Enpp2 haploinsufficiency induces an eye-open-at-birth phenotype in the DBA/2 background
Seiichi Koike, Kazuko Keino-Masu, Masayuki Masu

TL;DR
Reduced Enpp2 gene activity in mice leads to eyes opening at birth, likely due to maternal effects on embryonic development.
Contribution
Shows that Enpp2 haploinsufficiency in a specific genetic background causes eye-open-at-birth due to eyelid closure failure.
Findings
Enpp2 heterozygous mice in the DBA/2 background frequently exhibit eye-open-at-birth.
Wildtype pups from Enpp2 heterozygous dams also show the phenotype, indicating maternal LPA's role.
The phenotype results from eyelid closure failure during embryonic development.
Abstract
Autotaxin, encoded by the Enpp2 gene, produces lysophosphatidic acid (LPA), which exerts numerous biological functions via its cognate receptors. Enpp2 null mutant mice die by embryonic day 9.5 owing to aberrant vascular development in the yolk sac, preventing analysis after that period. In this study, we found that Enpp2 heterozygous mice in the DBA/2 genetic background showed an eye-open-at-birth phenotype at high frequency, caused by failure of eyelid closure during the embryonic stage. Notably, wildtype pups from the Enpp2 heterozygous dam showed the phenotype, although at lower frequency, suggesting that maternal LPA affects the embryonic development.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
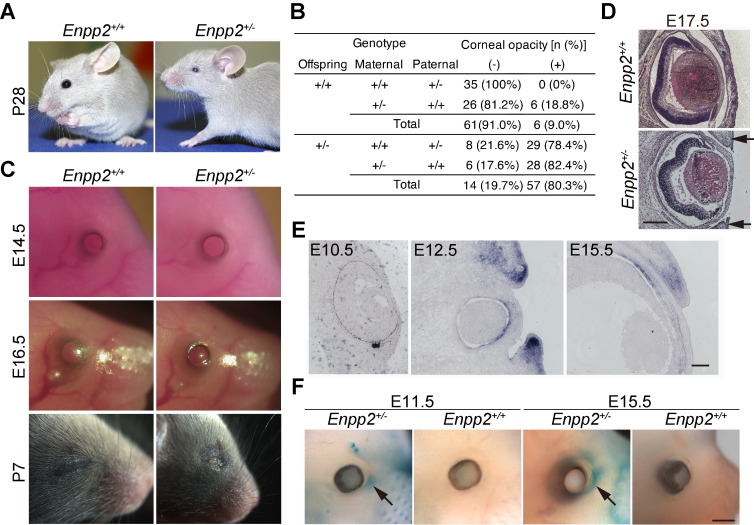 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCongenital heart defects research · Genomics and Rare Diseases · Metabolism and Genetic Disorders
Description
Autotaxin, also known as ectonucleotide pyrophosphatase/phosphodiesterase 2 (Enpp2), is an ectoenzyme responsible for the production of lysophosphatidic acid (LPA) in the extracellular space (Kano et al., 2022; Perrakis and Moolenaar, 2014; Stefan et al., 2005; Tokumura et al., 2002; Umezu-Goto et al., 2002) . LPA is a lipid mediator possessing a wide variety of biological functions, including cell proliferation, migration, and survival (Kano et al., 2022; Luquain et al., 2003) . These effects are mediated by its G protein-coupled receptors (LPA 1–6 ), which activate intracellular signal pathways (Kano et al., 2022; Yung et al., 2014) . Enpp2 ^-/-^ mice exhibit embryonic lethality at embryonic day 9.5 (E9.5) owing to angiogenic defects in the yolk sac (Koike et al., 2009; Tanaka et al., 2006; van Meeteren et al., 2006) . It is thus impossible to investigate the functions of autotaxin after this time period using conventional knockout (KO) mice. To overcome this difficulty, conditional KO mice or transgenic mice were used to examine autotaxin functions in the adult, including inflammation, fibrosis, and obesity (Dusaulcy et al., 2011; Nikitopoulou et al., 2022; Oikonomou et al., 2012) . Additionally, functional analysis has been performed using heterozygous mice, in which the LPA level in the circulation was reduced to approximately 50% of the level of that in wildtype (WT) mice (Tanaka et al., 2006; van Meeteren et al., 2006) : platelet-dependent thrombus formation, suppression of adipocyte hyperplasia, and impaired insulin signaling were reported in Enpp2 heterozygous mice (D’Souza et al., 2018; Nishimura et al., 2014; Pamuklar et al., 2009).
