The genome sequence of the Common Emerald, Hemithea aestivaria (Hübner, 1789)
Douglas Boyes, Peter W.H. Holland, Manuela Lopez Villavicencio, Arun Arumugaperumal

TL;DR
This paper provides the genome sequence of the Common Emerald moth, including gene annotations and chromosomal scaffolding.
Contribution
The study presents a high-quality genome assembly and gene annotation for Hemithea aestivaria.
Findings
The genome assembly spans 501.7 megabases and includes 31 chromosomal pseudomolecules.
Gene annotation identified 18,477 protein coding genes using Ensembl.
The mitochondrial genome is 17.05 kilobases in length.
Abstract
We present a genome assembly from an individual male Hemithea aestivaria (the Common Emerald; Arthropoda; Insecta; Lepidoptera; Geometridae). The genome sequence is 501.7 megabases in span. Most of the assembly is scaffolded into 31 chromosomal pseudomolecules, including the Z sex chromosome. The mitochondrial genome has also been assembled and is 17.05 kilobases in length. Gene annotation of this assembly on Ensembl identified 18,477 protein coding genes.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1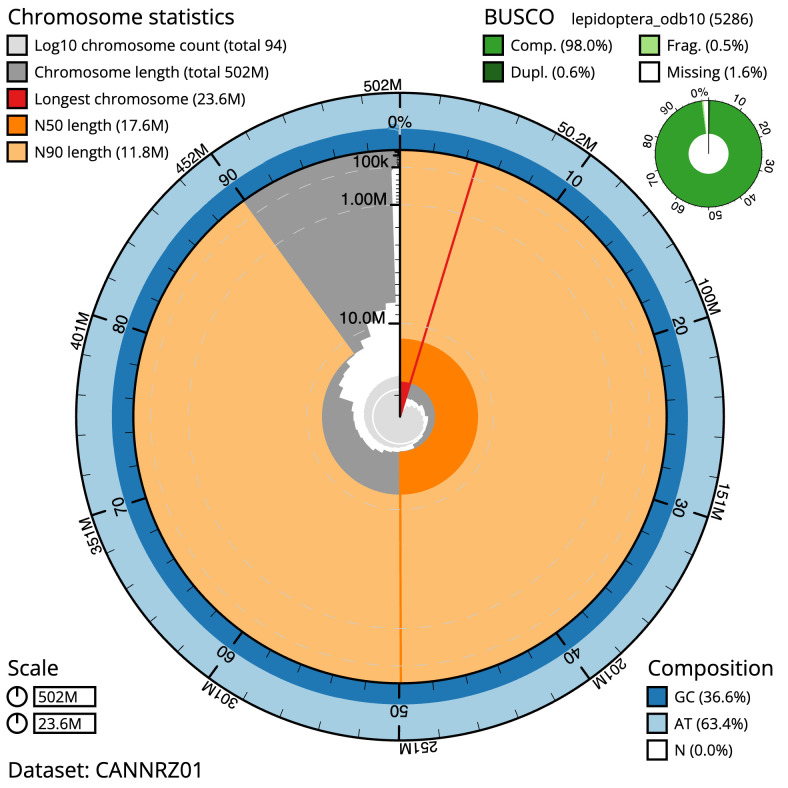 Figure 2
Figure 2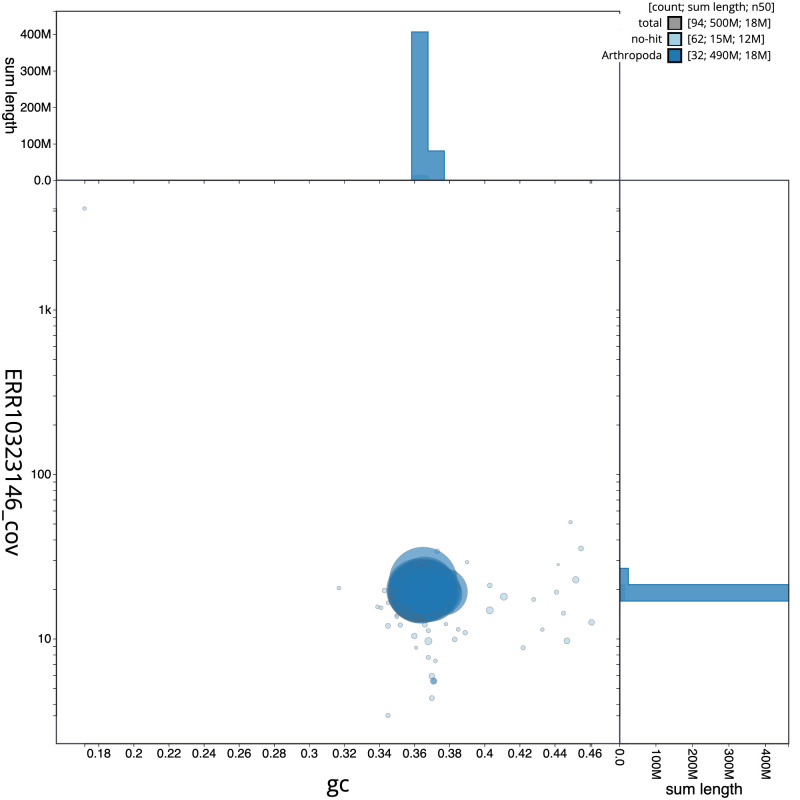 Figure 3
Figure 3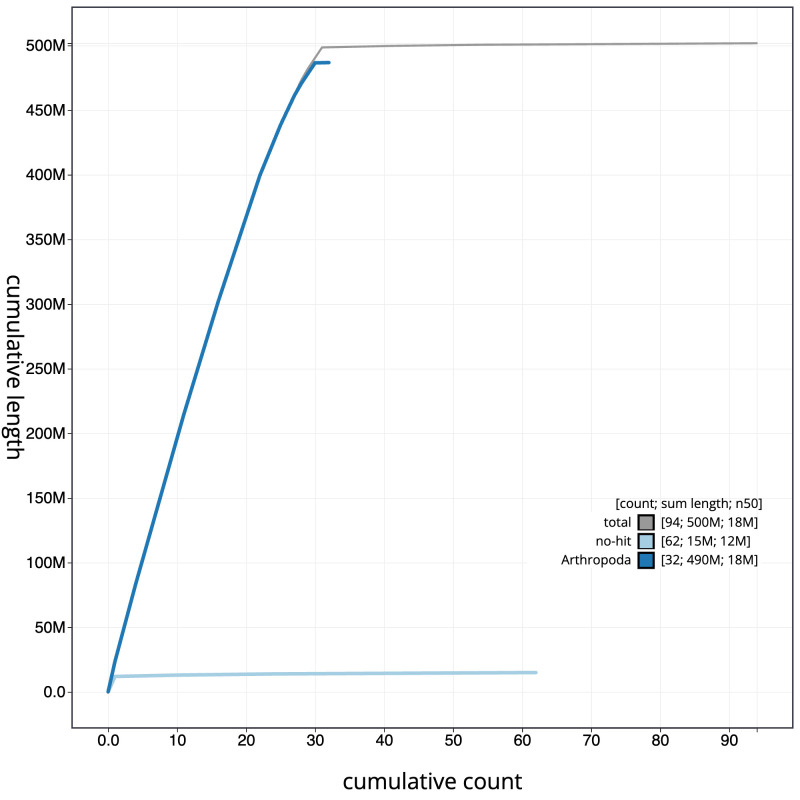 Figure 4
Figure 4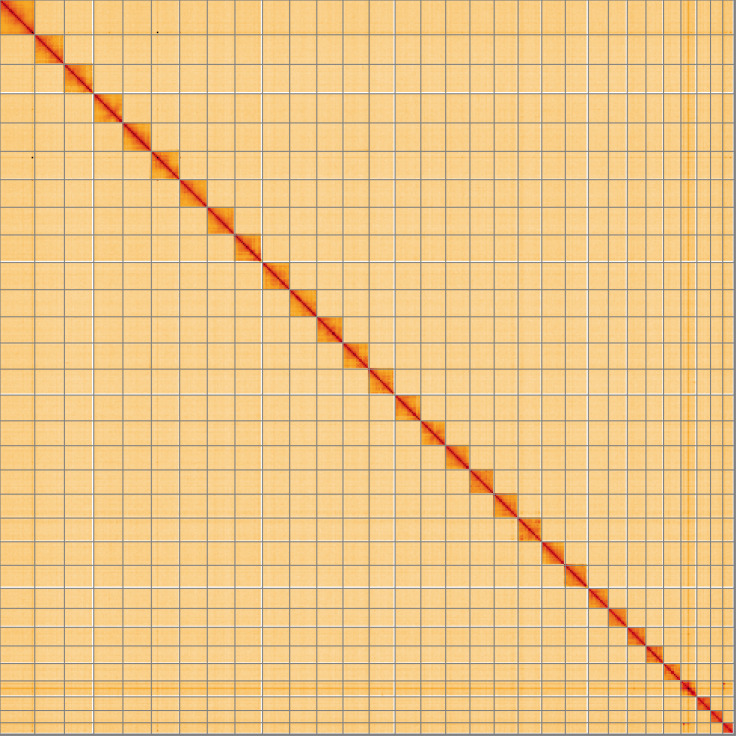 Figure 5
Figure 5| Project accession data | ||
|---|---|---|
| Assembly identifier | ilHemAest2.1 | |
| Species |
| |
| Specimen | ilHemAest2 | |
| NCBI taxonomy ID | 572857 | |
| BioProject | PRJEB56491 | |
| BioSample ID | SAMEA7701444 | |
