Genetic diversity and haplotype analysis of cattle hydatid cyst isolates using mitochondrial markers in Kazakhstan
Rabiga Uakhit, Sofiya Yalysheva, Aida Abdybekova, Ainura Smagulova, Lyudmila Lider, Karina Jazina, Aidana Tautanova, Vladimir Kiyan

TL;DR
This study identifies the genetic diversity of Echinococcus in cattle in Kazakhstan, finding that the G1 genotype is most common.
Contribution
The study provides new insights into the haplotype diversity and genetic variation of Echinococcus in Kazakhstani cattle using mitochondrial markers.
Findings
All isolates were identified as Echinococcus granulosus s.s., with 94.5% being the G1 genotype.
38 distinct haplotypes were identified through bioinformatic analysis of mitochondrial gene sequences.
Only one region showed coexistence of G1 and G3 genotypes, suggesting regional epidemiological differences.
Abstract
In Kazakhstan, the study of Echinococcus infection among farm animals is crucial to monitor the invasion among livestock and map the data obtained. Unfortunately, there are only partial data on the study of Echinococcus among cattle’s in Kazakhstan, which makes it difficult to conduct a comparative analysis of the epidemiological situation among livestock animals. The present study aimed to molecularly identify the species and haplotypes of the E. granulosus complex infecting cattle in Kazakhstan and investigate their genetic variation relative to mitochondrial (mt) targets. Individual cyst isolates (n = 700) were collected from infected cattle lungs and livers after slaughter from the slaughterhouse. Total DNA was extracted from the germinal layers of the cyst from each isolate. This DNA sequenced partial mt genes of cytochrome c oxidase 1 (450 bp) and NADH dehydrogenase 1 (1200 bp).…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
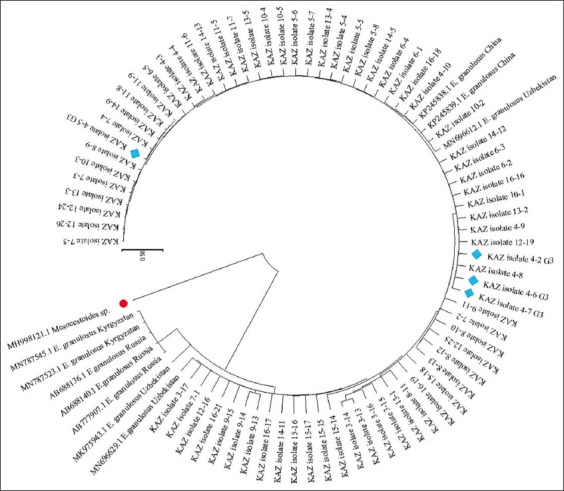 Figure-1
Figure-1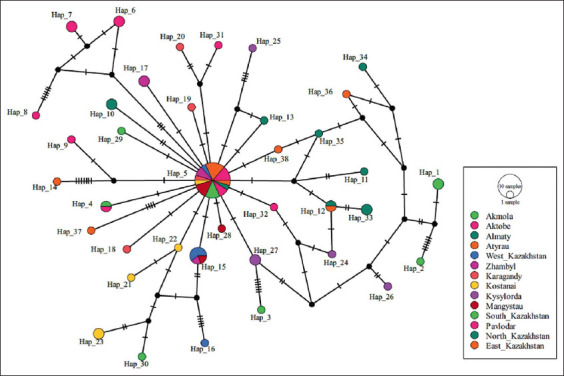 Figure-2
Figure-2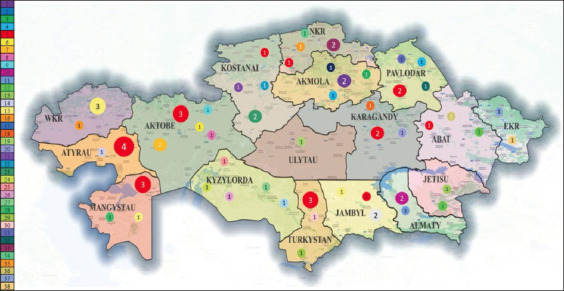 Figure-3
Figure-3| Geographical origin | Isolate | GB number (co×1) | GB number (nad1) | Genotype | Haplotype |
|---|---|---|---|---|---|
| Akmola region | 3-22-13 | G1 | hp1 | ||
| 3-22-14 | G1 | hp2 | |||
| 3-22-15 | G1 | hp3 | |||
| 3-22-16 | G1 | hp1 | |||
| 3-22-17 | G1 | hp4 | |||
| Aktobe region | 4-22-2 | G3 | hp5 | ||
| 4-22-3 | G1 | hp6 | |||
| 4-22-4 | G1 | hp5 | |||
| 4-22-5 | G3 | hp7 | |||
| 4-22-6 | G3 | hp7 | |||
| 4-22-7 | G3 | hp8 | |||
| 4-22-8 | G1 | hp9 | |||
| 4-22-9 | G1 | hp5 | |||
| Almaty | 5-22-4 | G1 | hp10 | ||
| 5-22-5 | G1 | hp10 | |||
| 5-22-6 | G1 | hp11 | |||
| 5-22-7 | G1 | hp12 | |||
| 5-22-8 | G1 | hp13 | |||
| Atyrau | 6-22-1 | G1 | hp5 | ||
| 6-22-2 | G1 | hp5 | |||
