Multitemporal single‐cell profiling decoding crosstalk between γδ17 T cells and neutrophils in radiation pneumonitis
Wenting Ren, Xiaoxiang Zhou, Ziming Jiang, Shiqi Li, Haoxuan Zhang, Jianrong Dai, Yexiong Li, Nan Bi, Yibo Gao, Jie He

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
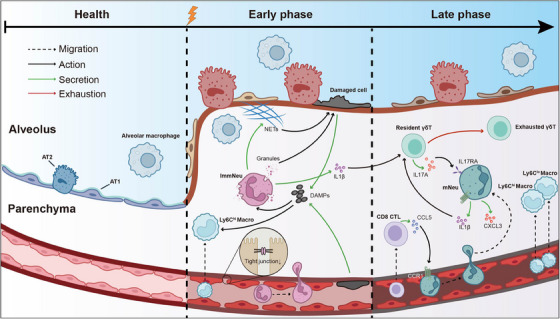 Figure 1
Figure 1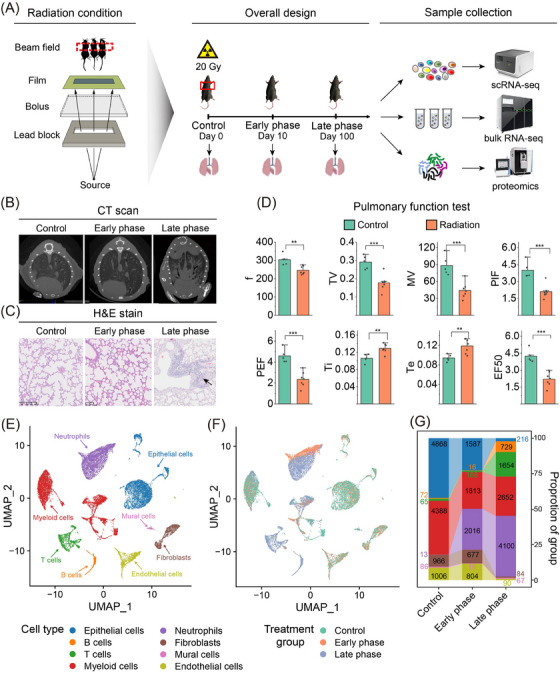 Figure 2
Figure 2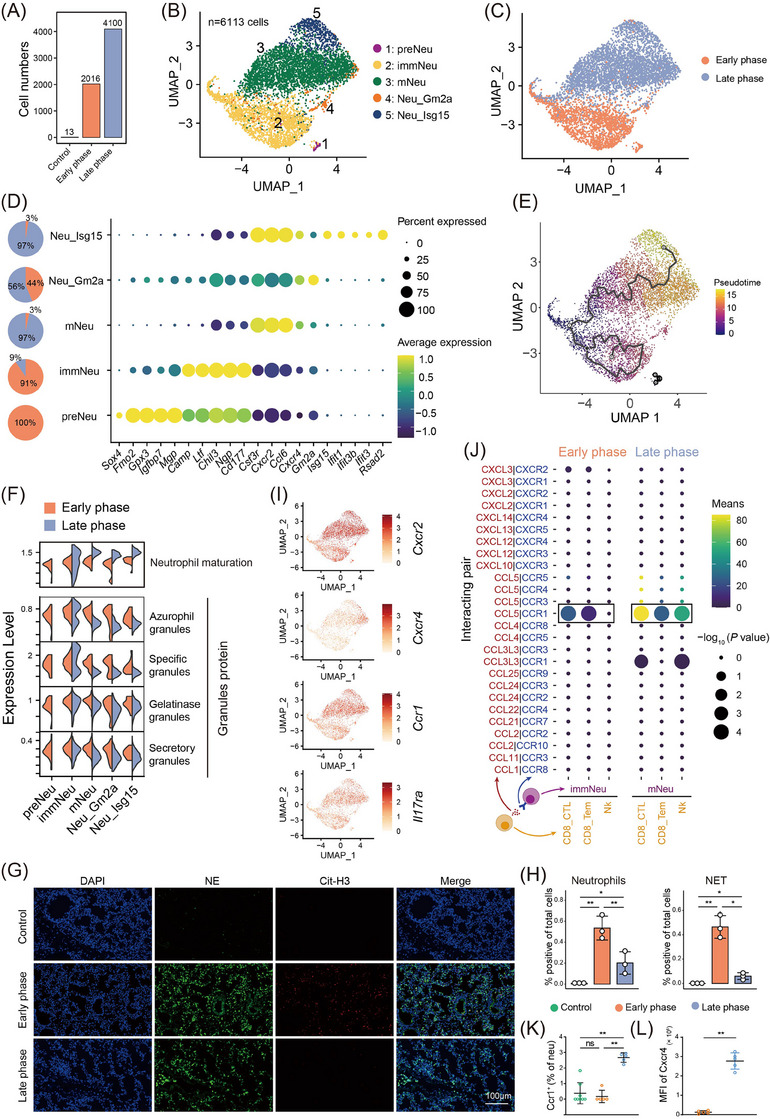 Figure 3
Figure 3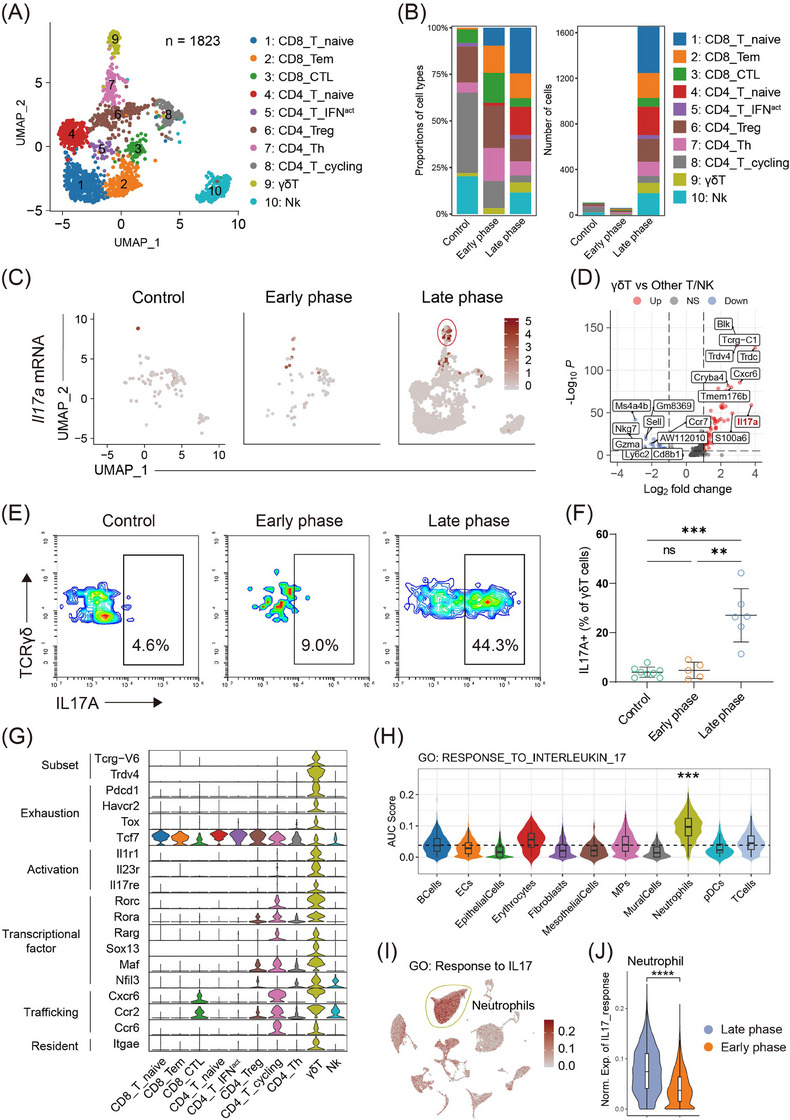 Figure 4
Figure 4- —National Key R&D Program of China 10.13039/501100012166
- —National Natural Science Foundation of China 10.13039/501100001809
- —R&D Program of Beijing Municipal Education Commission
- —CAMS Initiative for Innovative Medicine
- —Key‐Area Research and Development Program of Guangdong Province
- —Shenzhen Science and Technology Program
- —Shenzhen High‐level Hospital Construction Fund, Sanming Project of Medicine in Shenzhen
- —Shenzhen Clinical Research Center for Cancer
- —Aiyou Foundation
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsEffects of Radiation Exposure · Immune Response and Inflammation · Cancer Immunotherapy and Biomarkers
Dear Editor,
Radiation pneumonitis (RP), a form of radiation therapy‐induced normal tissue toxicity, significantly hinders cancer treatment outcomes.1 The involvement of immune cells in RP is not completely understood. Using single‐cell RNA sequencing (scRNA‐seq) combined with bulk RNA sequencing (bulk RNA‐seq) and proteomics, we identified the accumulation of neutrophils and T cells and investigated their function in RP progression. These findings reveal intricate and evolving immunologic alterations in RP, offering insights for potential therapeutic strategies.
