Hepatozoon spp. in stray cats from the metropolitan area of Rio de Janeiro, Brazil
Donato Traversa, Angela Di Cesare, Simone Morelli, Barbara Paoletti, Marika Grillini, Antonio Frangipane di Regalbono, Aline da Silva de Mattos Queiroz, Frederic Beugnet, Leonardo Brustenga, Piermarino Milillo, Luciano Antunes Barros

TL;DR
This study found Hepatozoon felis in 75% of stray cats in Rio de Janeiro, Brazil, and identified four new genetic variations of the parasite.
Contribution
The study reports the first detection of H. felis in Rio de Janeiro and identifies four previously unknown genetic polymorphisms.
Findings
Hepatozoon felis DNA was detected in 21 out of 28 sampled cats.
Four new genetic polymorphisms of H. felis were identified.
This is the first report of H. felis in cats from the state of Rio de Janeiro.
Abstract
In the last few years, the number of studies on feline hepatozoonosis has increased, but our knowledge on the actual species of Hepatozoon and/or different genotypes affecting felines is still incipient. At least three species, namely Hepatozoon felis, H. canis, and H. silvestris, have been isolated from domestic cats in various countries. Additionally, there are indications that other species and genotypes may affect felines in given geographic areas. This study was carried out to investigate the occurrence of Hepatozoon spp. in cats from Niterói, a municipality within the metropolitan area of Rio de Janeiro, Brazil. Individual blood samples were collected from 28 cats enrolled in a spaying/castration program. DNA was extracted from all samples and subjected to sequencing specific for Hepatozoon spp. DNA of H. felis was found in 21/28 cats (75%), and four genetic polymorphisms never…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
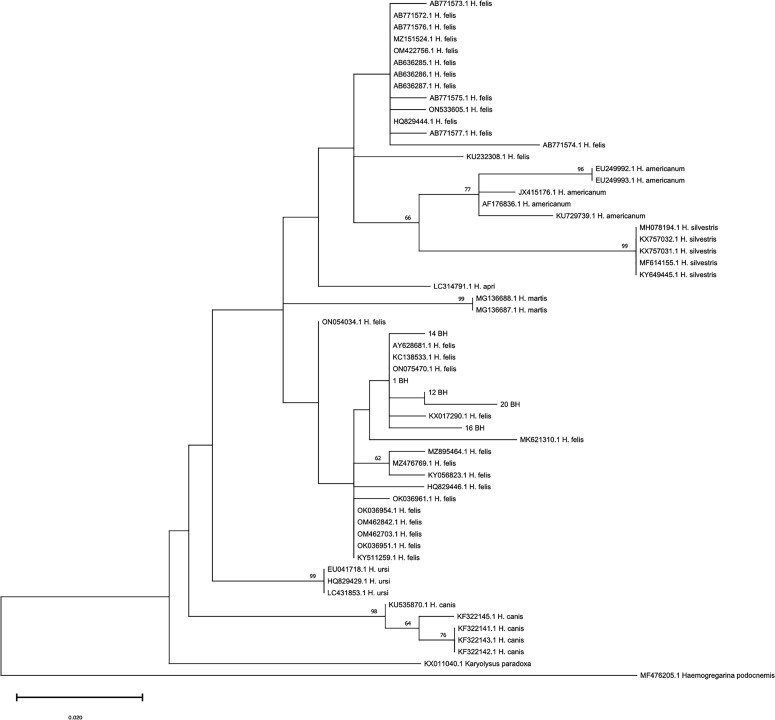 Figure 1
Figure 1 Figure 1
Figure 1- —Boehringer Ingelheim France10.13039/501100014089
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsWittgensteinian philosophy and applications · Literature, Culture, and Aesthetics · Italian Literature and Culture
Introduction
Ticks are vectors of pathogens causing emerging diseases in animals and people worldwide. Most tick-borne diseases (TBDs) are well-known from the epidemiological and clinical standpoints, and this is particularly true for those affecting dogs [21, 42]. Conversely, TBDs of cats are often underestimated and regarded as less important, probably because felines are less susceptible than dogs due to living habits and immunological features [13, 17, 57]. Moreover, many cases of TBDs in cats are often subclinical and the diseases remain undiagnosed and pathogens undetected [11, 12, 42].