| Isolate information | ilHemAest2 | |
| Assembly metrics
|
| |
| Consensus quality (QV) | 59.5 |
|
|
| 100% |
|
| BUSCO
| C:98.0%[S:97.4%,D:0.6%],
|
|
| Percentage of assembly mapped
| 99.38% |
|
| Sex chromosomes | Z chromosome |
|
| Organelles | Mitochondrial genome assembled |
|
| Raw data accessions | ||
| PacificBiosciences SEQUEL II | ERR10357397 | |
| Hi-C Illumina | ERR10323148 | |
| Genome assembly | ||
| Assembly accession | GCA_947507615.1 | |
|
| GCA_947461825.1 | |
| Span (Mb) | 501.7 | |
| Number of contigs | 377 | |
| Contig N50 length (Mb) | 2.6 | |
| Number of scaffolds | 93 | |
| Scaffold N50 length (Mb) | 17.6 | |
| Longest scaffold (Mb) | 23.6 | |
| Genome annotation | ||
| Number of protein-coding genes | 18,477 | |
| Number of gene transcripts | 18,682 | |
| INSDC accession | Chromosome | Length (Mb) | GC% |
|---|---|---|---|
| 1 | 20.12 | 36.5 | |
| 2 | 19.94 | 36.0 | |
| 3 | 19.83 | 36.5 | |
| 4 | 19.27 | 36.5 | |
| 5 | 19.24 | 36.5 | |
| 6 | 18.77 | 36.5 | |
| 7 | 18.76 | 36.0 | |
| 8 | 18.71 | 36.5 | |
| 9 | 18.48 | 36.0 | |
| 10 | 18.39 | 36.5 | |
| 11 | 17.79 | 36.5 | |
| 12 | 17.77 | 36.5 | |
| 13 | 17.63 | 36.5 | |
| 14 | 17.45 | 36.5 | |
| 15 | 16.89 | 36.5 | |
| 16 | 16.47 | 36.5 | |
| 17 | 16.46 | 36.5 | |
| 18 | 16.28 | 36.5 | |
| 19 | 16.08 | 36.5 | |
| 20 | 15.97 | 37.0 | |
| 21 | 15.65 | 37.0 | |
| 22 | 13.58 | 36.5 | |
| 23 | 12.88 | 36.5 | |
| 24 | 12.66 | 37.0 | |
| 25 | 11.92 | 36.5 | |
| 26 | 11.81 | 36.5 | |
| 27 | 10.84 | 37.5 | |
| 28 | 9.26 | 37.0 | |
| 29 | 8.3 | 37.0 | |
| 30 | 7.65 | 37.5 | |
| Z | 23.61 | 36.5 | |