In the RP murine model, Mice were exposed to 20 Gy thoracic irradiation, with Day 10 representing the early and Day 100 the late phase of RP (Figure 1A). Classic RP symptoms were observed,2 including weight loss (Figure S1B), hair colour change (Figure S1C), computed tomography scan anomalies (Figure 1B), defective pneumocytes, widening of the alveolar septum, immunocyte infiltration in hematoxylin and eosin (H&E)‐stained slices (Figure 1C), and reduced lung function (Figure 1D). We pooled three mice per group and collected a total of 28,287 cells on days 0, 10, and 100 (Figure 1A). Dynamic cellular changes indicate an increase in the proportions of neutrophils (Csf3r) and T/B lymphocytes (Cd3d/Cd79a) (Figure 1E–G and Figure S1D,E), as verified by H&E staining (Figure 1C). Bulk RNA‐seq and proteomics confirmed elevated S100a8, Ngp, and Retnlg expression in neutrophils (Figure S1E–I).
Post‐radiation, neutrophil infiltration accumulated in pneumonitis and was confirmed by flow cytometry (Figure 2A and Figure S2A,B). We categorized neutrophils into five subsets [pre‐neutrophils (preNeu), immature neutrophils (immNeu), mature neutrophils (mNeu), Isg15^+^ neutrophils (Neu_Isg15), Gm2a^+^ neutrophils (Neu_Gm2a)] (Figure 2B), with preNeu and immNeu prevalent in the early phase, and mNeu, Neu_Isg15, and Neu_Gm2a in the late phase (Figure 2C). Gene expression analysis revealed primary granule genes (Mgp and Camp) highly expressed in preNeu and immNeu, while immune defence genes (Ccl6) and maturation genes (Cxcr2 and Cxcr4) were prominent in mNeu, Neu_Isg15 and Neu_Gm2a (Figure 2D). Additionally, interferon‐stimulated genes (Ifit1 and Isg15) were notably expressed in Neu_Isg15.3
To investigate neutrophil roles in RP, we conducted transcription factor network analysis which revealed preNeu and immNeu enriched with stemness genes (Gata2 and Tbx4), while mNeu and Neu_Isg15 expressed maturation‐related transcription factors (Junb and Cebpd) (Figure S2C). Differential trajectory analysis indicated distinct differentiation states between early and late‐phase neutrophils with no sequential development (Figure 2E). Examining the granule protein gene signature by Xie et al.,3 we found divergent granule protein expression in neutrophil phases (Figure 2F). Surprisingly, early‐phase neutrophils showed increased expression of all granule types. Considering the cytotoxic effects of granule contents in neutrophils,4, 5 we analyzed gene profiles related to cytotoxicity. The immNeu subset showed increased neutrophil extracellular trap (NET) and reactive oxygen species production, causing tissue damage (Figure S2D,E), confirmed by NET enrichment in the early phase (Figure 2G,H). Conversely, mNeu displayed enhanced chemotaxis and transcriptional activity, suggesting stronger cellular interactions (Figure S2D,E). This subset also showed elevated proinflammatory cytokines (Cxcl3 and Il1b), implying persistent inflammation (Figure S2F). Further analysis revealed upregulated chemokine receptors (Il17ra, Ccr1, Cxcr2 and Cxcr4) in late‐phase neutrophils (Figure 2I), with potential CD8^+^ T cell recruitment (Figure 2J,K and Figure S2G). Late‐phase RP also saw increased maturation markers (Cxcr4 and Cxcr2), confirmed by flow cytometry (Figure 2L and Figure S2H).
