Effect of environmental variables on the abundance of Amblyomma ticks, potential vectors of Rickettsia parkeri in central Brazil
Isadora R. C. Gomes, Rodrigo Gurgel-Gonçalves, Gilberto S. Gazeta, Ana B. P. Borsoi, Karla Bitencourth, Letícia F. Leite, Nathália G. S. S. Coelho, Ricardo Dislich, Helga C. Wiederhecker, Eduardo G. Santos, Melina Guimarães, Shawky M Aboelhadid, Shawky M Aboelhadid

TL;DR
This study examines how environmental factors influence the abundance of Amblyomma ticks, which can carry Rickettsia parkeri, in central Brazil.
Contribution
The study identifies A. dubitatum as a potential vector of R. parkeri and highlights the role of climate over host presence in tick abundance.
Findings
Amblyomma ticks are most abundant during the dry season, especially in larval and nymphal stages.
Climatic factors, not vegetation or capybara presence, best explain tick abundance.
A. dubitatum ticks were found to carry Rickettsia parkeri, suggesting their role as potential vectors.
Abstract
Amblyomma ticks are vectors of both Rickettsia rickettsii and R. parkeri in the Americas, where capybaras (Hydrochoerus hydrochaeris) are the main hosts in urban areas, thus contributing to the transmission of spotted fever. Herein, we studied: (i) the seasonal dynamics and abundance of ticks in areas where capybaras live, (ii) the effect of environmental variables on tick abundance, and (iii) the presence of Rickettsia-infected ticks. Between September 2021 and September 2022, we sampled ticks using cloth-dragging at 194 sites on the shore of Lake Paranoá in