| MT | 0.02 | 18.0 |
| Software tool | Version | Source |
|---|---|---|
| BlobToolKit | 4.0.7 |
|
| BUSCO | 5.3.2 |
|
| Hifiasm | 0.16.1-r375 |
|
| HiGlass | 1.11.6 |
|
| Merqury | MerquryFK |
|
| MitoHiFi | 2 |
|
| PretextView | 0.2 |
|
| purge_dups | 1.2.3 |
|
| sanger-tol/genomenote | v1.0 |
|
| sanger-tol/readmapping | 1.1.0 |
|
| YaHS | yahs-1.1.91eebc2 |
|
- —Wellcome Trust
- —Wellcome Trust
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMarine Ecology and Invasive Species · Coral and Marine Ecosystems Studies · Ichthyology and Marine Biology
Species taxonomy
Eukaryota; Metazoa; Eumetazoa; Bilateria; Protostomia; Ecdysozoa; Panarthropoda; Arthropoda; Mandibulata; Pancrustacea; Hexapoda; Insecta; Dicondylia; Pterygota; Neoptera; Endopterygota; Amphiesmenoptera; Lepidoptera; Glossata; Neolepidoptera; Heteroneura; Ditrysia; Obtectomera; Geometroidea; Geometridae; Geometrinae; Hemithea; Hemithea aestivaria (Hübner, 1789) (NCBI:txid572857).
Background
The ‘emeralds’ are a group of over 2000 moth species, most of which have characteristic blue-green wings. Phylogenetic analysis using a small number of genes suggests the group is monophyletic and it is currently classified as a distinct subfamily Geometrinae within the family Geometridae ( Ban et al., 2018; Sihvonen et al., 2011). Of the 10–12 species found in Britain and Ireland, the Common Emerald, Hemithea aestivaria, is one of the most widespread and can be recognised by its dark green angular wings with black and white chequered fringes.
