Faster Club Hockey Athletes Have Reduced Upper Leg Muscular Co-contraction During Maximal-Speed Sprinting
Jason Williams, Joseph C. Watso

TL;DR
Faster hockey athletes show less muscle co-contraction in their upper legs during sprinting, which may help them run faster.
Contribution
This study provides novel insights into muscle coordination patterns during sprinting in collegiate athletes.
Findings
Faster athletes had lower co-contraction index (CCI) in rectus femoris and biceps femoris.
Reduced CCI was observed during the late swing phase of sprinting.
Findings suggest coordination between these muscles is linked to sprinting velocity.
Abstract
Most electromyographic (EMG) data for muscular activation patterns during ambulation is limited to older adults with existing chronic disease(s) walking at slow velocities. However, we know much less about the lower extremity muscle co-contraction patterns during sprinting and its relation to running velocity (i.e., performance). Therefore, we compared lower extremity muscular activation patterns during sprinting between slower and faster collegiate club hockey athletes. We hypothesized that faster athletes would have lower EMG-assessed co-contraction index (CCI) values in the lower extremities during over-ground sprinting. Twenty-two males (age = 21[1] yrs (median[IQR]); body mass = 77.1 ± 8.6 kg (mean ± SD)) completed two 20-m over-ground sprints with concomitant EMG and asynchronous force plate testing. We split participants using median running velocity (FAST: 8.5 ± 0.3 vs. SLOW:…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
