The genome sequence of the Grey Sedge caddis fly, Odontocerum albicorne (Scopoli, 1769)
Sue Skipp, Ian Wallace, Kevin M Moran, John Soghigian, Huiqing Yeo

TL;DR
This paper reports the genome sequence of the Grey Sedge caddis fly, including a detailed assembly of its chromosomes and mitochondrial DNA.
Contribution
The novel contribution is the first genome assembly for Odontocerum albicorne, including scaffolded chromosomal pseudomolecules and the mitochondrial genome.
Findings
The genome assembly spans 1,287.3 megabases and is scaffolded into 31 chromosomal pseudomolecules.
The mitochondrial genome is 16.57 kilobases in length and has been fully assembled.
Abstract
We present a genome assembly from an individual male Odontocerum albicorne (the Grey Sedge caddis fly; Arthropoda; Insecta; Trichoptera; Odontoceridae). The genome sequence is 1,287.3 megabases in span. Most of the assembly is scaffolded into 31 chromosomal pseudomolecules, including the Z sex chromosome. The mitochondrial genome has also been assembled and is 16.57 kilobases in length.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1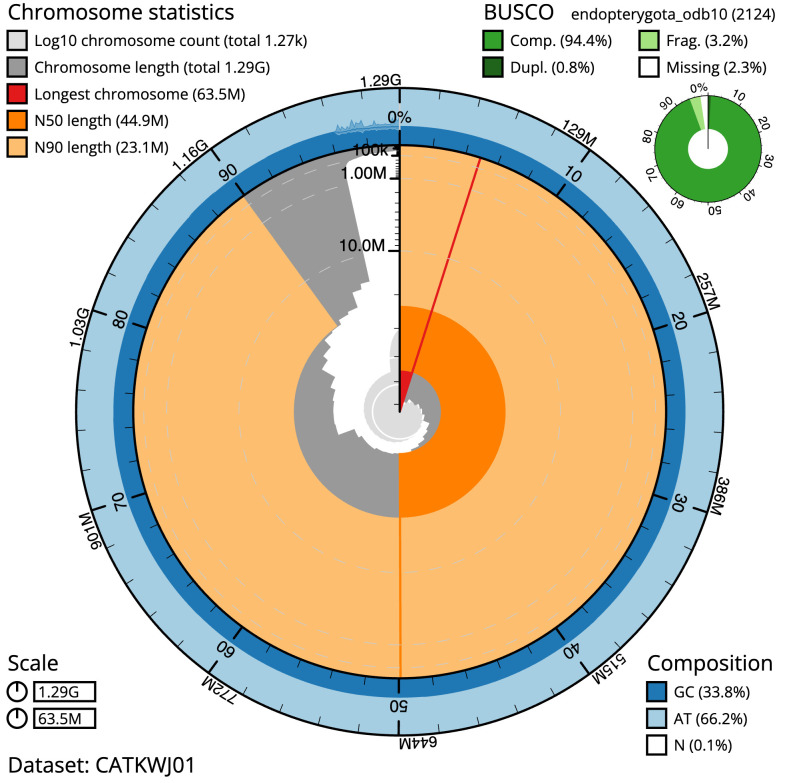 Figure 2
Figure 2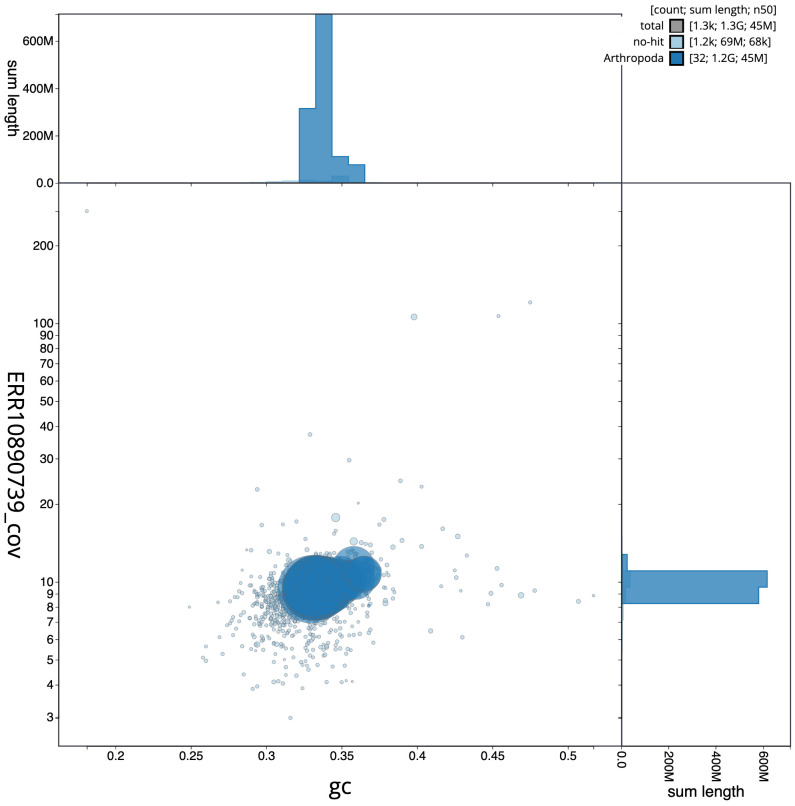 Figure 3
Figure 3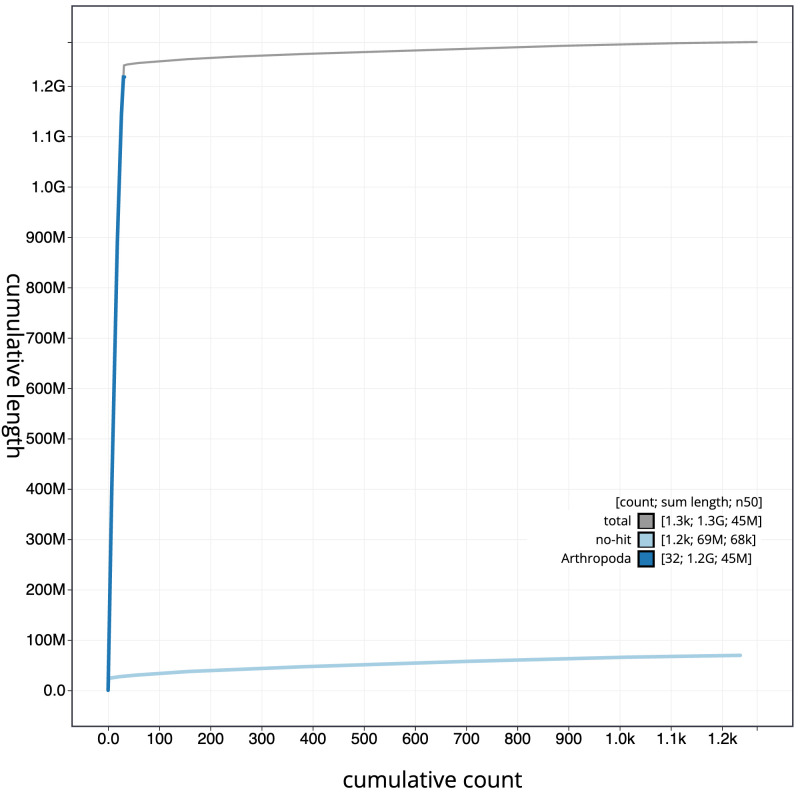 Figure 4
Figure 4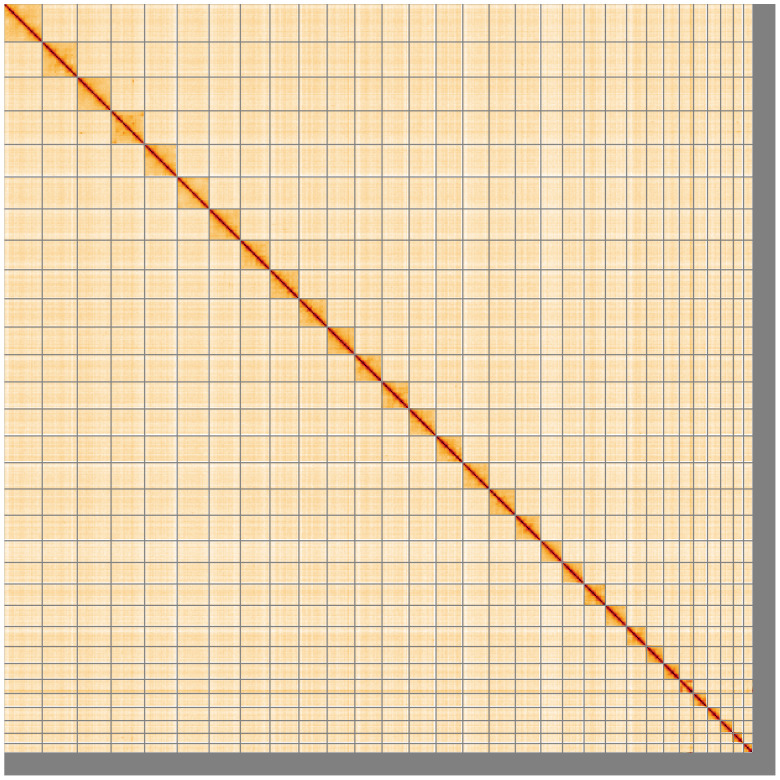 Figure 5
Figure 5| Project accession data | ||
|---|---|---|
| Assembly identifier | iiOdoAlbi1.1 | |
| Species |
| |
| Specimen | iiOdoAlbi1 | |
| NCBI taxonomy ID | 446452 | |
| BioProject | PRJEB59794 | |
| BioSample ID | SAMEA7520808 | |
| Isolate information | iiOdoAlbi1, male: terminal body (DNA and Hi-C sequencing) | |
| Assembly metrics
|
| |