| 6-22-3 | G1 | hp5 | |||
| 6-22-4 | G1 | hp5 | |||
| 6-22-5 | G1 | hp14 | |||
| West-Kazakhstan | 7-22-1 | G1 | hp5 | ||
| 7-22-2 | G1 | hp15 | |||
| 7-22-3 | G1 | hp16 | |||
| 7-22-4 | G1 | hp15 | |||
| 7-22-5 | G1 | hp15 | |||
| Dzhambul | 8-22-9 | G1 | hp5 | ||
| 8-22-10 | G1 | hp15 | |||
| 8-22-11 | G1 | hp5 | |||
| 8-22-12 | G1 | hp17 | |||
| 8-22-13 | G1 | hp17 | |||
| Karagandy | 9-22-11 | G1 | hp18 | ||
| 9-22-12 | G1 | hp5 | |||
| 9-22-13 | G1 | hp19 | |||
| 9-22-14 | G1 | hp5 | |||
| 9-22-15 | G1 | hp20 | |||
| Kostanay | 10-22-1 | G1 | hp5 | ||
| 10-22-2 | G1 | hp21 | |||
| 10-22-3 | G1 | hp22 | |||
| 10-22-4 | G1 | hp23 | |||
| 10-22-5 | G1 | hp23 | |||
| Kyzylorda | 11-22-5 | G1 | hp24 | ||
| 11-22-6 | G1 | hp25 | |||
| 11-22-7 | G1 | hp26 | |||
| 11-22-8 | G1 | hp27 | |||
| 11-22-9 | G1 | hp29 | |||
| Mangystau | 12-22-16 | G1 | hp15 | ||
| 12-22-19 | G1 | hp5 | |||
| 12-22-24 | G1 | hp28 | |||
| 12-22-25 | G1 | hp5 | |||
| 12-22-26 | G1 | hp5 | |||
| South-Kazakhstan region | 13-22-1 | G1 | hp5 | ||
| 13-22-2 | G1 | hp5 | |||
| 13-22-3 | G1 | hp5 | |||
| 13-22-4 | G1 | hp29 | |||
| 13-22-5 | G1 | hp30 | |||
| Pavlodar | 14-22-5 | G1 | hp4 | ||
| 14-22-9 | G1 | hp5 | |||
| 14-22-11 | G1 | hp5 | |||
| 14-22-12 | G1 | hp31 | |||
| 14-22-13 | G1 | hp32 | |||
| North-Kazakhstan region | 15-22-14 | G1 | hp33 | ||
| 15-22-15 | G1 | hp33 | |||
| 15-22-16 | G1 | hp5 | |||
| 15-22-17 | G1 | hp34 | |||
| 15-22-18 | G1 | hp35 | |||
| East-Kazakhstan region | 16-22-16 | G1 | hp36 | ||
| 16-22-17 | G1 | hp12 | |||
| 16-22-18 | G1 | hp37 | |||
| 16-22-19 | G1 | hp38 | |||
| 16-22-21 | G1 | hp5 |
| Nucleotide position | 709 | 710 | 735 | 736* | 737 | 738 | 741 | 742 | 743 | 750 | 757 | 852 | 853 | 854 | 856 | 875 | 876 | 887 | 944 | 975 | 986 | 999 | 1002 | 1047 | 1049 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Hp 1 | A | A | C | C | A | A | T | C | G | A | T | C | G | A | G | G | T | A | C | T | C | T | T | C | C |
| Hp 2 | . | . | . | . | . | . | . | . | . | . | . | A | . | . | . | . | . | . | . | C | . | . | . | . | |
| Hp 3 | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | T | . | T | . | A | A | |
| Hp 4 | C | . | . | . | . | . | . | . | . | . | . | A | . | . | . | . | . | C | T | . | T | . | A | A | |
| Hp 5 | . | . | . | . | . | . | . | . | . | . | . | A | . | . | . | . | . | C | T | . | T | . | A | A | |
| Hp 6 | . | G | . | T | . | T | . | T | . | . | . | A | . | . | . | . | . | C | T | . | T | . | A | A | |
| Hp 7 | . | G | . | T | . | T | . | T | . | . | . | A | . | . | . | T | C | C | T | . | T | . | A | A | |
| Hp 8 | . | G | . | T | T | . | . | T | . | . | . | A | . | . | . | . | C | C | T | . | T | . | A | A | |