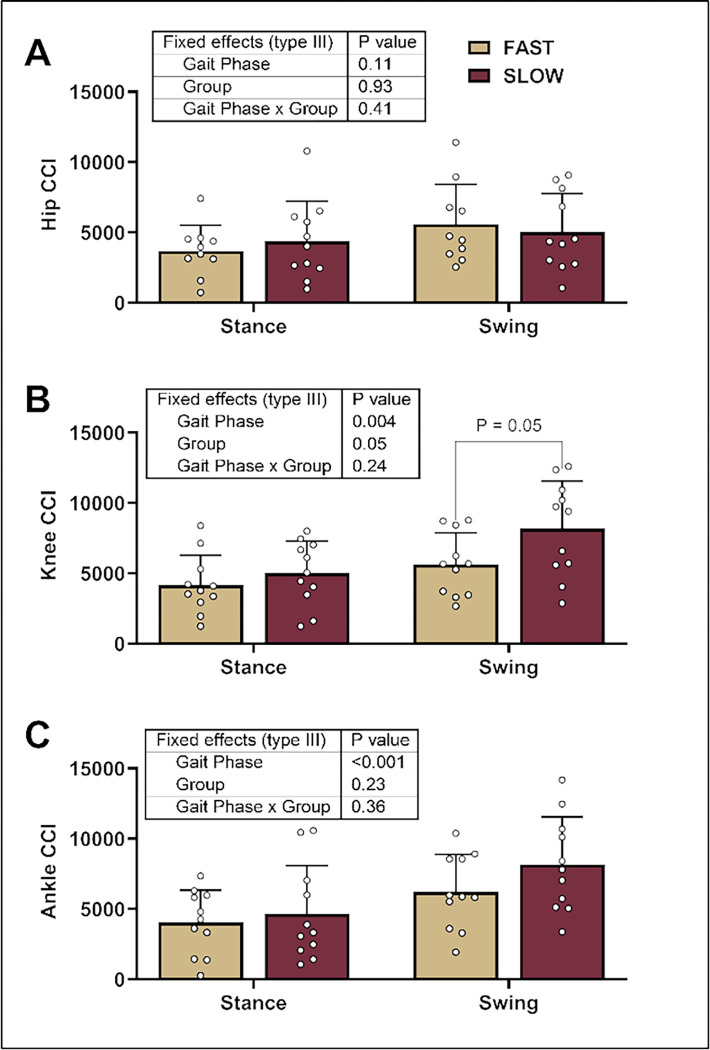 Figure 1
Figure 1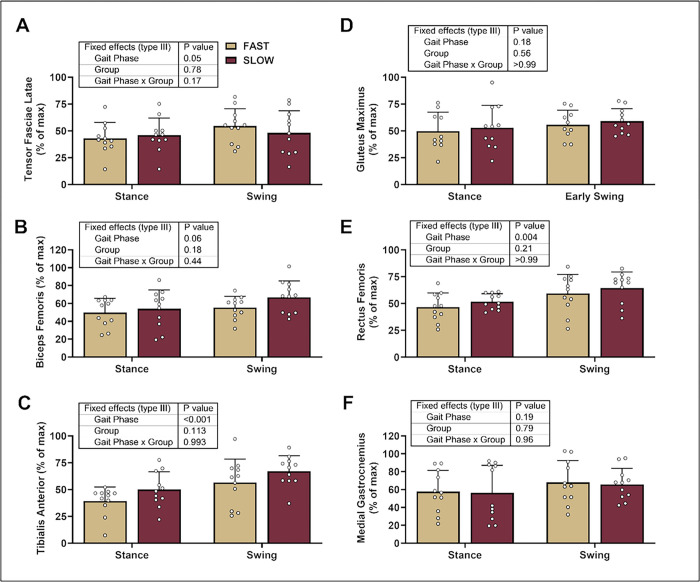 Figure 2
Figure 2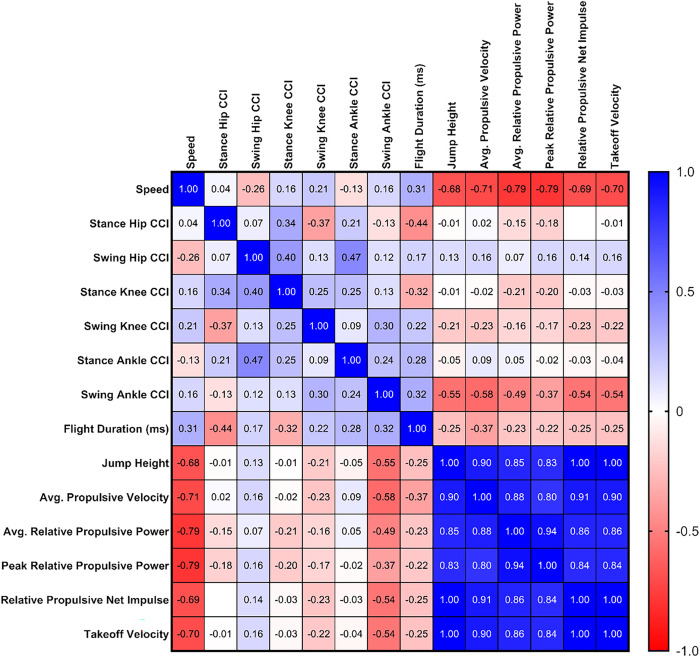 Figure 3
Figure 3- —National Institutes of Health
- —American Heart Association
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsSports injuries and prevention · Muscle activation and electromyography studies · Sports Performance and Training
Background
Muscle activity during maximal speed (upright) sprinting has been thoroughly covered in a systematic review by Howard and colleagues (2018; 1). Studies indicate peak quadriceps activity during the swing or ‘float’ phase (2;3). The quadriceps are also highly active on the ground during the stance phase (4). Similarly, the hamstrings are involved in the swing and stance phases (4; 5), with peak activity during the late swing phase (6; 3). Peak gluteus maximus (GM) activation occurs in the stance phase of sprinting (2;6). Studies also indicate GM activity during the late swing phase (7). Peak EMG activity normalized to maximum voluntary isometric contraction (MVIC) occurred during the foot strike (stance phase) and second float phase (late swing phase) in the biceps femoris (BF), gastrocnemius (GN), and GM, but not in the tibialis anterior (TA) and rectus femoris (RF) (8). Further, EMG values were lowest in the early swing phase for the BF, GN, and GM but highest in the TA and RF. This study is relevant as these athletes sprinted maximally in the upright position (8).