| Consensus quality (QV) | 57.7 |
|
|
| 99.99% |
|
| BUSCO
| C:94.4%[S:93.6%,D:0.8%],
|
|
| Percentage of assembly
| 96.62% |
|
| Sex chromosomes | Z chromosome |
|
| Organelles | Mitochondrial genome assembled |
|
| Raw data accessions | ||
| PacificBiosciences SEQUEL II | ERR10880461, ERR10879934 | |
| Hi-C Illumina | ERR10890741 | |
| Genome assembly | ||
| Assembly accession | GCA_949825065.1 | |
|
| GCA_949825055.1 | |
| Span (Mb) | 1,287.3 | |
| Number of contigs | 6038 | |
| Contig N50 length (Mb) | 0.4 | |
| Number of scaffolds | 1267 | |
| Scaffold N50 length (Mb) | 44.9 | |
| Longest scaffold (Mb) | 63.5 | |
| INSDC accession | Chromosome | Length (Mb) | GC% |
|---|---|---|---|
| 1 | 63.5 | 33.0 | |
| 2 | 58.15 | 33.5 | |
| 4 | 55.89 | 33.5 | |
| 3 | 55.65 | 33.5 | |
| 5 | 53.85 | 33.0 | |
| 6 | 51.78 | 33.0 | |
| 7 | 48.92 | 33.5 | |
| 8 | 48.05 | 33.0 | |
| 9 | 46.77 | 33.5 | |
| 10 | 45.87 | 34.0 | |
| 11 | 45.0 | 33.5 | |
| 12 | 44.9 | 33.0 | |
| 13 | 44.53 | 33.5 | |
| 14 | 44.34 | 33.5 | |
| 16 | 43.56 | 33.5 | |
| 15 | 43.56 | 33.5 | |
| 17 | 42.23 | 34.0 | |
| 18 | 35.88 | 34.0 | |
| 20 | 35.61 | 34.5 | |
| 19 | 35.53 | 34.0 | |
| 21 | 34.99 | 34.0 | |
| 22 | 33.13 | 34.5 | |
| 23 | 28.09 | 35.0 | |
| 24 | 26.23 | 35.0 | |
| 25 | 23.51 | 36.0 | |
| 26 | 23.06 | 35.0 | |
| 27 | 21.7 | 35.5 | |
| 28 | 21.04 | 36.0 | |
| 29 | 16.94 | 36.5 | |
| 30 | 15.61 | 36.5 | |
| Z | 52.95 | 33.0 | |
| MT | 0.02 | 18.0 |
| Software tool | Version | Source |
|---|---|---|
| BlobToolKit | 4.1.7 |
|
| BUSCO | 5.3.2 |
|
| gEVAL | N/A |
|
| Hifiasm | 0.16.1-r375 |
|
| HiGlass | 1.11.6 |
|
| Merqury | MerquryFK |
|
| MitoHiFi | 2 |
|
| PretextView | 0.2 |
|
| purge_dups | 1.2.3 |
|