| Hp 9 | . | . | . | . | . | . | . | . | . | . | . | A | . | . | . | T | . | C | T | . | T | . | A | A | |
| Hp 10 | . | . | . | . | . | . | . | . | . | . | C | A | . | . | . | . | . | C | T | . | T | . | . | A | |
| Hp 11 | . | . | . | . | . | . | . | . | . | . | . | A | . | . | . | . | . | C | T | . | . | . | A | A | |
| Hp 12 | . | . | . | . | . | . | . | . | . | . | . | A | . | . | . | . | . | C | T | C | . | . | A | A | |
| Hp 13 | . | . | . | . | . | . | . | . | . | . | . | A | . | . | A | . | . | C | T | . | T | . | A | A | |
| Hp 14 | . | . | G | G | . | . | . | . | C | . | . | A | . | . | . | T | . | C | T | . | T | . | A | A | |
| Hp 15 | . | . | . | . | . | . | . | . | . | . | . | A | . | . | . | . | . | C | T | . | T | . | A | A | |
| Hp 16 | . | . | . | . | . | . | C | . | . | . | C | A | . | . | . | . | . | C | T | . | T | C | A | A | |
| Hp 17 | . | . | . | . | . | . | . | . | . | . | . | A | . | . | . | . | . | C | T | . | T | . | A | A | |
| Hp 18 | . | . | . | . | . | . | . | . | . | . | . | A | . | . | . | . | . | C | T | . | T | . | A | A | |
| Hp 19 | . | . | . | . | . | . | . | . | . | . | . | A | . | . | . | . | . | C | T | . | T | . | A | A | |
| Hp 20 | . | . | . | . | . | . | . | . | . | . | . | A | T | T | . | . | C | T | . | T | . | A | A | ||
| Hp 21 | . | . | . | . | . | . | C | . | . | . | . | A | T | T | . | . | C | T | . | T | . | A | A | ||
| Hp 22 | . | . | . | . | . | . | C | . | . | . | . | A | . | . | . | . | C | T | . | T | . | A | A | ||
| Hp 23 | . | . | . | . | . | . | C | . | . | . | . | A | . | . | . | . | C | T | . | . | C | A | A | ||
| Hp 24 | . | . | . | . | . | . | . | . | . | . | . | A | . | . | . | . | C | T | C | . | . | A | A | ||
| Hp 25 | . | . | . | . | . | . | . | . | . | . | . | A | . | . | . | . | C | T | . | T | . | A | A | ||
| Hp 26 | . | . | . | . | . | . | . | . | . | . | . | A | . | . | T | . | . | . | . | . | . | . | A | ||
| Hp 27 | . | . | . | . | . | . | . | . | . | . | . | A | . | . | . | . | . | T | . | T | . | A | A | ||
| Hp 28 | . | . | . | . | . | . | . | . | . | . | . | A | . | . | . | . | C | T | . | T | . | A | A | ||
| Hp 29 | . | . | . | . | . | . | . | . | . | . | . | A | . | . | . | . | C | T | . | T | . | A | A | ||
| Hp 30 | . | . | . | . | . | . | C | . | . | C | . | A | . | . | . | . | C | T | . | . | C | A | A | ||
| Hp 31 | . | . | . | . | . | . | . | . | . | . | . | A | . | . | . | . | C | T | . | T | . | A | A | ||
| Hp 32 | . | . | . | . | . | . | . | . | . | . | . | A | T | . | . | . | C | T | . | T | . | A | A | ||
| Hp 33 | . | . | . | . | . | . | . | . | . | . | . | A | . | . | . | . | C | T | C | . | . | C | . | A | |