Mero (1992) stated that bi-articular muscles can be antagonistic or synergistic. In the former case, while the antagonist is opposing agonist activity, it provides stiffness and stability to a joint, reducing possible injury. For example, this is critical when an athlete cuts, decelerates, lands, and sprints at high speeds (9). However, co-contraction may hinder performance in certain athletic situations where producing force in minimal time is optimal. For example, increases in RF and BF coactivation increased knee joint stiffness in elite Kenyan runners. However, the energy cost of running increased, likely exerting a negative influence on performance at faster running velocities (10). Similarly, higher co-contraction increases energy cost, and slower running increases co-contraction (11). Also, individuals suffering from musculoskeletal injuries and disorders have higher co-contraction than individuals without musculoskeletal injuries and disorders (12;13). However, scant research exists on performance outcomes of co-contraction related to sprinting ad whether it is associated with sprinting velocity.
Co-contraction of the GM, RF, BF, GN, and TA muscles occurs during the initial contact phase of sprinting (8). During the swing phase, the RF and BF alternate twice as agonist and antagonist muscles (2; 9). The BF works as an agonist at the end of the stance phase by extending the hip while the RF antagonistically co-contracts to maintain posture and provide joint stability (2). Subsequently, the agonist RF flexes the hip and moves the thigh forward in early to mid-swing. This early to mid-swing switch is the ‘first switch’ between the RF and BF. The mid to late-swing switch is the ‘second switch’, where activation transitions from the RF to the BF to extend the hip joint (14). Also, it is essential during the late swing phase that the RF agonistically extends the knee, and the BF eccentrically, isometrically and antagonistically assists with knee extension (16, 5).
The rapid and frequent change of agonist and antagonist relationships between the RF and BF affects step frequency in elite Japanese track and field athletes with an average 100-m time of 10.46 s (15). This study focused on the timing of muscle excitation during the swing and stance phases of maximal sprinting between faster and slower club hockey players. Sprinting requires the rapid and harmonious recruitment of distal and proximal portions of muscles, which consist of activation and relaxation. We sought to investigate the magnitude of this activation via co-contraction. Thus, it is not an unreasonable position that greater co-contraction of muscle indirectly suggests less relaxation and is thus a negative outcome. Therefore, we hypothesized that slower athletes would exhibit greater co-contraction than faster athletes. A secondary purpose was to explore practical differences in muscle excitation between quicker and slower athletes.
Methods
Experimental Approach to the Problem
Subjects.
The Florida State University Institutional Review Board (IRB) approved the study. Subjects were informed of potential risks and benefits and made aware that participation in the experiment was voluntary. All participants provided written consent before testing. We excluded those with an injury in the past six months. The participants completed a short questionnaire to determine their training history, caffeine intake, and activity familiarity. We considered participants who trained more than one day a week for > two years without a detraining interruption of longer than six months as trained (n = 12 trained, n = 10 untrained). We instructed the athletes to avoid alcohol, caffeine, ergogenic aids, and food intake for three hours before each trial.
Day One Testing
Body Mass and Body Composition.
We measured body mass using a force plate (Hawkins Dynamics, ME). Immediately after weighing, we assessed body composition in the seated position using bioelectrical impedance analysis (Omron NBF-306C).
Force Plate Testing.
We assessed force-time metrics via a force plate (Hawkins Dynamics, ME). We evaluated force-time output during a countermovement jump (CMJ), rebound jump, and isometric mid-thigh pull. Participants then engaged in a short dynamic warm-up that consisted of five dynamic stretches of 10 yards each and three submaximal sprints of 70%, 80%, and 90% intensity.
Sprint Testing.
After a three-minute test period following the last submaximal sprint, the athletes completed two maximum 20-meter sprints, measured with electronic timing gates (VALD Performance, Australia). The athletes were given a 15-meter acceleration zone and instructed to reach full speed before hitting the first timing gate. We used the best of two attempts in the analyses.
