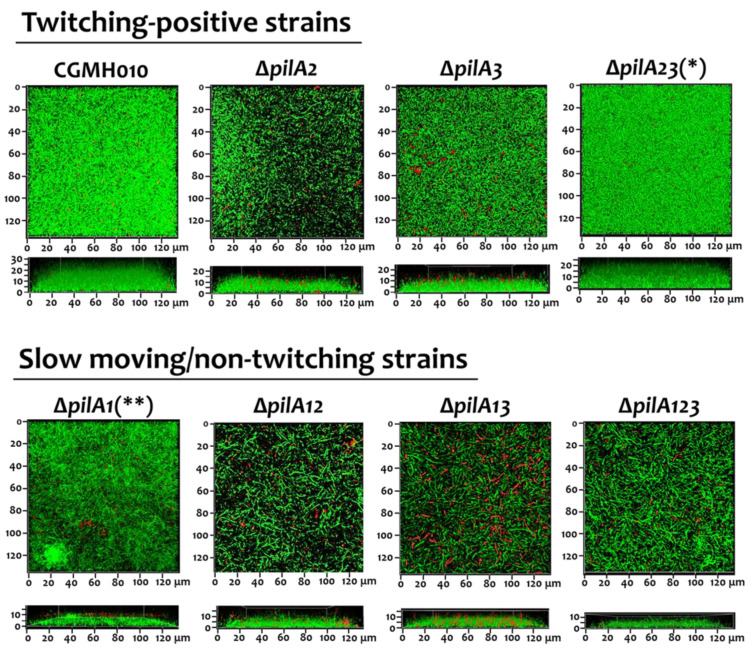 Figure 5
Figure 5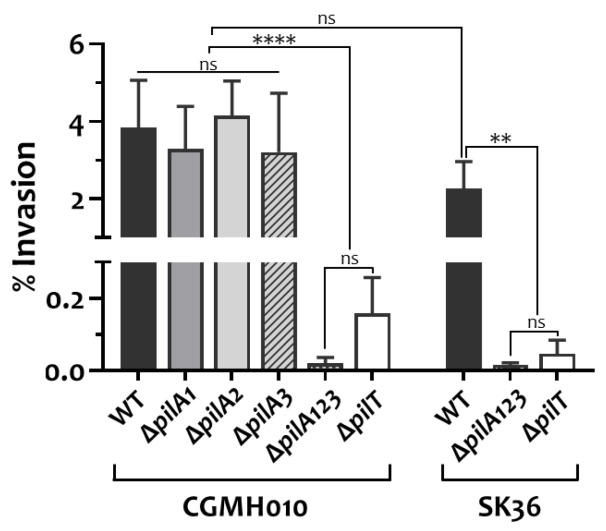 Figure 6
Figure 6- —National Science and Technology Council (NSTC) of Taiwan
- —Chang Gung Memorial Hospital of Taiwan
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsStreptococcal Infections and Treatments · Biochemical and Structural Characterization · Plant Pathogenic Bacteria Studies
1. Introduction
Type IV pili (Tfp), a structural homolog of the type II secretion system [1], are stretchable filaments present on the surface of archaea and bacteria [2]. Tfp are subdivided into Types IVa and IVb based on the leader sequence of the prepilin and the size of the mature major pilin [3]. Both types of Tfp have been extensively studied in Gram-negative bacteria [4]. Type IVa Tfp were recently identified in Gram-positive bacteria, including Clostridium spp. and Streptococcus sanguinis [5], though Type IVb Tfp are found only in Gram-negative bacteria. Genes encoding the proteins required for Type IVa Tfp biosynthesis are typically scattered in a few clusters in Gram-negative bacteria [5] and Clostridium spp. [6,7], while in S. sanguinis, the pil genes are arranged as an operon [8,9].
Tfp are made of major and minor pilins; the major pilins are present in thousands of copies in a pilus, whereas the minor pilins are present in lower abundance [10]. Some minor pilin proteins constitute an initiation complex to prime Tfp biosynthesis, and others act as ligands for interaction with host cells [11,12,13]. Notably, the Gram-negative systems generally feature a single major pilin protein, whereas S. sanguinis commonly harbors two to three homologous major pilin proteins [14]. Additionally, the Tfp system contains two ATPases for extension (PilB) and retraction (PilT/U) of the Tfp, respectively. The continuous action of PilB and PilT/U activates surface-dependent twitching motility [15]. Note that the nomenclature of the pil genes is not standardized; therefore, in this manuscript, we follow the convention used for Pseudomonas aeruginosa Tfp genes.