Brasília, Brazil. We measured environmental data (season, vegetation type, canopy density, temperature, humidity, and presence or vestige of capybara) at each site. Nymphs and adults were morphologically identified to the species level, and a selected tick sample including larvae was subjected to genotypic identification. We…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
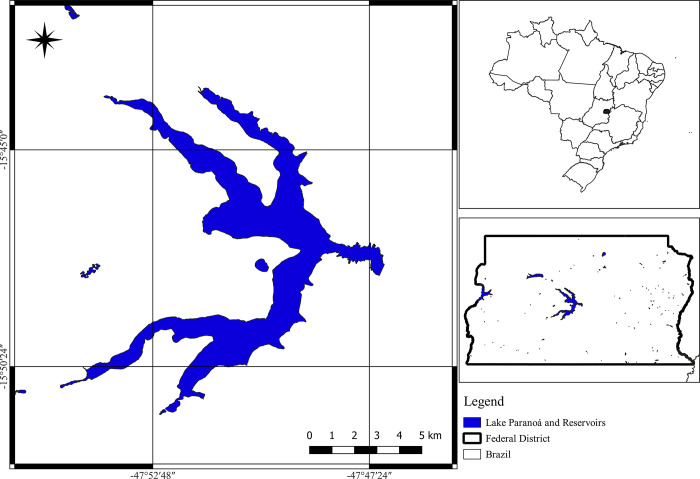 Fig 1
Fig 1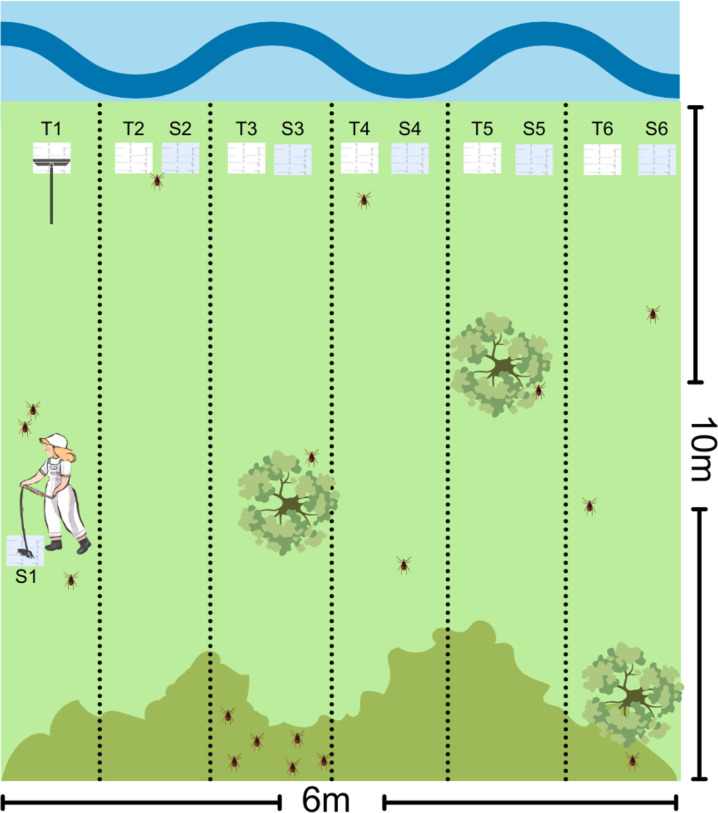 Fig 2
Fig 2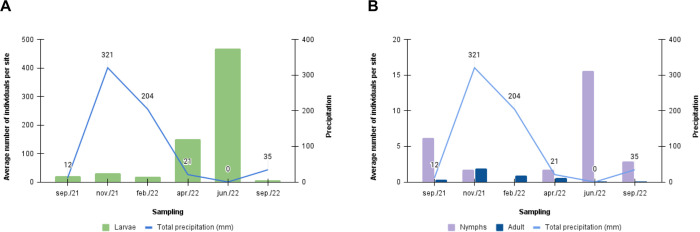 Fig 3
Fig 3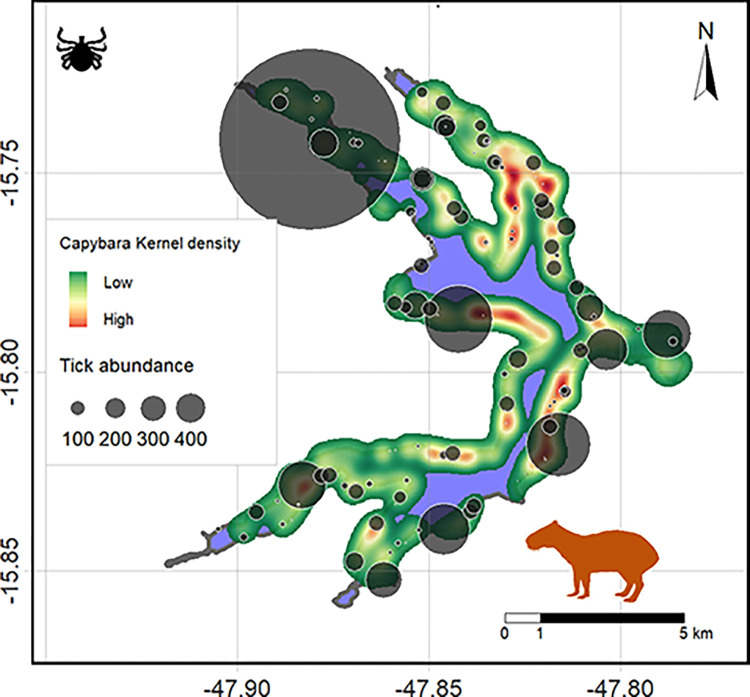 Fig 4
Fig 4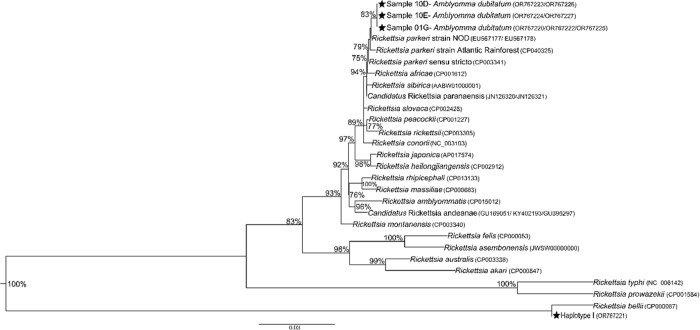 Fig 5
Fig 5- —http://dx.doi.org/10.13039/501100002322Coordenação de Aperfeiçoamento de Pessoal de Nível Superior
- —http://dx.doi.org/10.13039/501100003593Conselho Nacional de Desenvolvimento Científico e Tecnológico
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAdvanced Mathematical Theories and Applications · Advanced Mathematical Identities · Advanced Mathematical Theories
Introduction
Amblyomma ticks (Acari: Ixodidae) are vectors of both Rickettsia rickettsii and R. parkeri in the Americas, where capybaras (Hydrochoerus hydrochaeris) are the main hosts in urban areas, thus contributing to the transmission of spotted fever rickettsiosis [1–4]. Amblyomma ticks frequently infest capybaras [5,6], and Amblyomma sculptum Berlese 1888 is the most important vector of Brazilian Spotted Fever (BSF), a severe disease caused by R. rickettsii [7–10]. A milder form of human rickettsiosis caused by R. parkeri has also been reported in Brazil [1].