Day Two Testing
Three days later, the athletes returned individually for EMG analysis. Before sprinting, we placed electrodes, sensors, attached leads, and foot insoles (Noraxon, AZ). The participants executed two 20-yard flying sprints with four minutes between each repetition. The first sprint served as an MVIC, and we normalized the subsequent sprint to the MVIC. This method was chosen instead of a traditional (prone/supine) isometric contraction as Kyröläinen et al. (2005) found that the traditional isometric contraction is not a good indicator of the activation potential since some muscles recorded amplitudes far greater during sprinting (7).
Most often, but not always, the muscles of interest are the GM, RF, BF, GN, tensor fasciae latae (TFL), and TA. Thus, we used EMG to measure the GM, RF, BF, TFL, GN, and TA for each sprint using the MR Noraxon Software v3.4 (Noraxon USA Inc., Scottsdale, AZ). Standard sEMG electrodes were positioned with a 2-cm spacing along the longitudinal axis of the muscles on the right leg, based on the anatomical reference points and following the SENIAM guidelines for sensor placement. We palpated muscles to identify the surface electrode positioning. We shaved the area of each electrode site using a razor and cleaned it with alcohol-soaked cotton wool. Next, we secured the electrodes with tape to reduce motion artifacts (16;17). We set the acceptable impedance (noise) below ten microvolts (uV). Before data treatment, we applied custom-made digital filtering (Bandpass filter; 20–500 Hz), rectification, and smoothing (Root Mean Square algorithm, 100 ms) to the recorded signal. We calculated the reliability of relative muscle activation patterns using intraclass correlation coefficients (ICC; Version 29, SPSS Statistics, IBM, Armonk, NY, USA). The ICCs were: Swing − 0.03 for GM, 0.80 for TFL, 0.87 for BF, 0.60 for RF, 0.67 for GN, & 0.40 for TA; Stance − 0.83 for GM, 0.79 for TFL, 0.89 for BF, 0.07 for RF, 0.83 for GN, & 0.34 for TA.
We inserted Noraxon myoPRESSURE ^™^ foot insoles into the participant’s footwear to determine the swing and stance phases and the application of force by the feet during the stance phase. We instructed the participants to stand on each leg for three seconds to create average pressure plots. We recorded and analyzed the relative EMG activity of the right leg at touchdown and during the early-swing, mid-swing, and late-swing phases. We recorded but did not analyze the peak EMG to an epoch. We did not delineate the stance phase. When we found significance between groups with the overall swing phase, we parsed apart the swing phases. Specifically, the early swing phase begins when the toes leave the ground and ends one-third through the swing phase. The middle swing begins one-third through the swing phase and ends at approximately two-thirds of the total time to complete the swing phase. The late-swing phase starts about two-thirds of the total time to complete the swing phase and ends at ground contact (4; 5).
Muscular co-contraction, or muscular coactivation, is the simultaneous contraction of agonist and antagonist muscles crossing the same joint (18). We assessed muscular co-contraction as follows:
Where = Co-contraction index
Where = level of activity in the less active muscle
Where = level of activity in the more active muscle (19)
Statistical Analysis.
There was no a priori power analysis for this investigation. We split the athletes dichotomously into two groups using a median split of maximum-speed sprinting velocity during the 20 m sprint obtained during testing on day one. We compared groups using unpaired, two-tailed t-tests or Mann-Whitney tests when data failed (p > 0.05) the Shapiro-Wilk test for normality. We also used a Chi-squared test to compare the proportion of trained adults in each group. Using the EMG data from day two, we compared CCI using mixed effect models with Gait Phase (repeated factor) and Group (Independent factor) separately for the ankle (TA & GN), knee (RF & BF), and hip (GM & TFL) to address our primary hypothesis. We also compared gait phase duration using a mixed effect model with Gait Phase (repeated factor) and Group (Independent factor). Next, we compared muscle activation (expressed as absolute uV & as a percentage of their MVC) using mixed effect models with Gait Phase (repeated factor) and Group (Independent factor) for each of the six muscles examined. We employed Sidak’s multiple comparisons test for all mixed effects models for post hoc analyses. Additionally, we used Spearman’s correlational analysis to relate 20 m maximum-speed sprint velocity with several variables of interest. We present values as mean ± standard deviation (SD) for data normally distributed or median [interquartile range] for data not normally distributed. We analyzed data using GraphPad Prism (version 10.0 for Windows, GraphPad Software, San Diego, CA, USA).