| Hp 34 | . | . | . | . | . | . | . | . | . | . | . | A | . | . | . | . | . | T | . | . | . | . | . | ||
| Hp 35 | . | . | . | . | . | . | . | . | . | . | . | A | . | . | . | . | C | T | . | . | . | A | . | ||
| Hp 36 | . | . | . | . | . | . | . | . | . | . | . | A | . | . | . | . | C | T | . | . | . | . | . | ||
| Hp 3 | . | . | . | . | . | . | . | . | . | . | . | A | . | . | . | . | C | . | . | T | . | A | A | ||
| Hp 38 | . | . | . | . | . | . | . | . | . | . | . | A | . | . | . | . | C | T | . | T | . | A | . |
| DNA | n | H | hd | Tajima’s D | Fu’s Fs | FLD | p-value | FLF | p-value |
|---|---|---|---|---|---|---|---|---|---|
| E.g., | 76 | 5 | 0.352 | −2.44548 | −0.285 | −6.26037 | p < 0.01 | −5.59824 | p < 0.01 |
| E.g., | 74 | 34 | 0.843 | −2.51838 | −23.352 | −4.86860 | p < 0.02 | −4.46276 | p < 0.02 |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsParasitic infections in humans and animals · Parasite Biology and Host Interactions · Congenital Anomalies and Fetal Surgery
Introduction
Cystic echinococcosis (CE) is a chronic helminthic zoonosis with a worldwide distribution. CE affects a wide variety of livestock species acting as intermediate hosts (IHs), whereas humans represent aberrant hosts of IHs [1]. Echinococcus parasites require carnivores as final hosts and herbivores as IHs to complete their life cycle. The adult stage inhabits the small intestine of a carnivore and produces hundreds of worms that shed thousands of infected eggs in the stool of the host. As soon as the IH ingests the larvae, they move through the bloodstream to the internal organs, where they form fluid-filled cysts. These cysts may develop into thousands of protoscolices that can mature into adult worms if they are ingested by a definitive host. Human beings are accidental hosts that do not perpetuate the life cycle.
Thus, to date, the Echinococcus granulosus complex comprises five species: E. granulosus sensu stricto (s.s.) (clustering genotypes G1, G2, and G3), Echinococcus equinus (G4), Echinococcus ortleppi (G5), Echinococcus canadensis (grouping genotypes G6–G8 and G10), and Echinococcus felidis (G9) [2–5]. E. granulosus s.l. exhibits a high genotypic diversity with 10 genotypes (G1–G10) that have been molecularly distinguished so far, predominantly based on genetic polymorphism of mitochondrial (mt) genes [6, 7]. E. granulosus s.s. (genotypes G1, G3) is the most wide-spread species of the E. granulosus s.l. complex. In general, reported mt haplotypes are gathered by similarity around G1 or G3, whereas G2 seems to be a micro-variant of G1 or often of G3 [8, 9]. To date, data on the molecular typing of genotyping and haplotypes of E. granulosus complex circulating in Kazakhstan have not been published.