Between 2007 and 2021, Brazil has reported around 36,500 suspected cases of BSF, with 7% being confirmed, resulting in approximately 170 cases annually and a total of 837 deaths [11]. Reports have shown most cases in the southern and southeastern regions [12,13]. However, several studies have found ticks, capybaras, and other animals infected with Rickettsia in the central-western region [14,15]. In Brasília, one confirmed case of BSF was reported [11], and a study of ticks associated with capybaras on the shore of the Lake Paranoá found A. dubitatum Neumann 1899 infected with *Rickettsia parkeri-*like agent [15]. Additionally, 53 out of 55 capybara serum samples tested by indirect immunofluorescence reaction were positive for Rickettsia, of which 21 have antigen of Rickettsia bellii. Moreover, PCR revealed R. bellii in 25 out of 108 (23.1%) tick samples collected from capybaras [15].
Although previous studies support that Amblyomma ticks are infected with Rickettsia in our study area, the factors that influence tick abundance and infection in areas occupied by capybaras in Brasília are unknown. Several factors may influence tick abundance and infection in urban areas, such as vegetation density [16], climate [17], host availability [4,18–21], human behavior [22,23], and control measures [24,25]. Then, it is necessary to understand ecological factors influencing tick abundance and infection in urban areas where capybaras occur to estimate the potential risk of Rickettsia transmission to humans and to guide control measures, contributing to the surveillance of BSF. Herein, we studied: (i) seasonal dynamics and abundance of ticks in areas where capybaras live in Brasília, (ii) the effect of environmental variables on tick abundance, and (iii) the presence of Rickettsia-infected ticks.
Materials and methods
Study area
Lake Paranoá covers 37.5 km^2^ in an environmental protection area in Brasília [26] (Fig 1). The lake shore vegetation consists of a mixture of exotic vegetation, cerrado (Brazilian savanna) remnants, and gallery forests [27]. The region is classified as having a tropical savanna climate with a dry winter. The seasonality is marked, with a dry season from May to September and a wet season from October to April [28]. Brasília has an average annual rainfall of 1500 to 1800 mm [29]. Lake Paranoá is an artificial lake designed to increase the relative humidity of the air in Brasilia. It serves as an important leisure spot for the population of Brasília. The shores of the lake offer restaurants, entertainment, and sports facilities.
Study area on Lake Paranoá in Brasília, Federal District of Brazil.Shapefile: Instituto Brasileiro de Geografia e Estatística (IBGE–URL: https://www.ibge.gov.br/) and Instituto de Pesquisa e Estatística do Distrito Federal (IPEDF—URL: https://catalogo.ipe.df.gov.br/layers/geonode_data:geonode:Lagos_e_reservatorios).
Tick collection and identification
We used the dragging technique [30] to collect ticks. We sampled different sites around the Lake Paranoá during six periods (variable season): September 2021, November 2021, February/March 2022, April 2022, June 2022, and September 2022. We randomly chose sampling sites and did all collections in the morning. To access the sampling sites near the lake shore, we used a motorboat. Our sampling effort ranged from 5 to 6 days per period.
We used soft and terry cloth (50cm x 60cm) attached to cleaning rods to collect ticks. We dragged the rod in a delimited area of 6m x 10m. In each site, we utilized twelve pieces of cloth (six made of soft material and six made of terry cloth) to prevent losing ticks during the process (Fig 2). The collected ticks were preserved on-site in 70% alcohol, stored in previously identified sealed plastic bags, and transported to the laboratory for identification under a stereomicroscope.
Sampling of ticks on the shore of Lake Paranoá, Brasília, Federal District, Brazil, 2021/2022.S: Soft cloth. T: terry cloth. The picture displays dark green bushes, trees, and ticks on different parts of the landscape.
Tick identification and Rickettsia research
Each tick was assigned to one of three life stages: larva, nymph, or adult [5]. Larvae were identified at the genus level using external morphology, and nymphs and adults were identified at the species level using printed keys [3,5,31–33]. For confirmation/specific identification of ticks and Rickettsia research, 1,756 individuals collected from September 2021 to April 2022 were analyzed either individually (adults) or in pools of 5 nymphs and 10 larvae, separately. To extract genomic DNA (gDNA) from tick samples, we used the saturated sodium chloride solution protocol [34]. We identified the genotypes of these tick samples by using PCR amplification of a fragment of the mitochondrial 16S rDNA gene [35].
Rickettsia was found in the ticks by amplifying fragments of gltA—citrate synthase [36] and ompA—outer membrane protein A [37] in a screening PCR. Afterwards, we amplified fragments of the htrA gene, which encodes the 17-kilodalton structural membrane protein (nested PCR) [38,39], and the ompB gene, which encodes the outer membrane protein B (nested PCR) [40] on positive samples. Each test used 300 ng of R. rickettsii DNA as a positive control and ultrapure water without DNase and RNase as a negative control. We carried out all PCR reactions using a GeneAmp PCR System 9700 thermocycler (Applied Biosystems®, Carlsbad, USA).