However, in recent years, more importance has been attributed to feline TBDs, especially concerning their epidemiology, clinical aspects, and control [7, 14, 47, 49, 52]. Surveys have shown that cats are often positive for pathogens transmitted by ticks in various geographical regions [2, 3, 20, 41, 54, 55] and, among them, Hepatozoon spp. have been repeatedly reported in domestic and wild felids in Europe and elsewhere. Cats can be infected by different species of Hepatozoon, i.e. Hepatozoon felis, Hepatozoon silvestris, and Hepatozoon canis [25, 26, 29, 41], and some species could encompass different genotypes. Several genotypes have been identified in domestic and wild cats worldwide [29, 46], and genotypes of H. felis may have varying tropism for different felines, as suggested for Bengal tigers, Asiatic lions, Indian leopards, and domestic cats in India [46, 47]. Also, it has been hypothesized that some rare genotypes may occur in small/confined areas and that they may have different degrees of pathogenicity [27, 41].
With regard to South America, findings from the last two decades suggest the existence of a high diversity of Hepatozoon genotypes affecting felids [1, 9, 15]. In Brazil, the first records of hepatozoonosis in cats date back almost two decades, when the infection was diagnosed in São Paulo State [49, 53]. The molecular characterization of those isolates has suggested that they are closely related to H. canis [53]. Further studies have then shown that domestic and wild felids from other areas of Brazil may be infected by various species or genotypes of Hepatozoon, including H. canis(*-like), H. felis(-*like), or isolates close to other species [1, 18, 37, 40]. At present, few data are available on the occurrence of Hepatozoon in the State of Rio de Janeiro and they are limited to dogs [23, 43]. No data are available on feline hepatozoonosis in this region of South America, nor on the possible circulation of different species or genotypes of Hepatozoon affecting cats. Additionally, the role of individual single Hepatozoon species/genotypes in causing various diseases in cats remains to be understood. This study aimed to improve the knowledge of this disease in South America by describing the occurrence and genetic variability of Hepatozoon isolates in a population of stray cats.
Materials and methods
Animals, sampling, and ethics
Overall, 28 blood samples were collected from cats included in the sterilization program at the Centro de Castração Prefeitura de Niterói (Rio de Janeiro) – ethics approval No.: CIAEP 0101482014. All samples were stored in ethylenediaminetetraacetic acid (EDTA) tubes and shipped to the Laboratory of Parasitology of the Department of Veterinary Medicine, University of Teramo, Italy, to be molecularly examined for Hepatozoon spp. and Cytauxzoon spp. with the permission of Servicio Público Federal of the Universidade Federal Fluminense (LADDP/FV/UFF N°001/2023) and of the Italian Ministry of Health (0004371-15/02/2023-DGSAF-MDS-P).
Molecular analysis
DNA was extracted from each sample using a commercial kit (Exgene Blood extraction kit, GeneAll Biotech, Songpa-gu Seoul, South Korea), following the manufacturer’s instructions. Specific internal fragments of ~373 bp and of 408 bp of the 18S rRNA gene of Hepatozoon spp. and Cytauxzoon spp. [44], respectively were PCR-amplified as previously described [41, 44, 60], using appropriate positive controls. All amplicons generated (n. 21, see Results) were purified using a QIAquick^®^ Gel Extraction Kit (QIAGEN, GmbH, Hilden, Germany) and sequenced bidirectionally by a commercial laboratory (Macrogen Italy, Milan, Italy). Sequences were determined in both strands, electropherograms were visually checked to rule out the presence of heteroplasmy [50] or of wrong base calls, aligned, and then compared with each other and with those available in GenBank using the Basic Local Alignment Search Tool (BLAST; http://www.ncbi.nlm.nih.gov/BLAST).