| sanger-tol/genomenote | v1.0 |
|
| sanger-tol/readmapping | 1.1.0 |
|
| YaHS | 1.2a |
|
- —Wellcome Trust
- —Wellcome Trust
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGenomics and Phylogenetic Studies · Environmental DNA in Biodiversity Studies · Chromosomal and Genetic Variations
Species taxonomy
Eukaryota; Metazoa; Eumetazoa; Bilateria; Protostomia; Ecdysozoa; Panarthropoda; Arthropoda; Mandibulata; Pancrustacea; Hexapoda; Insecta; Dicondylia; Pterygota; Neoptera; Endopterygota; Amphiesmenoptera; Trichoptera; Integripalpia; Brevitentoria; Leptoceroidea; Odontoceridae; Odontocerinae; Odontocerum; Odontocerum albicorne (Scopoli, 1769) (NCBI:txid446452).
Background
The adult of Odontocerum albicorne ( Figure 1) has the common name of Silver or Grey Sedge, although the male has yellowish wings. Those are held partially rolled around the body when at rest. Adults also have toothed antennae, but the purpose of that is not known. Larvae live in permanent, moderate to fast flowing, streams and rivers on a substratum of stones sand and gravel. The species is absent from Shetland and the Outer Hebrides and scarce in East Anglia due to lack of suitable flowing waters. It is the only member of its family in Britain and Ireland.
Photograph of adult Odontocerum albicorne (not the specimen used for genome sequencing) by Janet Graham, CC-BY.
The distinctive adults can be easily identified in the wild, with the assistance of Wallace et al. (2022). The curved sand grain cases with their unique posterior closure are also easy to recognise and the pupal cases persist for some time after the adult has emerged. Larger larvae have a characteristic anchor mark on their pale heads. Larvae of all sizes can be identified using Wallace et al., 2003).
There is consensus that in Britain the life cycle takes one year but due to a long flight period, from May to September, the larvae are at various sizes throughout. Eggs are laid by females dropping an egg mass into slowly flowing water. The egg absorbs water and expands so the final jelly egg mass, which has a sculptured outer surface, is between 8 and 10 mm in diameter. The eggs do not have any delayed hatching.
The larva spends the day under stones and forages, especially in the early part of the night. Out of preference it is predaceous and eats animals it can subdue or find dead; it has been reported feeding on the eggs of the Bullhead Cottus gobio Linnaeus. However, it will also eat plant material if higher energy food is not available. The larva has several interesting features relating to its method of case construction. Other cased caddis join case particles with silk threads and then line the resulting tube with a layer of smooth silk. O. albicorne larvae firmly cement the outer case particles together with silk material dispensing with an internal silk lining. The case is very strong and can resist attack from some biting fish. However, if bent, it will snap rather than deform. Other caddis larvae have sharply pointed anal proleg claws that can grip the internal silk lining of the case and help prevent the larva being forcibly removed, but the claws of O. albicorne are blunt, with the result that they grip the knobbly inner surface of the case better. The posterior of the case is closed, with a large sand grain attached to the rim of the case, with holes in the attachment to allow for water movement.