PCR products were visualized by electrophoresis in 2% agarose gel stained with ethidium bromide and purified using the Wizard® SV Gel and PCR Clean-Up System Kit (PromegaCorp., Madison,WI,USA) according to the manufacturer’s protocols. DNA was sequenced in both directions using the same PCR primers and the BigDye Terminator™version3.1 Cycle Sequencing® Kit (Applied Biosystems, Foster City, CA, USA) on an automated ABI3730xl DNA analyzer (Applied Biosystems, Foster City, CA, USA). All obtained sequences are available on GenBank (accession numbers OR760224- OR760264; OR767220- OR767227).
The sequences were assembled and edited using ChromasPro, version 1.5 (Technelysium Pty Ltd., Tewantin, QLD, Australia) and subjected to BLASTn analyses (https://blast.ncbi.nlm.nih.gov/Blast.cgi) to identify first their closest similarities to other organisms available in GenBank. For Rickettsia sequences, we performed multiple alignments with the ClustalW algorithm and manually verified them. Protein genes were turned into amino acids without any stop codons. A combined Maximum-Likelihood phylogeny (gltA+ htrA+ ompB) was determined using PhyML Software Version 3.0 [41]. The evolutionary model GTR+I indicated by MEGA 6.0 was used through BIC. Alignments were concatenated with Seaview Software [42].
Environmental characterization
At each sampling site, the following environmental variables were recorded: vegetation type (veg), canopy density (can), temperature (t), relative air humidity (rh), and the presence or vestige of capybaras (capves), identified through their scat, beds, and tracks. We identified three types of vegetation: (1) grass; (2) shrubs and trees; and (3) exposed soil, grass, and shrubs. The measurement of t and rh was carried out with a multimeter (Instrutherm, Thal-300) and can was quantified using a convex spherical forest densitometer (Forestry Suppliers, model C).
Capybara count
Due to the presence of capybaras in our region, a species usually associated with the presence of ticks [15,43], we mapped the presence of capybaras along the shore of Lake Paranoá during the study period. The capybaras were counted using a boat (an aluminum boat with an outboard motor) at about 20 km/h at approximately 30 m from the shore. These surveys were carried out monthly during the late afternoon (16:00 to 18:00). During the surveys, the presence of the capybaras and their geographical location were recorded.
Statistical analysis
Statistical analyses were performed in the R statistical environment, version 4.2.3 [44], using the Performance Analytics, pscl, car, lmtest and KernSmooth packages. We ran exploratory analyses to test for variation in the number of ticks across life stages (larvae, nymphs, or adults), species, seasons, drag type, vegetation type, canopy density, temperature, relative air humidity, and presence or vestige of capybaras. To understand the relationship between the presence of capybaras and the abundance of ticks recorded, we created a kernel density map [45], using the KernSmooth package [46], with the records of capybaras for each month during the study. We then extracted the kernel intensity values (capker) for each tick collection site, according to the month of sampling. The map that describes the study area was generated using QGIS software version 3.28.13.
We analyzed the effect of environmental variables on tick abundances for each life stage by fitting Generalized Linear Models (GLMs), following the recommendations of Zuur et al. [47]. We ran an initial GLM including cloth type (soft or terry) as an explanatory variable and found no effect of this variable, so we used the total number of ticks (soft + terry) as the dependent variable in further analyses. The number of ticks is a count variable, indicating the use of a GLM with a Poisson error distribution and a log link function (Poisson GLM). In the case of moderate overdispersion (1.5 < φ < 15), we corrected the standard errors using a quasi-GLM model in which the variance is given by the mean multiplied by the dispersion parameter φ (quasi-Poisson GLM). In cases in which overdispersion in the data was high (φ > 15) and the frequency of zeros in the data was much higher than expected from a Poisson distribution, we used zero-inflated GLMs, "mixture" models in which the zeros are modeled as coming from two different processes: the binomial process (to model the probability of measuring a zero) and the count process, modeled by a Poisson (ZIP) or a negative binomial (ZINB) GLM [47]. In these cases, we used simple inflation models, where all zero counts have the same probability of belonging to the zero component. To assess if overdispersion was adequately considered, we compared corresponding ZIP and ZINB models by applying a likelihood ratio test.
The environmental explanatory variables considered were veg, season, can, t, rh, capves and capker. Data used are available in S1 Data. The collinearity among explanatory variables was analyzed using variance inflation factors (VIF) as implemented in the car package, considering 5 as the cutoff value (variables with VIF > 5 were excluded from the analysis). For each life stage, variable selection to find the optimal model was performed by applying backward procedures, systematically removing variables that did not contribute significantly (p > 0.05) to the model. For quasi-Poisson GLM, we used an appropriate analysis of deviance to compare nested models with the full model, and for ZINB, likelihood ratio tests were employed (S1 File).