Results
We present participant characteristics for the whole cohort and both groups in Table 1. None of these characteristics were significantly different between groups.
We present values as mean ± SD or median [interquartile range] for data that are not normally distributed.
CCI During Over Ground Sprinting.
We found a significant main effect of the gait phase, but not the gait phase * group, for all joints (Fig. 1). We found a lower CCI between the RF and BF (i.e., at the knee joint) in faster sprinters than in slower sprinters (Fig. 1B). With significance during the swing phase, we explored whether the groups differed during the early, mid, or late swing phases. We found a lower CCI between the RF and BF in faster sprinters during the late swing phases (faster: 948 ± 803 vs. slower: 2997 ± 2867; post hoc p = 0.02) but not early or mid-swing phases (post hoc p > 0.81) (gait phase: p < 0.001, group: p = 0.05, gait phase * group: p = 0.14). Finally, we did not observe group differences in the hip or ankle joints (Fig. 1A & 1C).
Muscle Activation During Over Ground Sprinting.
The gait phase significantly affected TFL, RF, and TA activation but not the group or gait phase * group (Fig. 2A, 2C, 2E). While we did not observe group differences in BF activation (Fig. 2B), we explored whether there were groups during the early, mid, or late swing phases for RF and BF activation based on the significant group differences for knee CCI values in the swing phase (Fig. 1B). Interestingly, we found no differences in muscle activation between groups in the RF or BF during the early, mid, or late swing phases (post hoc p ≥ 0.09) (gait phase: p ≤ 0.01, group: p ≥ 0.24, gait phase * group: p ≥ 0.18). Finally, we did not observe group differences in GM or GN activation (Fig. 2D & 2F).
The maximum-speed sprint velocity significantly correlated with all force plate testing outcomes (p < 0.001 for all) but not key CCI values nor flight duration (p > 0.15) (Fig. 3).
Discussion
The primary purpose of this investigation was to determine the muscle activity of three agonist/antagonist muscle pairs and determine if there is more or less muscle co-contraction between faster and slower club hockey players. The novel finding of this study was that faster collegiate club hockey athletes had lower EMG-assessed CCI values in the knee joint but not in the hip or ankle joints during over-ground sprinting. Specifically, the CCI value between the BF and RF CCI was lower in faster sprinters, particularly during the late swing phase. Additionally, we found lower activation of the TA throughout the gait cycle in faster sprinters. These findings suggest that increased relative muscular activation during maximal-speed sprinting may be negatively associated with performance.
During the swing phase, the primary role of the RF is to swing the thigh forward while its antagonist, the BF, extends the hip in preparation and through ground contact (20). During the late swing, the RF extends the tibia while the BF contracts to slow the momentum of the tibia as the knee extends immediately before ground contact (5). The RF activity during the swing phase is considered a hip flexor, not a knee extensor. Thus, abnormally high activation of the leg extensors during the late swing phase is considered atypical (10), which was found to be the case in our study. The RF relative activation percentage was similar between groups during the swing phase. We also found no differences in relative BF activation during the swing phase between groups. However, one should not dismiss the lack of significant BF activation. BF activation was approximately 15% higher in slower sprinters during the late-swing phase and this positively correlated to sprint speed (r = .40). The RF and the BF co-contraction during the swing phase, particularly the late swing phase, is positively coordinated with sprint speed (r = .45). The CCI of the knee during the late swing phase was moderately positively correlated (r = .66) with swing duration, and a longer swing duration results in a lower step frequency. In summary, the CCI values for the knee were significant due to higher activation of the BF in slower sprinters. Moreover, increased co-contraction resulted in slower sprinting, not more, which some researchers have encouraged as a possible positive indicator of faster sprinting.