This study aimed to molecularly identify species and haplotypes of the E. granulosus complex infecting cattle in Kazakhstan and to investigate their genetic variation respective to mt targets. Kazakhstan is considered highly endemic to hydatid cysts in humans and animals.
Materials and Methods
Ethical approval
Samples of Echinococcus cysts from cattle were collected from the slaughterhouse and delivered to the parasitological laboratory of the Faculty of Veterinary Medicine. This study was approved by the Animal Ethics Committee of the National Center for Biotechnology (protocol No. 1 dated April 01, 2022). All animal experiments were conducted in accordance with the World Medical Association Code of Ethics (Declaration of Helsinki) (http://ec.europa.eu/environment/chemicals/lab_animals/legal_en.htm).
Study period and location
The study was conducted from August 2022 to October 2023, in the laboratory of parasitology of the Seifullin Kazakh Agrotechnical University and the laboratory of biodiversity and genetic resources of the National Center of Biotechnology, located in Astana, Kazakhstan. Cattle samples were collected from 17 regions of Kazakhstan (Kostanay, Akmola, South Kazakhstan, Pavlodar, Karaganda, East Kazakhstan, West Kazakhstan, Aktobe, Atyrau, Ulytau, North-Kazakhstan, Almaty, Kyzylorda, and Mangystau regions).
DNA extraction and polymerase chain reaction (PCR) analysis
The cysts showed different conditions; some were calcified and others purulent. As a result, the most suitable samples were selected for future studies. DNA was successfully extracted from 73 cysts using a GeneJet genomic DNA purification kit (Thermo Fisher, USA, Cat.: K0701) with some modifications. Briefly, Echinococcus cysts extracted from organs were homogenized in Eppendorf tubes using a pestle, lyses buffer, and Proteinase K.
PCR was applied to identify the genetic diversity of Echinococcus spp. using two primer pairs targeting cytochrome c oxidase subunit 1 (cox1: forward 5′-TTTTTTGGGCATCCTGAGGTTTAT-3′ and reverse 5′-TAAAGAAAGAACATAATGAAAATG-3′) and dehydrogenase subunit 1 (nad1: forward 5′-TGGAACTCAGTTTGAGCTTTACTA-3′ and reverse 5′-ATATCAAAGTAACCTGCTA TGCAG-3′) [4, 10]. Reactions were performed in 15 μL 2^×^ GoTaq Hot Start MasterMix, 9 μL nuclease-free water, 1 μL total primers, and 2.5 μL extracted DNA.
Electrophoresis and sequencing
Agarose gels (1%) were prepared in 1× TBE solution containing 8 ng/µL ethidium bromide (Sigma, E1510). Electrophoresis was performed for 50 min at 120 V using 10 μL PCR products with a DirectLoad 100 bp Low ladder ready-to-use (Sigma, D3687-1VL). The PCR-amplified target gene fragment was purified according to the manufacturer’s protocol using a QIAquick PCR Purification Kit, (Qiagen, Germany, Cat.: 28106). Sequencing was performed using the Seq Studio Genetic Analyzer (Thermo Fisher Scientific Applied Biosystems, USA) according to the manufacturer’s instructions. The resulting nucleotide sequences were visually checked using BioEdit version 7.0. BLAST was used to compare the nucleotide sequences of the studied species with those of other sequences in the National Center for Biotechnology Information (NCBI) GenBank database. Nucleotide sequences of the studied species were deposited in the NCBI GenBank database.