An alignment was produced using MEGAX software [32] and along with the obtained sequences one sequence for each of the 30 H. felis haplotypes described by Panda et al. (2024) (Accession numbers: ON533605.1; ON075470.1; ON054034.1; OK036954.1; KY056823.1; KX017290.1; KC138533.1; AB636287.1; AB636286.1; AB636285.1; MZ895464.1; OM422756.1; OM462842.1; OM462703.1; OK036961.1; OK036951.1; MZ476769.1; MZ151524.1; MK621310.1; KY511259.1; KU232308.1; AB771577.1; AB771576.1; AB771575.1; AB771574.1; AB7771573.1; AB771572.1; HQ829446.1; HQ829444.1; AY628681.1), 5 sequences of Hepatozoon americanum (Accession numbers: EU249992.1; EU249993.1; JX415176.1; AF176836.1; KU729739.1), 1 sequence of Hepatozoon apri (Accession number: LC314791.1); 5 sequences of H. canis (Accession numbers: KU535870.1; KF322141.1; KF322142.1; KF322143.1; KF322145.1), 2 sequences of Hepatozoon martis (Accession numbers: MG136687.1; MG136688.1), 5 sequences of H. silvestris (Accession numbers: MH078194.1; KX757031.1; KX757032.1; MF614155.1; KY649445.1); 3 sequences of Hepatozoon ursi (Accession numbers: EU041718.1; HQ829429.1; LC431853.1, and finally one sequence of Karyolysus paradoxa (Accession number: KX011040.1) and Haemogregarina podocnemis (Accession numbers: MF476205.1). The alignment was analyzed in JModelTest [51] to determine the best fitting substitution model to perform a Maximum Likelihood phylogenetic analysis. The evolutionary history was inferred by using the Hasegawa-Kishino-Yano model [28] with discrete Gamma distribution to model evolutionary rate differences among sites. The phylogenetic tree was rooted using K. paradoxa and H. podocnemis as outgroups.
Results
DNA of Hepatozoon felis was found in 21/28 cats (75%), while none of the samples tested positive for Cytauxzoon spp. The sequences were generated from 19 of the 21 Hepatozoon amplicons and revealed the presence of five different genotypes. In particular, 11 isolates (1 BH, GenBank Accession Number PP497034) of this study had 100% identity with H. felis found in Italy in domestic cats (GenBank Accession Number KY649442.1) [25], Spain (GenBank Accession Number AY628681.1) [15], Israel (GenBank Accession Number KC138534) [5], and Uruguay (GenBank Accession Number MT210598 – Bazzano et al., – unpublished). This genotype had 95.80%–97.55% identity with H. felis isolates found in wild felids in Brazil (GenBank Accession Numbers MZ490540, KU232302 and KU232308) [24, 58].
Other sequences herein obtained showed different degrees of identity with the above H. felis isolates from Italy, Spain, Israel, and Uruguay, i.e. 99.74% (n = 2 sequences) (12 BH, GenBank Accession Number PP497035), 99.48% (n = 4 sequences) (14 BH, GenBank Accession Number PP497036), 99.22% (n = 1 sequence) (20 BH, GenBank Accession Number PP497038) and 98.19% (n = 1 sequence) (16 BH, GenBank Accession Number PP497037). Sequence 12 BH, 14 BH, 20 BH, and 16 BH displayed ~95–97% identity with other H. felis isolates from Brazil found in an ocelot (GenBank Accession Number MZ490540) [58], and in jaguars (GenBank Accession Numbers KU232308 and KU232302) [24].
The phylogenetic tree showed that all sequences obtained in the present study grouped within the H. felis clade 1 described by Panda et al., 2024 [46] (Figure 1). Four new and different genetic polymorphisms were found in sequences of the present study, i.e., 12 BH (n = 4 sequences – 1 representative sequence deposited, GenBank Accession Number PP497035), 14 BH (1 sequence, GenBank Accession Number PP497036), 20 BH (1 sequence, GenBank Accession Number PP497038), and 16 BH (1 sequence, GenBank Accession Number PP497037).
Figure 1. Phylogenetic tree showing relationships between isolates obtained in the present study and sequences used in the recent evolutionary analysis on Hepatozoon felis (Panda et al., 2024 [46]). The evolutionary history was inferred by using the Maximum Likelihood method and Hasegawa-Kishino-Yano model. The tree with the highest log likelihood (−783.89) is shown. The percentage of trees in which the associated taxa clustered together is shown above the branches. The tree is drawn to scale, with branch lengths measured in the number of substitutions per site. This analysis involved 58 nucleotide sequences, and there was a total of 184 positions in the final dataset.
Discussion
To the best of the authors’ knowledge, this is the first description of the genetic variability of Hepatozoon spp. affecting domestic cats living in the State of Rio de Janeiro, Brazil.
Epidemiological features of feline hepatozoonosis, including identity of vectors and routes of transmission, have not yet been elucidated.