Intentional anticipatory co-contraction or pretension may reduce muscle slack, resulting in greater stiffness in the series elastic component (tendon) and better performance (21). While studies support the idea that the contractile elements of the hamstring muscle isometrically contract during the swing phase, there is little evidence to support this theoretical framework during sprinting. To this author’s knowledge, no study has demonstrated that the voluntary act of co-contraction during high-speed sprinting would result in better performance. Moreover, inducing more co-contraction would have the opposite effect based on these data. Van Cutsem and Duchateau (2005) indicated that pretension before explosive movements reduced the rate of torque development (22). Thus, intentional muscle co-contraction and its impact on high-speed movement may not be advantageous.
Increased muscle co-contraction may increase muscle and joint stiffness, increasing passive elastic energy return and decreasing muscle slack (22;23). While it is generally agreed that increased stiffness improves performance, it is unclear whether this stiffness results from greater muscle co-contraction or mechanical properties of the tendon. Though only one CCI index was statistically significant, of the CCI calculations, 10 of these values were higher in the slower group than the faster group. Compelling evidence indicates the hamstrings passively lengthen at the beginning of the swing phase and primarily isometrically contract at the end of the swing phase (21). Researchers have investigated the progression of passive lengthening to maximal voluntary isometric contractions (MVIC) in these muscles, albeit at slower speeds but at similar joint angles to sprinting. Nine participants completed a baseline isometric contraction via a dynamometer at 150 degrees. The participants then completed a trial starting at 120 degrees and then were forced into an eccentric contraction to 150 degrees, followed by a maximal isometric contraction (post-stretch isometric contraction). Post-stretch isometric contractions at long muscle lengths resulted in a 20% increase in torque force. In contrast, the muscle activation of the hamstrings considerably decreased compared to maximal isometric performed at 150 degrees (24). Thus, more significant amounts of muscle activation and co-contraction may be compensatory outcomes related to a lack of tendinous and parallel elastic component elasticity and stiffness. In our study, relative RF activation during late swing was nearly identical between groups, whereas BF relative activation was greater (24).
Kakehata et al. (2023) indicated that step frequency depends on RF and BF coordination. The authors referenced this coordination in previous articles as ‘switch,’ which is defined as the relaxation period of the BF and RF muscles in the ipsilateral leg. The authors reported significant negative correlations and large effect size with step frequency and running speed (r = −0.659, −0.655; P < 0.001 for both) for the onset of muscle activity of the BF known as ‘switch 2’. Switch 2 occurred during the late swing phase before foot contact and had a significant positive correlation with step frequency (r = 0.495, P = 0.037). The authors concluded that co-contraction of the BF and RF is disadvantageous for faster sprinting during the swing phase (14). The authors indicated that greater step frequency resulted in harmonious switching between the BF and RF during the late swing phase (14). Importantly, step frequency, not step length, is more closely associated with higher velocity in maximal-speed sprinting (25).
Moore et al (2014) indicated a decrease in co-contraction occurs in the flexor and distal muscles at higher velocities due to the shorter duration of tibialis anterior activation (11). Our study found that faster athletes had lower EMG activity in the TA during sprinting. Relative TFL and RF activation in the mid-swing phase, though not statistically different between slower and faster runners, had a moderate negative correlation to sprint speed. This makes sense as the TFL and RF, both hip flexor muscles, play a crucial role in moving the femur forward and increasing step frequency (25).
Conclusions
These data indicate slower club hockey players experience greater co-contraction, specifically in the late swing phase. It is well known that the rapid activation of agonist muscles reflexively causes the antagonist muscle to relax, a phenomenon known as reciprocal inhibition. It is also understood that training, specifically stretch-shortening cycle activities, can improve the rate and harmonious coordination of agonist and antagonist activity. Thus, stretch-shortening activities designed to emulate the extension of the tibia before ground contact precisely may be advantageous in improving performance. These data provide compelling evidence for human performance practitioners to investigate co-contraction patterns further and work to attenuate them through training interventions.
This research provides evidence of higher values of co-contraction in slower sprinters, specifically between the BF and RF during the late swing phase. We recommend that future research investigates the EMG activity of the RF, BF, and TA during the swing phase in elite populations to see if similar co-contraction patterns exist. If more co-contraction during the late swing phase is differentiated between the fastest sprinters in elite populations, then training interventions to reduce BF activation before touchdown could be needed.