Tfp are known to mediate surface-dependent twitching motility, genetic transformation, surface sensing, adherence, and biofilm formation [4,16]. The functional significance of Tfp has been extensively studied in Gram-negative pathogens, including Neisseria meningitidis [17] and P. aeruginosa [18,19], while in Gram-positive systems, where Type IVa Tfp have not been widely explored, similar functions have also been described [8,9,14,20,21]. Gram-negative bacteria employ a single system for all observed activities, whereas Gram-positive bacteria utilize two independent Tfp systems for twitching motility and natural competence. Specifically, Streptococcus pneumoniae utilizes Tfp comprising Com proteins for DNA uptake during genetic transformation but not for surface-dependent twitching motility, despite the Tfp being morphologically identical to the motility-driven Tfp [22]. On the other hand, S. sanguinis utilizes a Com system for DNA uptake [23] and the Tfp produced by the pil operon for twitching motility and the adherence of host cells [8,9,14]. Similarly, the Tfp of Clostridium spp. enable twitching motility and biofilm formation [6,7,24,25,26], but are unable to take up extracellular DNA. Finally, the study by Martini et al. demonstrates that S. sanguinis SK36 can invade endothelial cells via the activity of Tfp, highlighting the crucial role of Tfp in pathogenesis [21], as commonly observed in Gram-negative pathogens.
S. sanguinis is a dominant oral isolate and an early colonizer of dental plaque [27]. Advances in genomic studies have confirmed that the pil operon is commonly present in the genome of S. sanguinis strains. Interestingly, S. sanguinis is the only streptococcal species harboring the pil operon, suggesting that Tfp could provide specific advantages for S. sanguinis. Although the arrangement and sequence of the pil genes are conserved, the number of major pilin genes (pilA) varies across strains, with two or three pilA genes being observed. For example, among the most-studied S. sanguinis strains, strains SK36 and CGMH010 contain three major pilin genes, pilA1, pilA2, and pilA3, whereas strain 2908 contains two major pilin genes: pilE1 and pilE2. Most of the pil genes share a sequence identity of approximately 90% or higher at the deduced amino acid level among homologs across strains, though loci encoding the major and minor pilins exhibit less conservation [14]. Previously, we examined the sequences of the pilA genes of 30 clinical isolates and found that although pilA genes are relatively less conserved between strains, significant identities between PilA proteins are present within a strain. Specifically, strains harboring three pilA genes typically display over 90% identity between two of the three pilA paralogs, while the identity with the remaining paralog ranges from 65% to 85%, suggesting that gene duplication occurred during evolution. Furthermore, a study with S. sanguinis 2908 demonstrated that a single major pilin can generate motile Tfp, but Tfp composed of PilE1 exhibit a faster velocity than Tfp made of PilE2 [20]. Whether all PilA proteins in the three-pilA system are required for the biosynthesis of functional Tfp and whether the Tfp-mediated activities are affected by the PilA proteins remain unexplored.
Our recent functional studies on two S. sanguinis strains encoding three pilA paralogs, the motile CGMH010 and non-motile SK36, confirmed that Tfp are essential for optimal adherence to host cells, regardless of motility [14]. Here, we investigated the role of the three highly homologous major pilins of CGMH010 and found that each protein exhibits different effects on the structure and activity of Tfp.
2. Results
2.1. Sequence Analysis of the PilA Proteins and Generation of Anti-PilA Antisera
PilA1 and PilA3 of S. sanguinis CGMH010 share 94% identity at the deduced amino acid level, and PilA2 shares 72% and 70% identity with PilA1 and PilA3, respectively. In agreement with the structural studies of the major pilin proteins [28], the N-terminal regions of PilA1, PilA2, and PilA3 are highly conserved and form an α-helix (α1). Specifically, PilA1 and PilA3 are identical from amino acids 1 to 53, whereas four conserved and two semi-conserved substitutions are present in PilA2. The helix-breaking amino acid, Pro-22, is also present in all three PilA proteins (Figure 1A). The C-terminal region, containing sequences that form the structurally conserved global domain, is less conserved between the PilA proteins. The predicted 3-D structures obtained from the AlphaFold protein structure database also indicate that all PilA proteins exhibit a characteristic “lollipop” architecture [3], with the global domain embedded in the C-terminal half of the α1 (Figure 1B).
To generate antibodies that are specific to each of the PilA proteins, a peptide unique to PilA1 was used to generate a PilA1-specific antiserum (Figure 1A, red box). This antiserum does not cross-react with PilA2 and PilA3. However, the antiserum generated using a histidine-tagged PilA2 protein (His-PilA2) cross-reacts with both PilA1 and PilA3 (with reduced intensity), whereas the antiserum generated by using a histidine-tagged PilA3 protein (His-PilA3) cross-reacts with PilA1 (Figure 1C). Although the anti-PilA2 and anti-PilA3 antisera exhibit cross-reactivity, it is sufficient to detect specific PilA proteins when analyzing the results obtained from all three antisera.