Phylogenetic analysis
Nucleotide sequences obtained in the current study were submitted to GenBank and published under accession numbers for cox1 gene and nad1 (Table-1). The obtained sequences were manually edited, and sequence similarity searches compared to the GenBank reference sequences were performed using BLAST (https://blast.ncbi.nlm.nih.gov). Nucleotide sequences for cox1 and nad1 partial genes were aligned with the MUSCLE multiple sequence alignment program. Phylograms were constructed using a concatenated dataset with the MEGA11 software [11] using the maximum likelihood (ML) method. Mesocestoides spp. (MH998121) was used as an out-group.
Haplotype analysis
The haplotype data file was generated using the DnaSP v.6 software. Statistical parsimony networks were used to analyze haplotype genealogy in cox1 and nad1 concatenated datasets using TCS (Statistical Parsimony by A. R. Templeton, K. A. Crandall, and C. F. Sing) implemented in PopART software [12]. We constructed the networks with a 95% probability limit. To distinguish between synonymous and non-synonymous mutations, nucleotide sequence translation was performed using the DnaSP v.6 software (http://www.ub.edu/dnasp/downloadTv6.html).
Results
Characterization of mtDNA haplotypes
Throughout the study, 700 hydatid cyst samples were meticulously collected from 14 distinct regions, with an average of 3.7 cysts per host. Subsequently, DNA extraction was successfully performed on 76 samples, and sequencing was performed on each of the selected samples. All cysts were identified as E. granulosus s.s. with 38 identified haplotypes among our datasets, including two concatenated mt gene markers: cox I and nad I.
The final aligned sequences with positions 1100–1400 were converted in FASTA and NEXUS format for further analysis using MEGAX, DnaSP v.6, PopArt v. 1.7 software. Point mutations in the sequences were identified after the alignment. Table-1 shows the nucleotide variation positions based on reference sequences. To evaluate the relationships among the different E. granulosus s.s. haplotypes, a network of mt haplotypes based on the concatenated sequences was constructed (Table-1).
Phylogenetic analysis
These samples were subjected to bioinformatic analysis, and the resulting sequences were deposited on the NCBI GenBank platform for further reference.
The obtained sequences were manually edited and sequence similarity searches compared to the GenBank reference sequences were performed using BLAST (https://blast.ncbi.nlm.nih.gov/BLAST/). Nucleotide sequences for concatenated cox1 and nad1 partial genes were aligned with the MUSCLE multiple sequence alignment algorithms.
Analysis of the concatenated sequences of the two partial genes revealed that all samples belonging to E. granulosus were grouped together. A tree was rooted with an out-group by Mesocestoides spp. to produce this grouping (Figure-1).
*Maximum Likelihood phylogenic tree from concatenated sequences of partial genes cox1 and nad1 showing relationships between the examined Kazakhstan isolates and GenBank retrieved related sequences of E. granulosus from neighborhood countries. (outgroup marked with red dot, E. granulosus studied in this research belong to the genotype G3 marked with blue sing). E. granulosus=Echinococcus granulosus.
A set of 84 nucleotide sequences was analyzed using the ML method and the Tamura-Nei model [11] to analyze the evolutionary history of a set of the resulting tree diagram, drawn on a scale, displays the relationships between sequences with branch lengths measured in terms of the number of substitutions per site. A tree with the highest log likelihood (11369.25) is presented, and the proportion of sites where at least one unambiguous base is present in at least one sequence for each descendent clade is indicated next to every internal node in the tree. The final dataset contained a total of 2551 positions.
Various point mutations were identified and analyzed after aligning the sequences. Table-2 summarizes the study findings, which highlight the specific positions of nucleotide variations observed in analyzed sequences.
After analyzing the mt cox1 and nad1 gene sequences, 25 polymorphic sites were identified via alignment. According to parsimonious analysis, 76% (19/25) of these polymorphic sites were informative.
Genetic profile of E. granulosus
After performing a thorough haplotype analysis, the essential focal haplotype was identified and used to configure E. granulosus s.s. in a circular orientation. Among the 73 samples tested, 38 unique haplotypes were identified, with the main haplotype (hp 5) accounting for 30.13% (22 out of 73) of the samples (Figures-2 and 3; Table-1).