In analogy to other Hepatozoon species, H. felis is likely transmitted only by ticks, while alternative modes of transmission (i.e., vertical infections) should also be considered [5, 41]. Studies have shown that hepatozoonosis may occur in different environments, e.g., from highly urbanized settings [20], to dry, wetland, or forested areas [2, 41], as in Brazil [9]. This variability is not surprising considering that Rhipicephalus sanguineus sensu lato (s.l.), i.e., the most distributed tick in the world for its capability to live and reproduce in several environments [16, 19, 33, 61], is probably one of the main vectors of H. felis [8, 54]. Accordingly, R. sanguineus s.l. is widespread in Rio de Janeiro State, where it extensively lives in domestic environments [56]. Other ticks that have been suspected to primarily occur in the biology of Hepatozoon in cats of Europe are Ixodes ricinus and Rhipicephalus turanicus [11, 30, 36]. To the best of our knowledge, the presence of I. ricinus and R. turanicus has never been documented in Brazil (https://datadryad.org/stash/dataset/doi:10.5061/dryad.860473k), but other tick species may be involved in the lifecycle of H. felis in this geographic area. Rhipicephalus sanguineus s.l. is the tick species that most commonly parasitizes cats in Brazil, including in the State of Rio de Janeiro [10, 22, 38, 39]. Infestations of Brazilian cats by other tick species, e.g., Amblyomma sculptum and Amblyomma aureolatum [39, 45], have been documented. To date, information on the occurrence of H. felis within different tick species in Brazil is lacking, and studies are needed to investigate whether different species or genotypes of Hepatozoon infecting felids in Brazil, as elsewhere, are associated with R. sanguineus lineages and/or other species of ticks, and what could be the practical implications for their biology, epidemiology and clinical diseases.
The number of samples that were examined molecularly in this study was selected for convenience; therefore, any sound and detailed epidemiological considerations are not possible. Data on infection rates by Hepatozoon in populations of domestic cats in Brazil and in South America are scant and a comparison of the present data in terms of percentage of positivity would be difficult. In any case, it is interesting to note that many cat samples examined in a short timeframe were found to be positive for Hepatozoon, as previously recorded in other surveys [41]. The grooming behavior of cats as they mechanically remove ticks from their body [17, 34] could lead to vector ingestion, thus favoring the transmission of Hepatozoon spp. Moreover, the frequent occurrence of Hepatozoon in populations of felids may also be explained by predation, as some species may be transmitted to vertebrates via ingestion of prey or ticks feeding on prey [4]. Although this has not yet been proven for H. felis or other species/genotypes affecting felids, there is a significant association between hepatozoonosis and outdoor lifestyle in cats, i.e., predation is a likely predisposing factor [5, 6, 35]. Accordingly, all cats infected by Hepatozoon in this study were stray animals living in Niterói municipality.
The taxonomical status of Hepatozoon spp*.* infecting felids is yet to be clarified. It is proposed that H. felis is a species-complex based on a high genetic variability recorded in different studies [27]. Recent findings from Europe, i.e., a new genotype described on a small island of Greece [41], and other phylogenetic analyses have supported this hypothesis [29]. Data from the genetic characterization of H. felis genotypes suggest that some are more widespread than others. For instance, one of the most distributed genotypes in various countries of Europe has also been recorded in the Middle East and South Africa [5, 15, 25, 27, 41]. It is thus interesting to note that the latter genotype showed 100% identity with 1 BH found herein, indicating that this genotype is present in South America and present worldwide. Different Hepatozoon spp. or more than one genotype of H. felis may circulate in cat populations from the same country [27, 29, 41, 46]. The present results are in accordance with this finding, as they showed four different genic polymorphisms never described thus far at the level of the DNA fragment examined, in the small sample of cats studied. Even though the polymorphisms detected here strongly indicate the existence of four new H. felis genotypes, this warrants further genetic investigations. Molecular data available in the literature on Hepatozoon in felids of South America are scant and derive from studies on wild animals, in which different genotypes have recently been found in jaguars and an ocelot [21, 58]. Accordingly, the present analysis, in particular the phylogenetic relationships of the 1 BH isolate and the sequence KU232308 from wild felids (Figure 1), confirms that different genotypes circulate in felids in Brazil, and suggests the occurrence of an H. felis species-complex within the cat population studied here and in felids in South America. On the whole, this scenario from Europe and South America supports the complex-species taxonomic classification recently proposed, and the existence of undescribed separate species [27, 41].
Knowledge on areas endemic for feline hepatozoonosis is of epidemiological relevance for different reasons. Pets travelling with their owners may bring pathogens into free areas or may become infected when visiting endemic regions and bring new pathogens when they return to their region of origin. A constant epidemiological update in these settings is of crucial importance for minimizing the risk that animals may acquire pathogens via arthropods, including hepatozoonosis using appropriate prevention measures [6, 31].