2.2. The Effect of PilA Proteins in the Assembly of Tfp in S. sanguinis CGMH010
To analyze the function of each of the PilA proteins in the structure and the function of Tfp, recombinant S. sanguinis strains with single, double, or triple deletions of the pilA genes were generated in S. sanguinis CGMH010 using non-polar erm or kan, as detailed in the Materials and Methods (Figure 2A). Western blot analysis with anti-PilA antisera demonstrated the presence of non-cognate PilA proteins in total cell lysates of all pilA-deficient derivatives, confirming that mutations were non-polar (Figure S1).
To analyze whether all three PilA proteins are required for the assembly of surface filaments, extracellular Tfp were purified from various strains and examined by transmission electron microscopy (TEM). Filaments were readily detected in the preparations of wild-type CGMH010, strains ΔpilA2, ΔpilA3, ΔpilA23, and ΔpilT (Figure 2B). Filaments were also detected in strain ΔpilA1, but the filaments were more difficult to find, suggesting that the loss of pilA1 affected filament formation. Additionally, the filaments derived from the ΔpilA2 and ΔpilA3 strains were similar to those of wild-type CGMH010, and the filaments purified from the ΔpilA1 and ΔpilA23 strains were shorter than those of the wild-type CGMH010. No filaments were found in ΔpilA13 and ΔpilA123, and only short, aggregated filaments were detected in the preparation from the ΔpilA12 strain. Strain ΔpilT, deficient in the retraction ATPase, generated long filaments. To confirm that the observed filaments were made of PilA proteins, the preparations were also subjected to immuno-gold labeling with anti-PilA2 or anti-PilA3 antiserum (Figure 2B). All observed filaments, regardless of the length, were the products of PilA proteins. Thus, PilA1 and PilA3 alone could generate extracellular filaments, but pili made of PilA3 (strain ΔpilA12) differed from pili derived from PilA1 (strain ΔpilA23), despite the high degree of homology between PilA1 and PilA3.
To further confirm the composition of the extracellular pili in the pilA-deletion mutants, the extracellular pilus preparations were subjected to Western blot analysis with PilA-specific antisera (Figure 3). Anti-CodY antiserum [29] was utilized as a control to ensure that the preparations were free of cytoplasmic proteins. Extracellular pilin proteins were detected in all strains except ΔpilA13 and ΔpilA123, indicating that the deletion of both pilA1 and pilA3 would abolish, or greatly reduce, Tfp biosynthesis.
2.3. The Twitching Activity of Tfp Composed of Different PilA Proteins
Next, we wanted to examine the impact of PilA proteins on Tfp-mediated twitching motility on an agar surface by using the above-described pilA-deletion derivatives of CGMH010. Our results demonstrated that the deletion of pilA1 resulted in the most significant reduction in the twitching zone on TH agar, compared to wild-type CGMH010 (Figure 4A). Among the strains without pilA1 (ΔpilA1, ΔpilA12, ΔpilA13, and ΔpilA123), only the ΔpilA1 strain expressed a marginal twitching zone, whereas no twitching zone was observed in the ΔpilA12 and ΔpilA13 strains. On the other hand, the deletion of pilA2 and pilA3 had little effect on the size of the twitching zone, indicating that PilA2 and PilA3 were dispensable for motile Tfp biosynthesis. The critical role of PilA1 in generating twitching-active pili was further confirmed in strain ΔpilA23, which generated a twitching zone smaller than that of the wild-type strain, but larger than that of strain ΔpilA1. As expected, the pilin-null strain (ΔpilA123) failed to generate a visible twitching zone. Taken together, these results suggest that PilA1 alone can generate a twitching-positive Tfp, whereas PilA2 and PilA3 together cannot fully compensate for the loss of pilA1 in the assembly of twitching-active Tfp.
As the growth rate of all PilA mutant strains was comparable to that of the wild-type CGMH010, presumably, strains that can move at high velocity would generate larger twitching zones compared to strains moving at low velocity after prolonged incubation. To verify this hypothesis, we analyzed the velocity of a single chain of cells by microscopic examination. Using a wet-mount preparation [14], it was found that the bacterial chains of a strain did not move at the same speed (Figure 4B); presumably, chain length would affect the sensitivity of detection. Nevertheless, in agreement with macroscopic examination, the median velocity of the motile strains (ΔpilA2 and ΔpilA3) was more similar to the wild-type CGMH010 than to the non-motile strains (ΔpilA12, ΔpilA13, ΔpilA123, and ΔpilT). Strain ΔpilA23, which generated a reduced twitching zone on the agar surface, exhibited a wide range of velocity, with a higher number of slow-moving chains compared to strains ΔpilA2 and ΔpilA3, resulting in a lower median velocity. Although strain ΔpilA1 generated a marginal twitching zone, most chains of ΔpilA1 expressed limited motility, confirming that PilA1 was critical for the biosynthesis of motile Tfp.