To decipher the roles of T/NK cells in RP development, we classified T/NK cells into ten subsets (Figure 3A). The identity of each subset was validated by T/NK cell lineage and function marker genes (Figure S3A). All T/NK subsets were enriched in the late phase of RP (Figure 3B). Notably, a subset of γδT cells, marked by high Il17a expression and known as γδ17 T cells, was almost exclusive to the late phase (Figure 3C,D).6 Flow cytometry confirmed an increased proportion of interleukin (IL)17A^+^ γδT cells in late‐phase (Figure 3E,F and Figure S3B). Uniquely, we identified γδ17 T cells by the γδ chain variable region and revealed they belonged to a novel Vδ4^+^ subset and a common Vγ6^+^ subset (Figure 3F), previously noted only in a mouse skin infection model, underscoring the rarity of this γδ17 T cell type.7
We systematically analyzed Vγ6^+^Vδ4^+^ γδ17 T cells in RP, focusing on markers related to exhaustion (Pdcd1, Havcr2), activation (Il1r1, Il23r and Il17re), and so on. These γδ17 T cells showed a terminally exhausted phenotype, evident from Tox gene expression and absence of Tcf7, underlining their critical effector role and progression into a terminally exhausted state. Marked by Itgae, a tissue‐resident memory T cell indicator,8 γδ17 T cells likely expand from tissue‐resident memory cells rather than peripheral recruitment. Furthermore, the elevated expression of activation markers such as Il1r1, Il23r and Il17re, along with previous findings that Il1β (the ligand of Il1r1) is mainly expressed in neutrophils, suggests that γδ17 T cells are likely activated by IL1β released from neutrophils (Figure 3G and Figure S3C,D).
The IL17 response signature, a well‐established Gene Ontology term, was utilized to identify downstream targets of γδ17 T cells. Neutrophils exhibited the highest IL17 response signature activity (Figure 3H,I). Notably, γδ17 T cells were predominantly found in the late stage of RP, suggesting that IL17 response is limited to this phase. We then compared IL17 response signature scores between early and late‐stage neutrophils, finding significant enrichment in the late phase (Figure 3J). These findings indicate that, during late‐stage RP, tissue‐resident γδ17 T cells, activated by IL1β from neutrophils, expand and produce IL17A, thereby enhancing neutrophil recruitment to the lungs. Given that γδ17 T cells and neutrophils conspire to promote breast cancer metastasis,9 targeting γδ17 T/neutrophil axis may enhance efficacy while reducing toxicity. Specifically, we observed an upregulation of Il17re in γδ17 T cells, which is the receptor of Il17c. However, we did not detect any transcriptomic expression of Il17c, making it challenging to trace the origin of Il17c. Given previous reports suggesting that Il17c is upregulated earlier than Il17a in various inflammatory conditions,10 future studies should aim to detect Il17c expression at early time points.
In summary, our study provided a comprehensive and dynamic single‐cell immunological profile of RP progression from the early phase to the late phase (Figure 4). We also detail the roles of other cell types like macrophages, epithelial, and endothelial cells in RP response in Supporting Information (Figure S4‐S6). Briefly, we observed an early‐phase manifestation of increased oxidative stress response in pulmonary alveolar type II cells and compromised endothelial cell integrity, followed by a late‐phase presence of pro‐inflammatory Ly6c^hi^ macrophages in RP. These insights could illuminate RP pathogenesis and suggest new therapeutic avenues.