The geographic range of H. aestivaria spans much of Eurasia, from Portugal and Ireland to Japan and Korea ( GBIF Secretariat, 2022). In Britain, the moth is most common in the southern counties of England and has a northern limit in the south of Scotland ( Randle et al., 2019). In Europe, the moth in univoltine with the adult flying in summer; the polyphagous larvae feed on low-growing herbaceous plants in autumn, and after overwintering eat the leaves of woody trees and bushes ( South, 1961). H. aestivaria is recorded as bivoltine in Japan ( Hausmann, 2001). The species is thought to have been introduced accidentally to North America: larvae were first recorded on fruit trees in British Columbia in 1973 before the species spread south to Oregon and Washington State where it is a minor pest of apple orchards ( Doǧanlar & Beirne, 1979; LaGasa, 1996; Looney et al., 2016; Schmidt & Anctil, 2021). Since 2019, H. aestivaria has also been recorded on the east coast of Canada in Ontario, Québec and Nova Scotia ( Schmidt & Anctil, 2021).
The green colour of emerald moths has long intrigued entomologists due to its propensity to fade in living individuals and in dried museum specimens. It also a recognisably different shade to the green on other lepidopteran wings. The colour is conferred by a pigment located in granules within the wing scales; extractions using wings of H. aestivaria and other emerald moths has shown it to be a light-sensitive polar molecule, most likely bound to protein ( Cook, 1993; Cook et al., 1994). The chemical structure of the pigment (named geoverdin) and the biochemical pathway for its production have not been determined.