2.4. Wild-Type Tfp Were Essential for Maintaining Biofilm Structure
Previously, we found that the inactivation of pilT (the ∆pilT strain), which abolishes twitching activity, reduced biofilm formation in a flow-cell system where constant shear forces act upon the biofilm [14]. The decreased biofilm formation in ∆pilT may be due to interference from longer Tfp, compared to wild-type CGMH010, produced in the absence of PilT (Figure 2B). To further analyze the impact of twitching activity on biofilm formation, we examined the ability of the pilA mutant strains (described above) to form biofilms in a flow-cell system. Confocal laser scanning microscope (CLSM) examination of areas close to the medium intake port revealed that the thickness and structure of biofilms generated by the motile strains, i.e., ∆pilA2, ∆pilA3, and ∆pilA23, were comparable to those of wild-type CGMH010. In contrast, the biofilm derived from strains without active twitching motility, i.e., ∆pilA1, ∆pilA12, ∆pilA13, and ∆pilA123, was thinner compared to biofilms generated by wild-type CGMH010 (Figure 5). Thus, Tfp-mediated twitching motility is required for optimal biofilm formation under constant shearing forces.
2.5. Motile Tfp Are Essential for the Invasion of Host Cells by S. sanguinis
A recent study by Martini et al. indicated that Tfp mediate the invasion of endothelial cells by S. sanguinis SK36, a non-motile strain, suggesting that invasion is independent of twitching motility [21]. To investigate whether Tfp-mediated invasion is common to all S. sanguinis strains, and to identify the role of twitching motility in this process, we analyzed the invasion of human coronary artery endothelial (HCAEC) cells by S. sanguinis CGMH010 and its pilA- and pilT-deletion derivatives. S. sanguinis CGMH010 and SK36 invaded HCAEC cells at a similar rate in a PilA protein-dependent manner (Figure 6). Strains lacking pilT were drastically reduced in their ability to invade HCAEC cells. The ∆pilA1, ∆pilA2, and ∆pilA3 strains of CGMH010 displayed comparable levels of invasion, while conversely, the Tfp-null strain (∆pilA123) could not invade HCAEC, confirming that filament structure plays a role in invasion. Thus, the ability to retract is essential for Tfp-mediated invasion, regardless of whether a twitching zone could be detected on an agar surface.
3. Discussion
The study in S. sanguinis 2908 indicated that motile pili can be generated by a single major pilin protein, suggesting a redundancy of major pilin proteins in Tfp assembly; However, the degree of Tfp-mediated twitching motility is dependent upon the major pilin proteins that compose Tfp [20]. In contrast, our results demonstrate that the three PilA proteins expressed by S. sanguinis CGMH010 exhibit diverse impacts on the assembly and function of Tfp. Specifically, we show that PilA1 is most critical for Tfp production, and, although PilA1 and PilA3 share a high level of amino acid sequence identity, neither PilA3 alone nor a combination of PilA2 and PilA3 could fully compensate for the loss of pilA1. These results indicate that PilA1 and PilA3 function differently in Tfp biosynthesis. In addition, the results of this study support the hypothesis that Tfp-mediated motility is required for optimal biofilm formation under constant shearing force, and that PilT activity is essential for the Tfp-mediated invasion of host cells, regardless of whether strains could generate a twitching zone on an agar surface.
Recent reconstruction studies using cryo-electron microscopy on the major pilins of N. meningitidis, N. gonorrhoeae, and P. aeruginosa indicated that, rather than being continuous, the α-helix contains a melted region between two conserved helix-breaking residues, Gly14 and Pro22 [30,31]. It is proposed that the melting of this region could facilitate the integration of the pilin subunit into the growing Tfp, and a fully extended conformation of this region could allow the pilus to reversibly elongate up to three times its original length [30]. Thus, this region is likely to play a critical role in the assembly, extension, and retraction of Tfp. A comparison of all major pilin proteins from both S. sanguinis CGMH010 and S. sanguinis 2908 revealed that these major pilins share an identical sequence at the α1 region, except for the PilA2 of CGMH010. Although all major pilin proteins harbor Pro22, but not Gly14, PilA2 contains three conserved substitutions flanking Pro22. It is tempting to suggest that the altered amino acids may reduce the activity of PilA2 in the assembly of filaments, and, in fact, PilA2 had a minor effect on the assembly of retractable filaments.