This is one of a handful of caddis larvae that have an ichneumonid parasite. Agriotypus armatus Curtis normally attacks goerid caddis pupae, but has this species as an alternative host ( Grenier, 1970).
The genome of O. albicorne was sequenced as part of the Darwin Tree of Life Project, a collaborative effort to sequence all named eukaryotic species in the Atlantic Archipelago of Britain and Ireland.
Genome sequence report
The genome was sequenced from one Odontocerum albicorne ( Figure 1) collected from River Itchen, Itchen Stoke, UK (51.09, –1.20). A total of 25-fold coverage in Pacific Biosciences single-molecule HiFi long reads was generated. Primary assembly contigs were scaffolded with chromosome conformation Hi-C data. Manual assembly curation corrected 329 missing joins or mis-joins and removed 24 haplotypic duplications, reducing the assembly length by 0.15% and the scaffold number by 7.04%, and increasing the scaffold N50 by 1.45%.
The final assembly has a total length of 1,287.3 Mb in 1267 sequence scaffolds with a scaffold N50 of 44.9 Mb ( Table 1). A summary of the assembly statistics is shown in Figure 2, while the distribution of assembly scaffolds on GC proportion and coverage is shown in Figure 3. The cumulative assembly plot in Figure 4 shows curves for subsets of scaffolds assigned to different phyla. Most (96.62%) of the assembly sequence was assigned to 10 chromosomal-level scaffolds, representing 30 autosomes and the Z sex chromosome. Chromosome Z was assigned by synteny to Glyphotaelius pellucidus (GCA_936435175.1) ( McSwan et al., 2023). Chromosome-scale scaffolds confirmed by the Hi-C data are named in order of size ( Figure 5; Table 2). While not fully phased, the assembly deposited is of one haplotype. Contigs corresponding to the second haplotype have also been deposited. The mitochondrial genome was also assembled and can be found as a contig within the multifasta file of the genome submission.
Table 1.: Genome data for Odontocerum albicorne, iiOdoAlbi1.1.
Genome assembly of Odontocerum albicorne, iiOdoAlbi1.1: metrics.The BlobToolKit Snailplot shows N50 metrics and BUSCO gene completeness. The main plot is divided into 1,000 size-ordered bins around the circumference with each bin representing 0.1% of the 1,287,343,752 bp assembly. The distribution of scaffold lengths is shown in dark grey with the plot radius scaled to the longest scaffold present in the assembly (63,503,693 bp, shown in red). Orange and pale-orange arcs show the N50 and N90 scaffold lengths (44,898,628 and 23,064,039 bp), respectively. The pale grey spiral shows the cumulative scaffold count on a log scale with white scale lines showing successive orders of magnitude. The blue and pale-blue area around the outside of the plot shows the distribution of GC, AT and N percentages in the same bins as the inner plot. A summary of complete, fragmented, duplicated and missing BUSCO genes in the endopterygota_odb10 set is shown in the top right An interactive version of this figure is available at https://blobtoolkit.genomehubs.org/view/Odontocerum%20albicorne/dataset/CATKWJ01/snail.
Genome assembly of Odontocerum albicorne, iiOdoAlbi1.1: BlobToolKit GC-coverage plot.Scaffolds are coloured by phylum. Circles are sized in proportion to scaffold length. Histograms show the distribution of scaffold length sum along each axis. An interactive version of this figure is available at https://blobtoolkit.genomehubs.org/view/Odontocerum%20albicorne/dataset/CATKWJ01/blob.