Ethics approval and consent to participate
The adopted procedures were in accordance with the ethical standards of the Research Ethics Committee of the Fiocruz (Rio de Janeiro, Brazil) and with the Helsinki Declaration of 1964, revised in 1975, 1983, 1989, 1996, and 2000. Access to the research site was authorized by the Department of Environment of the Federal District. Permission to collect ticks was granted by the Chico Mendes Institute for Biodiversity Conservation (ICMBio) through the Biodiversity Authorization and Information System (SISBIO), request number 77851, authentication code 0778510320220711.
Results
We collected 30,334 ticks from 115 of 194 sampled sites. Most ticks were captured during the larval stage (Table 1). Morphological identification confirmed that adult/nymphs were Amblyomma, mainly A. sculptum; genotypic identification of larvae showed that 95% were A. dubitatum (Table 2).
Table 1: Number of ticks captured on the Lake Paranoá shore in Brasília, Federal District of Brazil, between 2021 and 2022.
Table 2: Tick species captured on the shore of Lake Paranoá, Brasília, Federal District of Brazil, between 2021 and 2022, by species and life stage.
Tick life stages were detected in all sampling periods. The larvae were the most abundant life stage; most of them were found in April and June 2022, during the early dry season. Nymphs were more abundant in June of 2022 during the dry season, while adults were more common in November of 2021 during the wet season (Fig 3).
Average tick abundance.A, larvae; B, nymphs and adults. Number of individuals per site found on the shores of Lake Paranoá in the months of September and November 2021, and February, April, June and September 2022, and total rainfall for the collection periods.
During the initial analysis of larval abundance, a Poisson GLM revealed substantial overdispersion (φ = 227.3), leading to the use of zero-inflated models. A ZIP GLM including all the explanatory variables exhibited high VIF values (> 14) for t and rh. After excluding rh, the VIF for t was reduced to 6.72. However, due to persisting collinearity concerns, t was also excluded from further analyses. The optimal model was a ZINB GLM, which retained only the variable season. Jun2022 had a significantly higher larval count compared to sep2021 (p < 0.001), nov2021 (p < 0.001), feb2022 (p < 0.001), and sep2022 (p < 0.001). Variables veg, can, capves and capker showed no significant effect and were excluded from the optimal model during the selection procedure (S1 File).
For the analysis of nymphs, we identified moderate overdispersion (φ = 9.92), which required the use of a quasi-Poisson GLM. The optimal model retained the variables season, t, and capves; however, only the season variable showed a significant effect. Specifically, the data from jun2022 demonstrated a significantly higher number of individuals compared to sep2021 (p = 0.003), nov2021 (p < 0.001), apr2022 (p < 0.001), and sep2022. Variables veg, can, rh and capker showed no significant effect and were excluded from the optimal model during the selection procedure (p < 0.001) (S1 File).
For adult specimens, we observed moderate overdispersion (φ = 1.84), which led to the adoption of a quasi-Poisson GLM. The optimal model indicated a negative effect of rh (p = 0.015) and an effect of season, with nov2021 having a significantly higher number of individuals compared to sep2021 (p = 0.002), apr2022 (p = 0.048), jun2022 (p = 0.006), and sep2022 (p = 0.002). Variables veg, can, t and capves showed no significant effect and were excluded from the optimal model during the selection procedure (S1 File). The optimal model included the variable capker, but it did not show a significant effect (p = 0.092). Moreover, no clear agreement was observed when comparing the heat map of capybara distribution in Lake Paranoá throughout the year with the distribution of ticks captured in the same areas; low capybara densities were observed in some areas with high tick abundance (Fig 4).
Distribution of capybara and Amblyomma ticks at Lake Paranoá, Brasilia, Brazil.The heat map represents the kernel density based on the groupings of capybaras observed during the 12 months of sampling. Warm colors represent places where capybaras are more abundant. The size of the circles represents the abundance of ticks counted during the study. Shapefile: Instituto Brasileiro de Geografia e Estatística (IBGE–URL: https://www.ibge.gov.br/).
We tested 254 pool tick samples for Rickettsia, of which 37 (14.5%) were positive (in all months, mainly in November 2023, S1 Table) for the gltA gene, indicating the presence of bacteria of the genus Rickettsia. The positive samples were from larvae (mainly A. dubitatum), nymphs (4 A. dubitatum and 1 A. sculptum), and adults (1 A. sculptum). Three of these pools were also amplified for the htrA and ompB genes (September), indicating the presence of bacteria of the spotted fever group (SFG) (S1 Table). The concatenated phylogeny of the gltA, htrA, and ompB rickettsial gene fragments showed detection of Rickettsia infection in Amblyomma samples. The ompA gene fragment was not amplified. Three larvae samples of A. dubitatum (01G, 10D and 10E, S1 Table) were detected infected with the R. parkeri group. Additionally, 34 pools were identified with R. bellii (all the obtained sequences were 100% similar and classified as Haplotype I) (Fig 5).