The genome sequence of Hemithea aestivaria was determined as part of the Darwin Tree of Life project. The assembled genome sequence will facilitate research into the biochemistry underpinning pigment synthesis in insects, and contribute to the growing set of resources for studying lepidopteran ecology and evolution.
Genome sequence report
The genome was sequenced from one male Hemithea aestivaria ( Figure 1) collected from Wytham Woods, Oxfordshire (51.77, –1.34). A total of 36-fold coverage in Pacific Biosciences single-molecule HiFi long reads was generated. Primary assembly contigs were scaffolded with chromosome conformation Hi-C data. Manual assembly curation corrected 67 missing joins or mis-joins and removed 17 haplotypic duplications, reducing the assembly length by 0.64% and the scaffold number by 10.48%, and increasing the scaffold N50 by 0.48%.
Photograph of the Hemithea aestivaria (ilHemAest2) specimen used for genome sequencing.
The final assembly has a total length of 501.7 Mb in 93 sequence scaffolds with a scaffold N50 of 17.6 Mb ( Table 1). Most (99.38%) of the assembly sequence was assigned to 31 chromosomal-level scaffolds, representing 30 autosomes and the Z sex chromosome. Chromosome-scale scaffolds confirmed by the Hi-C data are named in order of size ( Figure 2– Figure 5; Table 2). While not fully phased, the assembly deposited is of one haplotype. Contigs corresponding to the second haplotype have also been deposited. The mitochondrial genome was also assembled and can be found as a contig within the multifasta file of the genome submission.
Table 1.: Genome data for Hemithea aestivaria, ilHemAest2.1.
Genome assembly of Hemithea aestivaria, ilHemAest2.1: metrics. The BlobToolKit Snailplot shows N50 metrics and BUSCO gene completeness. The main plot is divided into 1,000 size-ordered bins around the circumference with each bin representing 0.1% of the 501,713,186 bp assembly. The distribution of scaffold lengths is shown in dark grey with the plot radius scaled to the longest scaffold present in the assembly (23,607,823 bp, shown in red). Orange and pale-orange arcs show the N50 and N90 scaffold lengths (17,632,203 and 11,810,528 bp), respectively. The pale grey spiral shows the cumulative scaffold count on a log scale with white scale lines showing successive orders of magnitude. The blue and pale-blue area around the outside of the plot shows the distribution of GC, AT and N percentages in the same bins as the inner plot. A summary of complete, fragmented, duplicated and missing BUSCO genes in the lepidoptera_odb10 set is shown in the top right. An interactive version of this figure is available at https://blobtoolkit.genomehubs.org/view/ilHemAest2.1/dataset/CANNRZ01/snail.
Genome assembly of Hemithea aestivaria, ilHemAest2.1: BlobToolKit GC-coverage plot. Scaffolds are coloured by phylum. Circles are sized in proportion to scaffold length. Histograms show the distribution of scaffold length sum along each axis. An interactive version of this figure is available at https://blobtoolkit.genomehubs.org/view/ilHemAest2.1/dataset/CANNRZ01/blob.
Genome assembly of Hemithea aestivaria, ilHemAest2.1: BlobToolKit cumulative sequence plot. The grey line shows cumulative length for all scaffolds. Coloured lines show cumulative lengths of scaffolds assigned to each phylum using the buscogenes taxrule. An interactive version of this figure is available at https://blobtoolkit.genomehubs.org/view/ilHemAest2.1/dataset/CANNRZ01/cumulative.
Genome assembly of Hemithea aestivaria, ilHemAest2.1: Hi-C contact map of the ilHemAest2.1 assembly, visualised using HiGlass. Chromosomes are shown in order of size from left to right and top to bottom. An interactive version of this figure may be viewed at https://genome-note-higlass.tol.sanger.ac.uk/l/?d=b1h_oRM_RIu7RhwFE-mCfA.
Table 2.: Chromosomal pseudomolecules in the genome assembly of Hemithea aestivaria, ilHemAest2.