Genome assembly of Odontocerum albicorne, iiOdoAlbi1.1: BlobToolKit cumulative sequence plot.The grey line shows cumulative length for all scaffolds. Coloured lines show cumulative lengths of scaffolds assigned to each phylum using the buscogenes taxrule. An interactive version of this figure is available at https://blobtoolkit.genomehubs.org/view/Odontocerum%20albicorne/dataset/CATKWJ01/cumulative.
Genome assembly of Odontocerum albicorne, iiOdoAlbi1.1: Hi-C contact map of the iiOdoAlbi1.1 assembly, visualised using HiGlass.Chromosomes are shown in order of size from left to right and top to bottom. An interactive version of this figure may be viewed at https://genome-note-higlass.tol.sanger.ac.uk/l/?d=QqP4gJm9SsCzzU461RBsHA.
Table 2.: Chromosomal pseudomolecules in the genome assembly of Odontocerum albicorne, iiOdoAlbi1.
The estimated Quality Value (QV) of the final assembly is 57.7 with k-mer completeness of 99.99%, and the assembly has a BUSCO v5.3.2 completeness of 94.4% (single = 93.6%, duplicated = 0.8%), using the endopterygota_odb10 reference set ( n = 2,124).
Metadata for specimens, spectral estimates, sequencing runs, contaminants and pre-curation assembly statistics can be found at https://links.tol.sanger.ac.uk/species/446452.
Methods
Sample acquisition and nucleic acid extraction
A male Odontocerum albicorne (specimen ID NHMUK014361308, individual iiOdoAlbi1) was collected from River Itchen, Itchen Stoke, UK (latitude 51.09, longitude –1.20) on 2019-03-19 using a kicknet. The specimen was collected by Sue Skipp (Environment Agency) and identified by Ian Wallace, and then snap-frozen using a dry shipper.
The iiOdoAlbi1 sample was prepared for DNA extraction at the Tree of Life laboratory, Wellcome Sanger Institute (WSI). The specimen was weighed and dissected on dry ice with tissue set aside for Hi-C sequencing. Head and thorax tissue was disrupted using a Nippi Powermasher fitted with a BioMasher pestle. DNA was extracted at the WSI Scientific Operations core using the Qiagen MagAttract HMW DNA kit, according to the manufacturer’s instructions.
Sequencing
Pacific Biosciences HiFi circular consensus DNA sequencing libraries were constructed according to the manufacturers’ instructions. DNA sequencing was performed by the Scientific Operations core at the WSI on a Pacific Biosciences SEQUEL II (HiFi) instrument. Hi-C data were also generated from terminal body tissue of iiOdoAlbi1 using the Arima2 kit and sequenced on the HiSeq X Ten instrument.
Genome assembly, curation and evaluation
Assembly was carried out with Hifiasm ( Cheng et al., 2021) and haplotypic duplication was identified and removed with purge_dups ( Guan et al., 2020). The assembly was then scaffolded with Hi-C data ( Rao et al., 2014) using YaHS ( Zhou et al., 2023). The assembly was checked for contamination and corrected using the gEVAL system ( Chow et al., 2016) as described previously ( Howe et al., 2021). Manual curation was performed using gEVAL, HiGlass ( Kerpedjiev et al., 2018) and Pretext ( Harry, 2022). The mitochondrial genome was assembled using MitoHiFi ( Uliano-Silva et al., 2023), which runs MitoFinder ( Allio et al., 2020) or MITOS ( Bernt et al., 2013) and uses these annotations to select the final mitochondrial contig and to ensure the general quality of the sequence.
A Hi-C map for the final assembly was produced using bwa-mem2 ( Vasimuddin et al., 2019) in the Cooler file format ( Abdennur & Mirny, 2020). To assess the assembly metrics, the k-mer completeness and QV consensus quality values were calculated in Merqury ( Rhie et al., 2020). This work was done using Nextflow ( Di Tommaso et al., 2017) DSL2 pipelines “sanger-tol/readmapping” ( Surana et al., 2023a) and “sanger-tol/genomenote” ( Surana et al., 2023b). The genome was analysed within the BlobToolKit environment ( Challis et al., 2020) and BUSCO scores ( Manni et al., 2021; Simão et al., 2015) were calculated.