Phylogenetic results of Rickettsia spp.Concatenated phylogeny of the gltA, htrA, and ompB rickettsial gene fragments (380+409+403 bp) detected in Amblyomma samples in this study, inferred by maximum likelihood analysis with the GTR+I evolutionary model. The numbers on the branches represent the support values (70% cut-off). Stars (★) indicate sequences obtained in this study.
Discussion
Ticks collected from the shore of Lake Paranoá in Brasília all belonged to the Amblyomma genus, specifically A. sculptum and A. dubitatum. During the dry season, larvae and nymphs were more abundant, while adult ticks were primarily detected in the wet season. Vegetation variables available at the shore of Lake Paranoá showed no association with tick abundance. Surprisingly, the presence or vestige of capybaras showed no effect on the number of ticks of all life stages. Rickettsia sp. was detected in 14.5% of the samples analyzed. Of these, 92% were identified as R. bellii and 8% as R. parkeri group. The exclusive presence of R. parkeri in A. dubitatum ticks suggests that this tick species may be the potential vector of this spotted fever group bacteria in Brasília.
Cloth-dragging is a commonly used and cost-effective technique for sampling ticks [48]. In this study, we chose this method due to its convenience, considering the high number of sites visited during each sampling period. To maximize tick collection, we modified the standard technique by using clothes with different textures and drag multiple smaller cloth pieces on the vegetation. This method has proven effective for dragging over the dense vegetation of Cerrado, where a large cloth would tear easily. Our findings showed that both soft and terry cloth are suitable for collecting ticks of different life stages without distinction. Nonetheless, we noted a considerable number of absences throughout all sampling periods. Combining dragging with other collection methods, such as visual inspection and CO₂ traps, has been noted as a viable way to maximize tick collection [30,49]. However, Ramos et al. [50] found that dragging with soft cotton cloth was more efficient than visual inspection and CO₂ traps. Moreover, Queiroz [51] observed that dragging is more efficient for sampling A. sculptum due to its stalking behavior on vegetation, while the use of CO₂ traps would be more appropriate for sampling A. dubitatum due to its attacking behavior. We believe that the use of other sampling methods in addition to the drag technique would allow us to collect a greater richness and abundance of species. Additionally, some collection points produced clusters of larvae, which could have introduced bias into our study due to the likelihood that these larvae belong predominantly to one species.
We found a higher abundance of A. sculptum adults and nymphs compared to A. dubitatum, in contrast to the results of Quadros et al. [15] in the same location, who found a higher abundance of A. dubitatum compared to A. sculptum in capybaras. This disparity may be because Quadros et al. [15] collected ticks directly from capybaras rather than from the vegetation. The sampling-effect hypothesis is supported by Paula et al. [52], who sampled ticks from the vegetation using dragging, flagging, and visual search. They found a lower frequency of A. dubitatum compared to A. sculptum in the Federal University of Goiás campus, where capybaras, coatis, cattle, and dogs are present. Queirogas et al. [18] suggest that A. dubitatum is more common in areas near water, associated with the presence of capybaras, while A. sculptum prefers drier areas further from the shore. This provides new insight and nuance to Queirogas et al. [18] proposal, suggesting that A. dubitatum, although associated with capybaras, is more restricted to less harsh conditions than A. sculptum. The effect of vegetation characteristics on tick abundance has been reported previously. In a study analyzing Ixodes scapularis using flag/drag techniques within a forested environment, Ginsberg et al. [53] discovered that canopy cover was a more reliable predictor of tick abundance than other vegetation characteristics. These results suggest that the effect of vegetation on tick abundance may depend on the species and the ecosystems in which they live; our results indicate that the vegetation variables we measured on the shores of Lake Paranoá do not significantly influence the abundance of Amblyomma ticks.
The seasonal influence on tick abundance observed in Lake Paranoá has also been reported in the literature. High abundance of larvae/nymphs of Amblyomma ticks has been described during the dry season in a riparian forest in São Paulo, an ecotone region between Atlantic Forest and Cerrado [54], as well as in other parts of São Paulo State [55] and in Cerrado areas of Goiás [50]. Furthermore, the high number of adults during the wet season is consistent with the seasonal dynamics described for A. sculptum in the southern, southeastern, and central-western regions of Brazil [4,52,56,57]. In these regions, larvae are abundant between April and July, nymphs between July and October, and adults between November and February. The rainfall pattern and climatic conditions that imply high temperature and humidity may be responsible for the rapid completion of the tick life stage and increase in adult population [54,58].