The haplotype network for the concatenated mt cox1 and nad1 genes of Echinococcus granulosus. The size of the circles is proportional to the frequency of each haplotype. The number of mutations separating haplotypes is indicated by dash marks. Hap=Haplotype, mt=mitochondrial, cox1=Cytochrome c oxidase subunit 1.
Geographical locations of haplotypes analyzed in the study. The colors of circles on the map are indicated in accordance with the haplotype, and the number of identified haplotypes is shown inside the circle [Source: https://www.qgis.org/en/site/about/].
The findings presented in Table-3 of the current report indicate that the nad1 gene exhibits an increased number of haplotype variations and nucleotide differences.
The analysis indicates that the mt nad1 gene undergoes a higher number of mutations than the cox1 gene, as evidenced by the large number of haplotypes observed. In addition, the haplotype diversity of the nad1 gene was significantly higher, with an indicator value of 0.843 compared to cox1 of 0.352. In addition, the analysis highlights a noteworthy observation regarding the significantly high negative value of Fu’s Fs indicator for the nad1 gene (23.352). This observation could be indicative of recent population expansion or natural selection.
The analysis of haplotypes involved the examination of concatenated mt region of cox1 and nad1 marker regions. Figure-2 clearly illustrates these haplotypes, where each haplotype is differentiated by a unique color. The detailed analysis provides a comprehensive overview of the genetic variation within the studied population.
Discussion
E. granulosus sensu stricto (s.s.) is widely recognized as one of the most prevalent genotypes responsible for cystic Echinococcus (CE) worldwide [13]. mt cox1 and NADH dehydrogenase subunit 1 (nad1) genes serve as crucial evolutionary markers to distinguish inter- and intra-specific variants. Asexual reproduction of Echinococcus spp. occurs during the larval stage, which can result in numerous genetic mutations and variations at both the genus and species level [14, 15].
In the present study, we performed thorough analyses to identify the specific genotypes and haplotypes of Echinococcus cysts that were selected from the cattle population. The aim of this study was to gain a comprehensive understanding of the genetic makeup of these cysts, which would enable us to develop more effective strategies for their management and control.
Because only a few studies have addressed the genetic diversity of the parasite in Kazakhstan [16–18], we sequenced partial mt genes of cox 1 (450 bp) and nad 1 (1200 bp) in cattle metacestodes.
Figure-2 provides a clear visual representation of the widespread distribution of haplotype 5 across the country, with the presence of haplotype 5 being detected in 10 distinct regions. Significantly, haplotype 5 is also an ancestral haplotype, which is further important to its prevalence and distribution. The information presented in Figure-2 highlights the possibility of a specific genetic variant spreading and visually represents the extensive prevalence of echinococcosis across the entire country.
Our research on genotyping revealed two distinct types of genotypes: G1 and G3. The G3 genotype was found exclusively in the Aktobe region. The genotypes G1 and G3 were classified into different haplotypes when constructing the haplotype network. Romig et al. [19] also observed a low level of differentiation between G1 and G3 in the haplotype network. In general, the results of the current study demonstrate low nucleotide diversity but high haplotype diversity. Two genes were negative for Tajima’s D, suggesting population expansion or purifying selection. Recent population expansion or hitchhiking in E. granulosus s.s. can be attributed to the significantly higher negative Fu’s Fs values in mt nad1 (23.352) than in cox1 (0.285) gene sequence comparison, which revealed the presence of rare haplotypes. Fu’s Fs test was created on the basis of haplotype distribution.
CE is prevalent in North Caucasus, Transcaucasia, Kazakhstan, Russia, Kyrgyzstan, Uzbekistan, Moldova, Ukraine, and Turkey, where livestock breeding is common [20–23]. The increase in the incidence of CE can be attributed to changes in agricultural practices following the collapse of the Soviet Union in 1991, which led to the introduction of private slaughterhouses in the backyard and subsequently to the promotion of dog infection. This disease is more common in rural areas where livestock are reared. Several factors contribute to the persistence of CE, including frequent illegal and home slaughtering of animals for food, feeding of raw offal to dogs, low public awareness of the disease, large populations of stray dogs, and poor hygiene conditions [24, 25].