Table 3 contains a list of relevant software tool versions and sources.
Wellcome Sanger Institute – Legal and Governance
The materials that have contributed to this genome note have been supplied by a Darwin Tree of Life Partner. The submission of materials by a Darwin Tree of Life Partner is subject to the ‘Darwin Tree of Life Project Sampling Code of Practice’, which can be found in full on the Darwin Tree of Life website here. By agreeing with and signing up to the Sampling Code of Practice, the Darwin Tree of Life Partner agrees they will meet the legal and ethical requirements and standards set out within this document in respect of all samples acquired for, and supplied to, the Darwin Tree of Life Project.
Further, the Wellcome Sanger Institute employs a process whereby due diligence is carried out proportionate to the nature of the materials themselves, and the circumstances under which they have been/are to be collected and provided for use. The purpose of this is to address and mitigate any potential legal and/or ethical implications of receipt and use of the materials as part of the research project, and to ensure that in doing so we align with best practice wherever possible. The overarching areas of consideration are:
• Ethical review of provenance and sourcing of the material
• Legality of collection, transfer and use (national and international)
Each transfer of samples is further undertaken according to a Research Collaboration Agreement or Material Transfer Agreement entered into by the Darwin Tree of Life Partner, Genome Research Limited (operating as the Wellcome Sanger Institute), and in some circumstances other Darwin Tree of Life collaborators.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Abdennur N Mirny LA : Cooler: Scalable storage for Hi-C data and other genomically labeled arrays. Bioinformatics. 2020;36(1):311–316. 10.1093/bioinformatics/btz 540 31290943 PMC 8205516 · doi ↗ · pubmed ↗
- 2Allio R Schomaker‐Bastos A Romiguier J : Mito Finder: Efficient automated large‐scale extraction of mitogenomic data in target enrichment phylogenomics. Mol Ecol Resour. 2020;20(4):892–905. 10.1111/1755-0998.13160 32243090 PMC 7497042 · doi ↗ · pubmed ↗
- 3Bernt M Donath A Jühling F : MITOS: Improved de novo metazoan mitochondrial genome annotation. Mol Phylogenet Evol. 2013;69(2):313–319. 10.1016/j.ympev.2012.08.023 22982435 · doi ↗ · pubmed ↗
- 4Challis R Richards E Rajan J : Blob Tool Kit - interactive quality assessment of genome assemblies. G 3 (Bethesda). 2020;10(4):1361–1374. 10.1534/g 3.119.400908 32071071 PMC 7144090 · doi ↗ · pubmed ↗
- 5Cheng H Concepcion GT Feng X : Haplotype-resolved de novo assembly using phased assembly graphs with hifiasm. Nat Methods. 2021;18(2):170–175. 10.1038/s 41592-020-01056-5 33526886 PMC 7961889 · doi ↗ · pubmed ↗
- 6Chow W Brugger K Caccamo M : g EVAL — a web-based browser for evaluating genome assemblies. Bioinformatics. 2016;32(16):2508–10. 10.1093/bioinformatics/btw 159 27153597 PMC 4978925 · doi ↗ · pubmed ↗
- 7Di Tommaso P Chatzou M Floden EW : Nextflow enables reproducible computational workflows. Nat Biotechnol. 2017;35(4):316–319. 10.1038/nbt.3820 28398311 · doi ↗ · pubmed ↗
- 8Grenier S : Biologie d’ Agriotypus armatus Curtis (Hymenoptera: Agriotypidae), parasite de nymphes de Trichoptères. Ann Limnol. 1970;6(3):317–361. 10.1051/limn/1970010 · doi ↗