The estimated Quality Value (QV) of the final assembly is 59.5 with k-mer completeness of 100%, and the assembly has a BUSCO v5.3.2 completeness of 98.0% (single = 97.4%, duplicated = 0.6%), using the lepidoptera_odb10 reference set ( n = 5,286).
Metadata for specimens, spectral estimates, sequencing runs, contaminants and pre-curation assembly statistics can be found at https://links.tol.sanger.ac.uk/species/572857.
Genome annotation report
The Hemithea aestivaria genome assembly (GCA_947507615.1) was annotated using the Ensembl rapid annotation pipeline ( Table 1; https://rapid.ensembl.org/Hemithea_aestivaria_GCA_947507615.1/Info/Index). The resulting annotation includes 18,682 transcribed mRNAs from 18,477 protein-coding genes.
Methods
Sample acquisition and nucleic acid extraction
The specimen selected for genome sequencing was a male Hemithea aestivaria (ilHemAest2), collected from Wytham Woods, Oxfordshire (biological vice-county Berkshire), UK (latitude 51.77, longitude –1.34) on 2020-07-05. The specimen was taken from woodland habitat by Douglas Boyes (University of Oxford) using a light trap. The specimen was identified by the collector, and then snap-frozen on dry ice.
DNA was extracted at the Tree of Life laboratory, Wellcome Sanger Institute (WSI). The ilHemAest2 sample was weighed and dissected on dry ice with tissue set aside for Hi-C sequencing. Whole organism tissue was disrupted using a Nippi Powermasher fitted with a BioMasher pestle. High molecular weight (HMW) DNA was extracted using the Qiagen MagAttract HMW DNA extraction kit. HMW DNA was sheared into an average fragment size of 12–20 kb in a Megaruptor 3 system with speed setting 30. Sheared DNA was purified by solid-phase reversible immobilisation using AMPure PB beads with a 1.8X ratio of beads to sample to remove the shorter fragments and concentrate the DNA sample. The concentration of the sheared and purified DNA was assessed using a Nanodrop spectrophotometer and Qubit Fluorometer and Qubit dsDNA High Sensitivity Assay kit. Fragment size distribution was evaluated by running the sample on the FemtoPulse system.
Sequencing
Pacific Biosciences HiFi circular consensus d DNA sequencing libraries were constructed according to the manufacturers’ instructions. Poly(A) RNA-Seq libraries were constructed using the NEB Ultra II RNA Library Prep kit. DNA and RNA sequencing was performed by the Scientific Operations core at the WSI on the Pacific Biosciences SEQUEL II (HiFi) instrument. Hi-C data were also generated from tissue of ilHemAest2 that had been set aside, using the Arima2 kit and sequenced on the Illumina NovaSeq 6000 instrument.
Genome assembly, curation and evaluation
Assembly was carried out with Hifiasm ( Cheng et al., 2021) and haplotypic duplication was identified and removed with purge_dups ( Guan et al., 2020). The assembly was then scaffolded with Hi-C data ( Rao et al., 2014) using YaHS ( Zhou et al., 2023). The assembly was checked for contamination and corrected described previously ( Howe et al., 2021). Manual curation was performed using HiGlass ( Kerpedjiev et al., 2018) and Pretext ( Harry, 2022). The mitochondrial genome was assembled using MitoHiFi ( Uliano-Silva et al., 2022), which runs MitoFinder ( Allio et al., 2020) or MITOS ( Bernt et al., 2013) and uses these annotations to select the final mitochondrial contig and to ensure the general quality of the sequence.
A Hi-C map for the final assembly was produced using bwa-mem2 ( Vasimuddin et al., 2019) in the Cooler file format ( Abdennur & Mirny, 2020). To assess the assembly metrics, the k-mer completeness and QV consensus quality values were calculated in Merqury ( Rhie et al., 2020). This work was done using Nextflow ( Di Tommaso et al., 2017) DSL2 pipelines “sanger-tol/readmapping” ( Surana et al., 2023a) and “sanger-tol/genomenote” ( Surana et al., 2023b). The genome was analysed within the BlobToolKit environment ( Challis et al., 2020) and BUSCO scores ( Manni et al., 2021; Simão et al., 2015) were calculated.