AUTHOR CONTRIBUTIONS
Conceptualization, methodology and project administration: Wenting Ren and Yibo Gao; Data curation: Wenting Ren; Formal analysis and visualization: Xiaoxiang Zhou, Ziming Jiang and Shiqi Li; Investigation and resources: Haoxuan Zhang; Writing‐original draft: Wenting Ren, Xiaoxiang Zhou, Ziming Jiang and Shiqi Li; Writing‐review & editing: Yibo Gao; Supervision: Jianrong Dai, Yexiong Li, Nan Bi, Yibo Gao and Jie He; Funding acquisition: Yibo Gao and Jie He; All authors read the manuscript, offered feedback and approved it before submission.
CONFLICT OF INTEREST STATEMENT
The authors declare no conflict of interest.
FUNDING INFORMATION
This work was supported by grants from the National Key R&D Program of China (2021YFC2501900), National Natural Science Foundation of China (82122053 and 82188102), R&D Program of Beijing Municipal Education Commission (KJZD20191002302), CAMS Initiative for Innovative Medicine (2021‐1‐I2M‐012), Key‐Area Research and Development Program of Guangdong Province (2021B0101420005), Shenzhen Science and Technology Program (RCJC20221008092811025 and ZDSYS20220606101604009), Shenzhen High‐level Hospital Construction Fund, Sanming Project of Medicine in Shenzhen [grant number SZSM202211011], Shenzhen Clinical Research Center for Cancer [grant number (2021)287], and Aiyou Foundation [grant number KY201701].
ETHICS STATEMENT
All animal experiments conducted in this study were approved by the Ethics Committee of the Cancer Hospital Chinese Academy of Medical Sciences (Approval No. NCC2022A028).
Supporting information
Supporting Information
Supporting Information
Supporting Information
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Palma DA , Senan S , Tsujino K , et al. Predicting radiation pneumonitis after chemoradiation therapy for lung cancer: an international individual patient data meta‐analysis. Int J Radiat Oncol Biol Phys. 2013;85(2):444‐450.22682812 10.1016/j.ijrobp.2012.04.043PMC 3448004 · doi ↗ · pubmed ↗
- 2Wang Z‐D , Qiao Y‐L , Tian X‐F , et al. Toll‐like receptor 5 agonism protects mice from radiation pneumonitis and pulmonary fibrosis. Asia Pac J Cancer Prevent. 2012;13(9):4763‐4767.10.7314/apjcp.2012.13.9.476323167416 · doi ↗ · pubmed ↗
- 3Xie X , Shi Q , Wu P , et al. Single‐cell transcriptome profiling reveals neutrophil heterogeneity in homeostasis and infection. Nat Immunol. 2020;21(9):1119‐1133.32719519 10.1038/s 41590-020-0736-z PMC 7442692 · doi ↗ · pubmed ↗
- 4Cassatella MA , Östberg NK , Tamassia N , Soehnlein O . Biological roles of neutrophil‐derived granule proteins and cytokines. Trends Immunol. 2019;40(7):648‐664.31155315 10.1016/j.it.2019.05.003 · doi ↗ · pubmed ↗
- 5Segel GB , Halterman MW , Lichtman MA . The paradox of the neutrophil's role in tissue injury. J Leukoc Biol. 2011;89(3):359‐372. doi:10.1189/jlb.0910538 21097697 PMC 6608002 · doi ↗ · pubmed ↗
- 6Papotto PH , Reinhardt A , Prinz I , Silva‐Santos B . Innately versatile: γδ17 T cells in inflammatory and autoimmune diseases. J Autoimmun. 2018;87:26‐37.29203226 10.1016/j.jaut.2017.11.006 · doi ↗ · pubmed ↗
- 7Marchitto MC , Dillen CA , Liu H , et al. Clonal Vgamma 6(+)Vdelta 4(+) T cells promote IL‐17‐mediated immunity against Staphylococcus aureus skin infection. Proc Nat Acad Sci USA. 2019;116(22):10917‐10926. doi:10.1073/pnas.1818256116 31088972 PMC 6561199 · doi ↗ · pubmed ↗
- 8Kok L , Masopust D , Schumacher TN . The precursors of CD 8+ tissue resident memory T cells: from lymphoid organs to infected tissues. Nat Rev Immunol. 2022;22(5):283‐293.34480118 10.1038/s 41577-021-00590-3PMC 8415193 · doi ↗ · pubmed ↗