Conclusion
The findings of our research indicate that Echinococcus in Kazakhstan has two distinct genetic variations (G1 and G3). Our analysis showed that the G3 genotype is predominant in a specific region, whereas the G1 genotype is prevalent throughout the country. As the first study of its kind on the territory of Kazakhstan, our research is highly significant in understanding the genetic characteristics of Echinococcus and its potential implications for public health. In addition, our study highlights the need to continue monitoring the spread of echinococcosis throughout the country.
Authors’ Contributions
VK: Contributed to the conception and designed the study and drafted, revised, and finalized the manuscript for submission. RU and AS: Performed DNA extraction and PCR, interpreted the data, and performed phylogenetic analysis, and drafted the manuscript for submission. SY, AA, and KJ: Collected samples and performed parasitological isolation and typing. LL and AT: Analyzed the data and performed the statistical analyses. All authors have read, reviewed, and approved the final manuscript.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Deplazes P Rinaldi L Alvarez Rojas C.A Torgerson P.R Harandi M.F Romig T Antolova D Schurer J.M Lahmar S Cringoli G Magambo J Thompson R.C Jenkins E.J Global distribution of alveolar and cystic echinococcosis Adv. Parasitol 2017953154932813136510.1016/bs.apar.2016.11.001 · doi ↗ · pubmed ↗
- 2Bowles J Blair D Mc Manus D.P Genetic variants within the genus Echinococcus identified by mitochondrial DNA sequencing Mol. Biochem. Parasitol 1992542165173143585710.1016/0166-6851(92)90109-w · doi ↗ · pubmed ↗
- 3Knapp J Nakao M Yanagida T Okamoto M Saarma U Lavikainen A Ito A Phylogenetic relationships within Echinococcus and Taenia tapeworms (Cestoda:Taeniidae):An inference from nuclear protein-coding genes Mol. Phylogenet. Evol 20116136286382190729510.1016/j.ympev.2011.07.022 · doi ↗ · pubmed ↗
- 4Hüttner M Nakao M Wassermann T Siefert L Boomker J.D Dinkel A Sako Y Mackenstedt U Romig T Ito A Genetic characterization and phylogenetic position of Echinococcus felidis (Cestoda:Taeniidae) from the African lion Int. J. Parasitol 20083878618681806297310.1016/j.ijpara.2007.10.013 · doi ↗ · pubmed ↗
- 5Saarma UJõgisalu I Moks E Varcasia A Lavikainen A Oksanen A Simsek S Andresiuk V Denegri G González L.M Ferrer EGárate T Rinaldi L Maravilla PA novel phylogeny for the genus Echinococcus, based on nuclear data, challenges relationships based on mitochondrial evidence Parasitology 200913633173281915465410.1017/S 0031182008005453 · doi ↗ · pubmed ↗
- 6Scott J.C Stefaniak J Pawlowski Z.S Mc Manus D.P Molecular genetic analysis of human cystic hydatid cases from Poland:identification of a new genotypic group (G 9) of Echinococcus granulosus Parasitology 1997114 Pt 13743901107210.1017/s 0031182096008062 · doi ↗ · pubmed ↗
- 7Lavikainen A Lehtinen M.J Meri T Hirvelä-Koski V Meri S Molecular genetic characterization of the Fennoscandian cervid strain, a new genotypic group (G 10) of Echinococcus granulosus Parasitology 2003127 Pt 32072151296482310.1017/s 0031182003003780 · doi ↗ · pubmed ↗
- 8Snábel V Altintas ND'Amelio S Nakao M Romig T Yolasigmaz A Gunes K Turk M Busi MHüttner M SevcováD Ito A Altintas N DubinskýP Cystic echinococcosis in Turkey:Genetic variability and first record of the pig strain (G 7) in the country Parasitol. Res 200910511451541923487610.1007/s 00436-009-1376-2 · doi ↗ · pubmed ↗