Our GLM results revealed that all life stages showed no significant effect of presence or vestiges of capybara. This was an unexpected result considering that A. sculptum and A. dubitatum are commonly associated with capybaras [18,59]. The presence of adult ticks (A. dubitatum and A. sculptum) on capybaras has been reported in our study area [15], suggesting that capybaras are hosts for the maintenance of the life cycle of Amblyomma on the shores of Lake Paranoá. The association between Amblyomma ticks and capybaras was also observed by Nunes et al. [25] in São Paulo. After the culling of capybaras, the number of both A. dubitatum and A. sculptum dropped to almost zero, and the low abundance of adult A. sculptum ticks coincided with relatively low capybara numbers. Queirogas et al. [18] also found a positive correlation between capybara and tick abundance; however, the tick species had an uneven distribution and environmental factors rather than host availability should influence tick abundance. One hypothesis to explain the lack of a positive association between tick abundance and capybara found here could be related to the diversity of food sources. Amblyomma ticks on the shore of Lake Paranoá may rely on other mammalian and avian hosts besides capybaras to complete their life cycle, such as Cavia aperea, Dasyprocta azarae, Vanellus chilensis, Callithrix penicillata, Didelphis albiventris, Gracilinanus agilis, domestic dogs and cats, all animals observed in the shore during sampling. To investigate this issue in depth, molecular techniques would ideally be used to analyze the food sources of the collected ticks [60]. In this sense, it is reasonable to consider other animals besides capybaras as responsible for maintaining the circulation of R. rickettsii in BSF-endemic areas. The shores of Lake Paranoá are occupied by a variety of species, ranging from residential and protected areas to clubs and parks, which are visited daily by humans and domestic animals. These animals may have occasional contact with A. sculptum and carry ticks into residences, highlighting the role of pets as possible carriers of BSF vectors into human settlements [61]. We emphasized that the number of A. sculptum ticks may be influenced by the number of domestic animals, especially dogs and cats [62,63], and therefore we recommend that further studies in the Lake Paranoá consider the frequency of domestic animals in addition to capybaras presence.
The presence of R. bellii in ticks collected directly from capybaras near recreational sites was previously observed in Brasília [15]. R. bellii is commonly found in ticks and is not pathogenic to humans [64,65]. However, R. parkeri, a pathogenic bacterium of the spotted fever group, has been reported in capybaras in Brasília [15]. R. parkeri has also been reported in southern and southeastern Brazil, associated with Amblyomma ovale in Mato Grosso do Sul [66], A. tigrinum in the Pampas [67], and A. triste in Minas Gerais [68]. Recently, a new focus of spotted fever caused by R. parkeri has been described in Rio de Janeiro. Martiniano et al. [69] detected R. parkeri in human skin and in A. ovale from a dog, expanding the known range of this rickettsial disease in Brazil.
The BSF-endemic areas were characterized by much higher tick burdens on both capybaras and in the environment, when compared to the BSF-nonendemic areas [8]. Additionally, there is no serologic evidence of R. rickettsii infection in capybaras found in BSF-nonendemic areas, where capybaras have been found to be infected with other Rickettsia species, particularly R. bellii which may be related to the dominance of A. dubitatum ticks in [8]. Our results support the suggestion of Quadros et al. [15] that the Federal District of Brazil is not currently endemic for BSF, because none of the samples of A. sculptum or A. dubitatum tested positive for R. rickettsii. The non-detection of R. rickettsia in Brasília may be due to a low infection rate of capybaras and ticks, as observed in other regions of Brazil, where capybaras show 0.05%-1.28% infection [7]. At the same time, the uneven distribution of tick species might implicate an unequal risk of tick-borne diseases within the same urban area [18]. A. sculptum is the most frequent human-biting tick in southeastern Brazil and is the most important vector of BSF [7]. Furthermore, A. dubitatum has been commonly found to parasitize capybaras in southeastern Brazil, without any direct impact on BSF-epidemiology [70,71]. In São Paulo, A. dubitatum was found to be heavily infected with R. bellii [8], and experimental studies showed that A. dubitatum infected with R. bellii was only partially refractory to R. rickettsii and unable to transmit R. rickettsii transovarially [72]. These findings suggest that if A. dubitatum becomes prevalent in a particular area, R. rickettsii may not be able to establish an effective infection in either A. dubitatum or A. sculptum. In our study, it seems that most of the larvae captured in the environment were A. dubitatum, which could also explain why R. rickettsii was not detected in examined ticks.