Unfortunately, a detailed clinical history was not available for the cats included in the present study, as they were stray cats in a spaying/castration program. However, a good general health condition was assessed prior to the spaying/castration for all the cats (data not shown). In most cases, hepatozoonosis of cats due to H. felis is subclinical or a mild disease [5], though cases of severe or fatal signs are described [6]. Considering the variability in Hepatozoon infecting felids in various geographical regions, it is plausible that different Hepatozoon spp. or H. felis genotypes may have different tropism and pathogenicity. For instance, cats may also be infected by H. silvestris, a different species with a different tropism within the host (e.g., striated muscle) and a different pathogenic potential, that seem to be higher than H. felis [31, 59]. To date, there are no reports of H. silvestris in South America.
As discussed elsewhere [41], 1 BH found herein has a ~98% identity with a genotype involved in a severe case of hepatozoonosis in a domestic cat in Austria [6]. Nevertheless, clinical implications based on isolates retrieved in this study cannot be discussed here, as a thorough clinical examination with complete blood analyses was not performed for cats in our study.
In conclusion, this study has provided novel information on the occurrence of Hepatozoon spp. infection in Brazil and new comprehensive data on the existence of different genotypes circulating in felids of South America and Europe. Further investigations are needed to ultimately clarify their taxonomical status and whether there is any difference in their relevance and importance in feline clinical practice in terms of epidemiology and pathogenesis.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1André MR, Adania CH, Teixeira RHF, Vargas GH, Falcade M, Sousa L, Salles AR, Allegretti SM, Felippe PAN, Machado RZ. 2010. Molecular detection of Hepatozoon spp. in Brazilian and exotic wild carnivores. Veterinary Parasitology, 173(1–2), 134–138.20630658 10.1016/j.vetpar.2010.06.014 · doi ↗ · pubmed ↗
- 2Attipa C, Papasouliotis K, Solano-Gallego L, Baneth G, Nachum-Biala Y, Sarvani E, Knowles TG, Mengi S, Morris D, Helps C, Tasker S. 2017. Prevalence study and risk factor analysis of selected bacterial, protozoal and viral, including vector-borne, pathogens in cats from Cyprus. Parasites & Vectors, 10, 130.28285597 10.1186/s 13071-017-2063-2PMC 5346881 · doi ↗ · pubmed ↗
- 3Ayllón T, Diniz PPV, Breitschwerdt EB, Villaescusa A, Rodríguez-Franco F, Sainz A. 2012. Vector-borne diseases in client-owned and stray cats from Madrid, Spain. Vector-Borne and Zoonotic Diseases, 12, 143–150.22022820 10.1089/vbz.2011.0729 · doi ↗ · pubmed ↗
- 4Baneth G. 2011. Perspectives on canine and feline hepatozoonosis. Veterinary Parasitology, 181(1), 3–11.21620568 10.1016/j.vetpar.2011.04.015 · doi ↗ · pubmed ↗
- 5Baneth G, Sheiner A, Eyal O, Hahn S, Beaufils JP, Anug Y, Talmi-Frank D. 2013. Redescription of Hepatozoon felis (Apicomplexa: Hepatozoidae) based on phylogenetic analysis, tissue and blood form morphology, and possible transplacental transmission. Parasites & Vectors, 6, 102.23587213 10.1186/1756-3305-6-102PMC 3639113 · doi ↗ · pubmed ↗
- 6Basso W, Görner D, Globokar M, Keidel A, Pantchev N. 2019. First autochthonous case of clinical Hepatozoon felis infection in a domestic cat in Central Europe. Parasitology International, 72, 101945.31228586 10.1016/j.parint.2019.101945 PMC 7108250 · doi ↗ · pubmed ↗
- 7Bergmann M, Hartmann K. 2017. Vector-borne diseases in cats in Germany. Tierärztliche Praxis Ausgabe K: Kleintiere/Heimtiere, 45(5), 329–335.28905982 10.15654/TPK-160874 · doi ↗ · pubmed ↗
- 8Bhusri B, Sariya L, Mongkolphan C, Suksai P, Kaewchot S, Changbunjong T. 2017. Molecular characterization of Hepatozoon felis in Rhipicephalus sanguineus ticks infested on captive lions (Panthera leo). Journal of Parasitic Diseases, 41(3), 903–907.28848300 10.1007/s 12639-017-0902-x PMC 5555946 · doi ↗ · pubmed ↗