Studies have shown that the primary sequence of the C-terminal region of the pilin protein, which contains sequences of an αβ-loop, a 4-stranded antiparallel β-sheet, and a variable D-loop, is less conserved among strains and species [4,28]. Using I-TASSER (https://zhanggroup.org/I-TASSER/, accessed on 16 February 2024), 4-, 2-, and 3-stranded β-sheets were predicted in PilA1, PilA2, and PilA3, respectively, suggesting that subtle differences in the structures could exist between these proteins. As the pil genes are arranged in an operon, and all three pilA genes contain a putative ribosomal binding site at an appropriate position, the different activities between PilA proteins in the assembly of Tfp should be related to the sequence, but not the expression level of the proteins. Similar to variations in Tfp-mediated twitching motility between S. sanguinis strains, a recent study in Acidovorax citrulli found that PilA proteins of strains belonging to different groups differ in sequence, and Tfp made of different PilA proteins exhibit distinct activities in twitching, biofilm formation, and even interspecies competitive abilities [32]. Furthermore, similar to the Tfp system of S. sanguinis, Haloferax volcanii produces multiple major pilins for Tfp biosynthesis, and Tfp composed of different major pilins exhibit different functions in the process of biofilm formation [33]. Thus, although PilA proteins are highly conserved in a S. sanguinis strain, the subtle sequence differences in the PilA proteins contribute significantly to the activity of the Tfp.
Unlike Gram-negative pathogens, in which Tfp-mediated twitching motility plays a critical role in virulence [4], past and current studies have revealed that Tfp, but not twitching motility, are more critical for S. sanguinis in its primary niche, the oral cavity. This notion is supported by the following observations: first, only a portion of the S. sanguinis isolates express twitching motility on an agar surface [14,34]; second, Tfp-mediated adherence [8,14] and invasion of host cells (this study) are observed in both motile and non-motile S. sanguinis strains. Interestingly, our recent sequence study on 30 clinical S. sanguinis strains from the Chang Gung Memorial Hospital (CGMH) bacteria bank found only one strain carrying two major pilin genes, indicating that strains encoding three major pilin genes are more prevalent. Like S. sanguinis 2908, the CGMH strain harboring two major pilin genes also expresses twitching motility. As the nature of the major pilins and the composition of Tfp could determine twitching activity, it is tempting to suggest that the major pilin gene underwent duplication during evolution, resulting in the production of three major pilin genes. The presence of the third PilA protein may lead to the production of Tfp expressing different levels of twitching motility. Since Tfp-mediated motility may not provide significant advantages for S. sanguinis to survive in the oral cavity, perhaps the non-motile Tfp evolved for energy conservation. On the other hand, a recent study in P. aeruginosa revealed that Tfp-mediated twitching motility promotes the surface departure of P. aeruginosa under reduced shearing forces. In contrast, high shearing forces enhance the adhesion of bacterial cells by counteracting Tfp retraction [35]. This observation raises the possibility that twitching motility serves different roles for S. sanguinis in the environment under high shearing forces, i.e., the bloodstream, and low shearing forces, i.e., the oral cavity. Our previous study on 81 clinical S. sanguinis CGMH strains revealed a total of 44 strains that could express twitching activity on either TH agar or blood agar, representing 54% of the isolates [14], which is higher than the incidence of the oral isolates reported by Henriksen et al. (49%) [34]. Thus, one could speculate that Tfp-mediated motility could provide additional benefits for S. sanguinis to establish outside the oral cavity.
Initially, we tried to determine the differences in the Tfp morphology between the pilA mutant strains by examining surface filaments on whole cells by TEM. However, S. sanguinis generates multiple surface hair-like structures, which is common in oral streptococci [36], and thus, we were unable to distinguish the differences in Tfp between S. sanguinis strains with confidence. On the other hand, although the pilT-deletion strain is routinely used as the non-motile control, the reduced invasion by ΔpilT may be caused by the lengthy Tfp, but not the motility per se. Nevertheless, loss of pilT abolished invasion by both motile (CGMH010) and non-motile (SK36) strains, and the ability to retract, but not the production of a twitching zone, is critical for Tfp-mediated invasion by S. sanguinis. Similarly, as non-motile *pilA-*knockout strains of CGMH010 did not express long Tfp, the reduced biofilm formation of these strains was unrelated to the length of the Tfp.
The significance of expressing multiple highly homologous major pilin proteins in S. sanguinis is yet undefined, although sequence variations and post-translational modification of the pilin proteins may contribute to antigenic variation and immune evasion [30,37,38]. However, the antigenic variation from the highly identical amino acid sequences of the PilA isomers, especially PilA1 and PilA3, remains to be elucidated. The results from this study suggest that Tfp provide competitive advantages for S. sanguinis, similar to their role in Gram-negative pathogens, and further demonstrate that the nature of the major pilin proteins plays an essential role in the twitching activity of S. sanguinis CGMH010.