The endemism of BSF in an area can be attributed to the size of the capybara population. In 2006, a study was conducted in São Paulo before the endemism of BSF, where 78 capybaras and few Amblyomma ticks were sampled (0.7 A. sculptum/trap and 3.3 A. dubitatum/trap); after 6 years, this area had become endemic for BSF, and there was an increase in the number of capybaras (~3 times higher) and ticks (33 A. sculptum/trap and 2.1 A. dubitatum/trap) [57]. The study suggests that the emergence of BSF is linked to the growth of the capybara population, which offers a food source for A. sculptum. Consequently, capybara populations in urban areas are associated with high environmental tick infestation. This, in turn, increases the risk of tick bites and pathogen transmission to humans. Our results highlight the importance of monitoring capybaras and ticks (A. dubitatum and A. sculptum) in Lake Paranoá to study their population dynamics [73] and infection thus providing relevant data for the surveillance of R. rickettsii in Brasilia and preventing outbreaks of BSF transmission, which have been observed in the southeastern region of Brazil [1,7,8,10].
Our study indicates that ticks are more influenced by climatic factors than by the presence of capybara. Therefore, prevention measures should focus on avoiding exposure to ticks in the study areas, especially during the dry season. This can be achieved through personal protective measures, such as wearing appropriate clothing, as well as by displaying signs warning residents of the presence of ticks, which are potential vectors of BSF. Health education campaigns can also be conducted to inform people about what to do if they find a tick and when ticks are most abundant in the area [74–76].
Conclusions
We conclude that: (i) Amblyomma ticks are widely distributed in Paranoá Lake throughout the year; (ii) the abundance of Amblyomma ticks is explained more by climatic factors than by vegetation or presence of capybaras; (iii) A. dubitatum ticks are potential vectors of R. parkeri in Brasília. Based on tick infection data, there is a potential risk of Rickettsia transmission to humans in this non-endemic but vulnerable area.
Supporting information
S1 TableRickettsia-infected ticks from Lake Paranoá shore, Brasília, Federal District of Brazil, between 2021 and 2022.(XLSX)
S1 DataData used in the GLM analyses to estimate the effect of environmental variables on tick abundances for each life stage.(XLSX)
S1 FileRcode used to estimate the effect of environmental variables on tick abundances for each life stage.(TXT)
S1 Graphical abstract(PPTX)
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1SzabóMPJ, Pinter A, Labruna MB. Ecology, biology and distribution of spotted-fever tick vectors in Brazil. Front Cell Infect Microbiol. 2013; 3–19.23875178 10.3389/fcimb.2013.00027 PMC 3709097 · doi ↗ · pubmed ↗
- 2Dantas-Torres F, Onofrio VC, Barros-Battesti DM. The ticks (Acari: Ixodida: Argasidae, Ixodidae) of Brazil. Syst Appl Acarol. 2009;14: 30–46.
- 3Dantas-Torres F, Martins TF, Muñoz-Leal S, Onofrio VC, Barros-Battesti DM. Ticks (Ixodida: Argasidae, Ixodidae) of Brazil: updated species checklist and taxonomic keys. Ticks Tick Borne Dis. 2019; 10:1012. doi: 10.1016/j.ttbdis.2019.06.012 31255534 · doi ↗ · pubmed ↗
- 4Dantas-Torres F, Melo MF, Sales KGS, Sousa-Paula LC, Silva FJ, Figueredo LA, et al. Seasonal dynamics and rickettsial infection in free-living Amblyomma dubitatum in the Atlantic forest biome in north-eastern Brazil. Acta Trop. 2021; 217:105854.33561429 10.1016/j.actatropica.2021.105854 · doi ↗ · pubmed ↗
- 5Barros-Battesti DM, Arzua M, Bechara GH. Carrapatos de importância médico-veterinária da Região Neotropical: Um guia ilustrado para identificação de espécies. Vox/ICTTD-3/, Butantan, 2006.
- 6Ramírez-Hernández A, Uchoa F, Serpa MCA, Binder LC, Souza CE, Labruna MB. Capybaras (Hydrochoerus hydrochaeris) as amplifying hosts of Rickettsia rickettsii to Amblyomma sculptum ticks: Evaluation during primary and subsequent exposures to R. rickettsii infection. Ticks Tick Borne Dis 2020; 11(5): 101463.32723630 10.1016/j.ttbdis.2020.101463 · doi ↗ · pubmed ↗
- 7Costa FB, Gerardi M, Binder LC, Benatti HR, Serpa MCA, Lopes B, et al. Rickettsia rickettsii (Rickettsiales: Rickettsiaceae) infecting Amblyomma sculptum (Acari: Ixodidae) ticks and capybaras in a Brazilian spotted fever-endemic area of Brazil. J Med Entomol. 2020; 9;57(1): 308–311.10.1093/jme/tjz 14131504641 · doi ↗ · pubmed ↗
- 8Luz HR, Costa FB, Benatti HR, Ramos VN, Serpa MCA, Martins TF, et al. Epidemiology of capybara-associated Brazilian spotted fever. P Lo S Negl Trop Dis. 2019; 6;13(9): e 0007734. doi: 10.1371/journal.pntd.0007734 31490924 PMC 6750615 · doi ↗ · pubmed ↗