Table 3 contains a list of relevant software tool versions and sources.
Genome annotation
The BRAKER2 pipeline ( Brůna et al., 2021) was used in the default protein mode to generate annotation for the Hemithea aestivaria assembly (GCA_947507615.1) in Ensembl Rapid Release.
Wellcome Sanger Institute – Legal and Governance
The materials that have contributed to this genome note have been supplied by a Darwin Tree of Life Partner. The submission of materials by a Darwin Tree of Life Partner is subject to the ‘Darwin Tree of Life Project Sampling Code of Practice’, which can be found in full on the Darwin Tree of Life website here. By agreeing with and signing up to the Sampling Code of Practice, the Darwin Tree of Life Partner agrees they will meet the legal and ethical requirements and standards set out within this document in respect of all samples acquired for, and supplied to, the Darwin Tree of Life Project.
Further, the Wellcome Sanger Institute employs a process whereby due diligence is carried out proportionate to the nature of the materials themselves, and the circumstances under which they have been/are to be collected and provided for use. The purpose of this is to address and mitigate any potential legal and/or ethical implications of receipt and use of the materials as part of the research project, and to ensure that in doing so we align with best practice wherever possible. The overarching areas of consideration are:
Ethical review of provenance and sourcing of the materialLegality of collection, transfer and use (national and international)
Each transfer of samples is further undertaken according to a Research Collaboration Agreement or Material Transfer Agreement entered into by the Darwin Tree of Life Partner, Genome Research Limited (operating as the Wellcome Sanger Institute), and in some circumstances other Darwin Tree of Life collaborators.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Abdennur N Mirny LA : Cooler: Scalable storage for Hi-C data and other genomically labeled arrays. Bioinformatics. 2020;36(1):311–316. 10.1093/bioinformatics/btz 540 31290943 PMC 8205516 · doi ↗ · pubmed ↗
- 2Allio R Schomaker-Bastos A Romiguier J : Mito Finder: Efficient automated large‐scale extraction of mitogenomic data in target enrichment phylogenomics. Mol Ecol Resour. 2020;20(4):892–905. 10.1111/1755-0998.13160 32243090 PMC 7497042 · doi ↗ · pubmed ↗
- 3Ban X Jiang N Cheng R : Tribal classification and phylogeny of Geometrinae (Lepidoptera: Geometridae) inferred from seven gene regions. Zool J Linn Soc. 2018;184(3):653–672. 10.1093/zoolinnean/zly 013 · doi ↗
- 4Bernt M Donath A Jühling F : MITOS: Improved de novo metazoan mitochondrial genome annotation. Mol Phylogenet Evol. 2013;69(2):313–319. 10.1016/j.ympev.2012.08.023 22982435 · doi ↗ · pubmed ↗
- 5Brůna T Hoff KJ Lomsadze A : BRAKER 2: Automatic eukaryotic genome annotation with Gene Mark-EP+ and AUGUSTUS supported by a protein database. NAR Genom Bioinform. 2021;3(1):lqaa 108. 10.1093/nargab/lqaa 108 33575650 PMC 7787252 · doi ↗ · pubmed ↗
- 6Challis R Richards E Rajan J : Blob Tool Kit – interactive quality assessment of genome assemblies. G 3 (Bethesda). 2020;10(4):1361–1374. 10.1534/g 3.119.400908 32071071 PMC 7144090 · doi ↗ · pubmed ↗
- 7Cheng H Concepcion GT Feng X : Haplotype-resolved de novo assembly using phased assembly graphs with hifiasm. Nat Methods. 2021;18(2):170–175. 10.1038/s 41592-020-01056-5 33526886 PMC 7961889 · doi ↗ · pubmed ↗
- 8Cook M : The systematics of Emerald moths (Geometridae, Geometrinae). University of Oxford. 1993; (Accessed: 23 May 2023). Reference Source