4. Materials and Methods
4.1. Bacterial Strains and Culture Conditions
The bacterial strains and plasmids used in this study are listed in Table 1. S. sanguinis strains were routinely cultivated in Todd Hewitt (TH) medium at 37 °C in a 10% CO_2_ atmosphere or in an anaerobic jar. Where indicated, erythromycin (Em) at 10 μg mL^−1^ or kanamycin (Km) at 500 μg mL^−1^ was included for the selection of recombinant S. sanguinis strains. Recombinant E. coli strains were cultivated in LB broth containing Km at 50 µg mL^−1^, with continuous shaking at 37 °C.
4.2. Generation of pilA Knockout Derivatives of S. sanguinis CGMH010
All mutant derivatives of S. sanguinis CGMH010 were generated by ligation mutagenesis [40]. The primers used in this study are listed in Table S1. To generate a specific knockout, two fragments, 5′ and 3′ to the target gene, respectively, were generated by PCR with specific primers. Restriction endonuclease recognition sequences were included in the primers to facilitate subsequent ligation reactions. The PCR products were digested and mixed with DNA fragments containing the non-polar Em resistance gene (erm) [41] or non-polar Km resistance gene (kan) [42] in a reaction that allows for the ligation of the 5′ flanking fragment, followed by the erm or kan fragment and then the 3′ flanking fragment. For the generation of strains ΔpilA1, ΔpilA2, ΔpilA3, ΔpilA12, ΔpilA23, and ΔpilA123, the ligation mixture was used to transform S. sanguinis CGMH010 [43] with selection for Em resistance. The ΔpilA13 strain was generated by transforming a ligation mixture containing the pilA3 inactivation construct into the ΔpilA1 strain and selecting for Km resistance. The allelic exchange event in the isogenic knockout strains was confirmed by colony PCR with primers located outside the erm or kan insertion site with specific primers.
4.3. Expression of the Pilin Proteins and Preparation of Pilin Protein-Specific Antibody
An anti-PilA1 antibody was generated in rabbits immunized with an internal peptide of PilA1 containing amino acid residues 95 to 104 (AKGGDKIADA) (Figure 1, red box). The peptide and an antiserum specific to the peptide were generated commercially (GenScript, Piscataway, NJ, USA).
A histidine-tagged PilA2 protein (His-PilA2) was constructed using the pET28a(+) expression plasmid (Novagen, Darmstadt, Germany), and purified His-PilA2 was used for anti-PilA2 antiserum production in rabbits. Briefly, the coding sequence for amino acids 38 to 152 of PilA2 was amplified from S. sanguinis CGMH010 by PCR (primer sequences in Table S1), cloned into pET28a(+), and established in E. coli BL21. The sequence of the recombinant plasmid was verified by sequencing. Induction by IPTG and the purification of His-PilA2 by Ni^2+^ affinity chromatography were performed under denaturing conditions using the standard procedure (Qiagen, Hilden, Germany). The identity of the purified His-PilA2 protein was confirmed by Matrix-Assisted Laser Desorption/Ionization–Time of Flight Mass Spectrometry. Approximately 2.5 mg of purified His-PilA2 protein was separated on 12% SDS-PAGE gel, and the expected protein band was excised from the gel and used to generate polyclonal antiserum in the rabbits (LTK BioLaboratories, Taoyuan, Taiwan).
A similar approach was used to generate a histidine-tagged PilA3 (His-PilA3). The coding sequence for amino acid residues 38 to 148 of PilA3 (primer sequences in Table S1) was cloned in plasmid pET28a(+), expressed, purified, and confirmed as described above, and then used for antiserum production in rabbits.
To examine the specificity and titer of the anti-PilA1 antiserum, a histidine-tagged PilA1 protein (amino acid residues 38 to 147; His-PilA1) was also prepared using the same approach.
4.4. Protein Structure Prediction
The 3D structures of the PilA proteins were predicted by using the AlphaFold protein structure database (https://alphafold.ebi.ac.uk, accessed on 29 April 2024).