Limitations
This study has clear limitations, including sport specificity, timing of peak muscle activity, limited run through space, an unaccounting for fatigue, and equipment/location constraints. First, the reliability for the stance RF EMG activity was low (ICC = 0.07), but it should be noted the reliability for the stance BF, swing RF, and swing BF were moderate to good (ICC = 0.60–0.89). Second, the cohort was club-level hockey players who may exhibit sport-specific adaptions on the ice that were not captured in the present data collected during overground sprinting. The timing of peak muscle activation is unknown relative to a specific gait cycle phase. Several studies have indicated that RF activates earlier, followed by the BF with increased running speed (6;25). Thus, relative mean values during each swing phase but without precise ‘switch’ moments. The exact moments of muscular activation during each phase are not known. Moreover, the precise moments of peak muscle activation within each phase are unknown, which may be important. As Jonhagen et al. (1996) noted, there is no linear relationship between the relative or magnitude of EMG activity and exerted muscular force (8). A stronger muscle may need less activation, and a weak muscle may require more activation, or the opposite can occur. The reasons for these variations are complex and vast. For example, a more relaxed runner may experience less muscle activation and benefit more from the elastic properties of the musculotendon component. This leads to another limitation: the lack of a musculosketal model to estimate musculotendon length to limit the inevitable crosstalk of multiple muscles crossing multiple joints. Third, the run-up to reach maximal speed was limited to 15 m. Thus, it is possible that maximum velocity was not reached and kinematics more related to acceleration mechanics could have occurred, which would reduce the usefulness of this study to practitioners seeking to employ training interventions. Finally, training volume leading up to testing was uncontrolled. Thus, it is possible some others may have been fatigued, which has been indicated to decrease leg angular velocity, and hip and knee flexion in the swing-phase, which would ultimately affect muscle activation (4).
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Howard R, Conway R, Harrison A. Muscle activity in sprinting: a review. Sports Biomech. 2018;1:1–17.10.1080/14763141.2016.125279028632055 · doi ↗ · pubmed ↗
- 2Higashihara A, Ono T, Kubota J, Okuwaki T, Fukubayashi T. Functional differences in the activity of the hamstring muscles with increasing running speed. J Sports Sci. 2010.https://.10.1080/02640414.2010.49430820672221 · doi ↗ · pubmed ↗
- 3Mann RA, Hagy J. Biomechanics of walking, running, and sprinting. Am J Sports Med. 1980;8:345–50.7416353 10.1177/036354658000800510 · doi ↗ · pubmed ↗
- 4Pinniger G, Steele J, Groeller H. Does fatigue induced by repeated dynamic efforts affect hamstring muscle function? Official J Am Coll Sports Med. 2000;32:647–53.10.1097/00005768-200003000-0001510731008 · doi ↗ · pubmed ↗
- 5Yu B, Queen R, Abbey A, Lui Y, Moorman C, Garrett W. Hamstring muscle kinematics and activation during overground sprinting. J Biomech. 2008;41:3121–6.18848700 10.1016/j.jbiomech.2008.09.005 · doi ↗ · pubmed ↗
- 6Kuitunen S, Komi P, Kyröläinen H. Knee and ankle joint stiffness in sprint running. Med Sci Sports Exerc. 2002;34:166–73.11782663 10.1097/00005768-200201000-00025 · doi ↗ · pubmed ↗
- 7Kyröläinen H, Avela J, Komi P. Changes in muscle activity with increasing running speed. J Sports Sci. 2005;23:1101–9.16194986 10.1080/02640410400021575 · doi ↗ · pubmed ↗
- 8Jon Hagen S, Ericson J, Nemeth G, Erickson E. Amplitude, and timing of electromyographic activity during sprinting. Scand J Med Sci Sports. 1996;6:15–21.8680937 10.1111/j.1600-0838.1996.tb 00064.x · doi ↗ · pubmed ↗