4.5. Purification of Extracellular Tfp
Extracellular Tfp of S. sanguinis strains were purified as previously described with minor modifications [20]. Briefly, cultures (100 mL) of S. sanguinis strains were grown in TH medium to an O.D.600 of 0.8, harvested, washed once with 10 mM NaPO_4_, and then concentrated 100-fold in pilus buffer (20 mM Tris-HCl [pH 7.6], 50 mM NaCl). The concentrated suspensions were vortexed at full speed for 2 min to shear Tfp. At the end of the shearing step, the suspensions were centrifuged at 6000× g for 10 min and the supernatant was recovered. The sheared filaments in the supernatant were recovered by ultra-centrifugation at 100,000× g, 4 °C, for 1 h. The final pellet was dissolved in 60 μL pilus buffer. A total of 15 μL of the final suspension was separated on 12% SDS-PAGE, and 5 μL of the preparation was used for TEM examination.
4.6. Immunogold Labeling, Negative Staining, and TEM Observation
For transmission electron microscopy (TEM) examination, 5 μL of the extracellular Tfp preparation was applied onto a 200-mesh carbon-coated copper grid (Agar, Essex, UK) and allowed to set for 30 s. The sample was subjected to negative staining directly or immunogold labeling with anti-PilA antibody, followed by negative staining. Negative staining of Tfp preparations was performed by adding 5 μL 2% methylamine tungstate (Nanoprobes, New York, NY, USA) to the sample, allowing it to set for 10 s, and then placing the sample in a moisture-proof box (44% relative humidity) for 24 h before TEM examination. The grid was observed using a JEM-2100 Plus electron microscope (JEOL, Japan, Tokyo).
For immunogold labeling, the sample was fixed on the grid with 10 μL of a fixing solution composed of 0.2% glutaraldehyde and 4% paraformaldehyde at room temperature for 5 min. At the end of the fixing reaction, the grid was washed three times with pilus buffer to remove the fixing solution. The sample was then blocked with TBST (150 mM NaCl, 50 mM Tris [pH 7.6], 0.5% Tween 20) containing 3% BSA at room temperature for 30 min, followed by incubation with anti-PilA2 or anti-PilA3 antiserum at 1:10 dilution in TBST for 1 h. The unbound antibody was removed by rinsing with drops of the pilus buffer. The bound antibody was recognized by goat anti-rabbit IgG conjugate with 10 nm gold particles (Sigma, St. Louis, MI, USA) at room temperature for 1 h. The grid was again rinsed with pilus buffer 10 times to remove the secondary antibody prior to negative staining and TEM examination.
4.7. Gel Electrophoresis and Western Blot Analysis
Protein preparations were separated on 12% SDS-PAGE for Western blot analysis. The separated proteins were transferred from gels to PVDF membranes (Merck, Darmstadt, Germany). After blocking with 5% skim milk in TBST at 4 °C overnight, the blot was hybridized with antibodies specific to each of the pilin proteins in TBST containing 5% skim milk for 1 h at room temperature. The anti-PilA1 antibody was used at 1:1000; the anti-PilA2 and anti-PilA3 antisera were used at 1:5000. Horseradish peroxidase-conjugated anti-rabbit antibody (GeneTex, Irvine, CA, USA) and luminol-based Immobilon western chemiluminescent HRP substrate (Millipore, Darmstadt, Germany) were used to detect bound PilA-specific antisera. The results were imaged using a UVP BioSpectrum Imaging system (Vilber, Marne-la-Vallée, France).
4.8. Examination of Twitching Motility
Twitching motility was examined macroscopically on TH agar (1%) as previously described [14]. Briefly, S. sanguinis strains were inoculated in straight lines on TH agar (1%) and incubated in an anaerobic jar with 30 mL water added to the bottom of the jar (to maintain humidity) at 37 °C for 66 h.
Wet-mount preparations were used for microscopic examination of twitching motility, as previously described, with minor modifications [14]. An 8 μL aliquot of log-phase cultures (O.D.600 = 0.4) of S. sanguinis strains grown in TH medium was used to prepare the wet-mount. The wet-mount sample was incubated at 37 °C in a 10% CO_2_ atmosphere for 30 min before microscopic examination at 100× magnification (Olympus BX41, Tokyo, Japan). The movement of the bacteria was recorded for 10 s using a TrueChrome AF microscope camera (Tucsen, Fuzhou, China). The velocity of bacterial movement was analyzed using ImageJ (https://imagej.net/) with a particle-tracking plugin (MultiTracker). The velocity distribution is presented as a violin plot created using GraphPad Prism 7.
4.9. Preparation of Flow-Cell Biofilm and Examination by CLSM
Biofilms in the flow-cell system were prepared as previously described [14]. Briefly, overnight cultures of S. sanguinis strains grown in biofilm medium (BM) [44] containing 40 mM glucose (BMG) were diluted in BMG to an O.D.600 of 0.4. A total of 300 μL of the diluted culture was injected into the growth chamber. The chamber was kept upside down without medium flow for 4 h at 37 °C. The medium flow was set to 5 mL^−1^ h^−1^. The chamber was incubated for 5 h at 37 °C. At the end of the incubation, the biofilms were stained with SYTO 9 and propidium iodide (PI) for 30 min using a LIVE/DEAD biofilm viability kit (Invitrogen, Carlsbad, CA, USA). After staining, the samples were washed with 500 μL double-distilled H_2_O. The stained biofilms were examined by confocal laser scanning microscopy (CLSM) with a 63× oil immersion objective lens (LSM780, Zeiss, Jena, Germany). The images were displayed by the ZEN acquisition software (Zen 2012 SP5 FP3 black, Zeiss).
4.10. Invasion Assay
Primary human coronary artery endothelial cells (HCAEC, ATCC, Manassas, VA, USA) were grown in vascular cell basal medium (ATCC) supplemented with an Endothelial Cell Growth Kit—VEGF (ATCC). HCAEC cells were seeded in 24-well plates and grown for 12 h before the assay. Cells were infected with exponential-phase cultures (O.D.600 = 0.4) of S. sanguinis strains at an MOI of 20 for 2 h. At the end of incubation, the extracellular bacteria were removed by washing with PBS twice, followed by a penicillin (100 units mL^−1^)–gentamicin (300 μg mL^−1^)–streptomycin (100 μg mL^−1^) treatment at 37 °C for 1 h. At the end of the treatment, the cells were washed with PBS twice, then lysed with 0.1% Triton X-100 for 10 min to recover the intracellular bacteria. Intracellular bacterial counts were determined by serial dilution and plating. The invasion rate was calculated as (CFU recovered/CFU of the inoculum) × 100%. Significant differences between strains were analyzed using one-way ANOVA followed by Tukey’s test. Differences were considered significant if p < 0.01.
5. Conclusions
Multiple major pilin proteins are commonly observed in S. sanguinis Tfp. Although the major pilin proteins share high levels of identity within a strain, presumably as a result of gene duplication, these pilin proteins contribute differently to the assembly of motile Tfp. As the Tfp-mediated adherence and invasion of host cells are observed in both motile and non-motile S. sanguinis strains, Tfp-mediated twitching motility may provide limited advantages for S. sanguinis competitiveness. Thus, the production of various major pilin proteins may allow S. sanguinis to modulate twitching motility for energy conservation.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Denise R. Abby S.S. Rocha E.P.C. Diversification of the type IV filament superfamily into machines for adhesion, protein secretion, DNA uptake, and motility P Lo S Biol.201917 e 300039010.1371/journal.pbio.300039031323028 PMC 6668835 · doi ↗ · pubmed ↗
- 2Daum B. Gold V. Twitch or swim: Towards the understanding of prokaryotic motion based on the type IV pilus blueprint Biol. Chem.201839979980810.1515/hsz-2018-015729894297 · doi ↗ · pubmed ↗
- 3Giltner C.L. Nguyen Y. Burrows L.L. Type IV pilin proteins: Versatile molecular modules Microbiol. Mol. Biol. Rev. MMBR 20127674077210.1128/MMBR.00035-1223204365 PMC 3510520 · doi ↗ · pubmed ↗
- 4Craig L. Forest K.T. Maier B. Type IV pili: Dynamics, biophysics and functional consequences Nat. Rev. Microbiol.20191742944010.1038/s 41579-019-0195-430988511 · doi ↗ · pubmed ↗
- 5Pelicic V. Type IV pili: E pluribus unum?Mol. Microbiol.20086882783710.1111/j.1365-2958.2008.06197.x 18399938 · doi ↗ · pubmed ↗
- 6Bordeleau E. Purcell E.B. Lafontaine D.A. Fortier L.C. Tamayo R. Burrus V. Cyclic di-GMP riboswitch-regulated type IV pili contribute to aggregation of Clostridium difficile J. Bacteriol.201519781983210.1128/JB.02340-1425512308 PMC 4325102 · doi ↗ · pubmed ↗
- 7Varga J.J. Nguyen V. O’Brien D.K. Rodgers K. Walker R.A. Melville S.B. Type IV pili-dependent gliding motility in the Gram-positive pathogen Clostridium perfringens and other Clostridia Mol. Microbiol.20066268069410.1111/j.1365-2958.2006.05414.x 16999833 · doi ↗ · pubmed ↗
- 8Chen Y.M. Chiang Y.C. Tseng T.Y. Wu H.Y. Chen Y.Y. Wu C.H. Chiu C.H. Molecular and functional analysis of the type IV pilus gene cluster in Streptococcus sanguinis SK 36Appl. Environ. Microbiol.201985 e 02788-1810.1128/AEM.02788-1830635384 PMC 6414370 · doi ↗ · pubmed ↗