In this study, we examined the impact of genetic background on the Enpp2 mutation because in general, differences in genetic background can weaken or strengthen the phenotypes of KO mice (Montagutelli, 2000) . We backcrossed *Enpp2 * heterozygous mice, maintained on a mixed genetic background of C57BL/6N and 129SvJ, to various inbred strains, including C57BL/6N, 129SvJ, BALB/cA, C3H/HeJ, and DBA/2J. After backcrosses to DBA/2J for 3 consecutive generations, we noticed for the first time that some postnatal mice developed corneal opacity ( Fig. 1A ): none of the mice backcrossed to other inbred strains developed such a phenotype nor showed other visible changes. The frequency of corneal opacity at postnatal day 28 (P28) was about 80% in the * Enpp2 ^+/-^ * mice in the offspring backcrossed to DBA/2J for 4 consecutive generations ( Fig. 1B ). Intriguingly, a small number (8.9%) of the WT mice also showed a similar phenotype ( Fig. 1B ). To eliminate the possibility that a spontaneous mutation affecting eye development occurred during backcrossing, we conducted an independent backcross to DBA/2J and found that the same phenotype appeared at a similar frequency. When examined separately by each parent’s genotype, the WT pups with corneal opacity were observed in those born from * Enpp2 ^+/-^ * females mated with WT males, but not in those born from WT females mated with * Enpp2 ^+/-^ * males ( Fig. 1B ). These data imply that the maternal genotype influences the phenotypic appearance of the offspring.
Previous studies have demonstrated a correlation between corneal opacity and an eye-open-at-birth (EOB) phenotype. In mice, eyelid development begins with the groove extending over the cornea around E12, and the eyelid primordia meet and fuse around E16.5 (Findlater et al., 1993) . The eyelids remain fused until P12-14 when they reopen as a result of apoptosis of the eyelid tissues. When the eyelids are not closed at the embryonic stage, corneal injury induces inflammation and opacity after birh (Nunomura et al., 2021; Wu et al., 2016) . To determine whether the ocular abnormalities observed in Enpp2 ^+/-^ mice are caused by EOB, we examined the eye development of embryos generated by mating WT female mice with * Enpp2 ^+/-^ * male mice to minimize maternal effects. The eyelid primordia were formed in all the embryos at E12.5 to E14.5 ( Fig. 1C ). At E16.5, the eyelids had already fused in the WT embryos, but they remained open in the * Enpp2 ^+/-^ * embryos ( Fig. 1C ). At P7, the eyelids remained closed in the WT mice, whereas the * Enpp2 ^+/-^ * mice showed opened eyelids and corneal opacity ( Fig. 1C ). Histological analysis of transverse slices of the eye at E17.5 revealed that the epithelial sheets over the eye were not formed in the * Enpp2 ^+/-^ * embryos, whereas no apparent abnormalities were observed in the lens, retina, or cornea ( Fig. 1D ). These data indicate that ocular abnormalities observed in postnatal * Enpp2 ^+/-^ * mice resulted from a failure of eyelid extension and closure during the embryonic stage. The corneal opacity seems to be a secondary effect resulting from the injury caused by the absence of eyelids and loss of physical protection.
Finally, we examined the Enpp2 expression in the eye region of the embryos. Initially, we detected *Enpp2 * mRNA using
- in situ* hybridizaton. At E10.5, *Enpp2 * signals were undetectable around the eye ( Fig. 1E ). At E12.5, strong signals were detected in the mesenchymal cells, especially beneath the extending epithelium, but not in the epithelial cells of the eyelids. The expression remained detectable until E15.5. Next, we investigated
- Enpp2* expression by means of wholemount LacZ staining, as the LacZ gene was knocked in the *Enpp2 * KO allele. The signal was weakly detected around the eye at E11.5 and and had become stronger at E15.5 ( Fig. 1F ). The signals were not detected in the WT mice, suggesting the specificity of the lacZ reaction ( Fig. 1F ).
Our data indicate that *Enpp2 * is expressed in the mesenchymal cells of eyelids when the eyelids extend and that decreased
- Enpp2 * expression induces the EOB phenotype in the DBA/2J background, suggesting that Enpp2 is required for the proliferation and/or migration of mesenchymal cells in the eyelids. Why the EOB phenotype appears only in the DBA/2J background is unknown. However, given that DBA/2J is a model for congenital experimental glaucoma (Turner et al., 2017) , the strain may have unidentified susceptibility to eye diseases associated with maldevelopment of the anterior eye segment. Interestingly, heterozygosity of the dam partially induces the EOB phenotype in their WT offspring, suggesting that LPA in the dam somehow affects the eye development of their embryos. Although it also remains unknown whether LPA is supplied from the matermal circulation to the embryo, it is possible that an unidentified LPA transporter in the placenta facilitates transport of LPA as amino acids and fatty acids are transported (Brett et al., 2014) and extracellular LPA can activate peroxisome proliferator-activated receptor (PPARγ) in monocytic cells intracellularly across cell membranes (McIntyre et al., 2003) . Investigating how maternal LPA impacts embryonic development will be necessary in future.
Methods
Animals
All experimental procedures involving animals were approved by the Animal Care and Use Committee of the University of Tsukuba and performed in accordance with its guidelines. Enpp2 KO mice (B6;129SvJ- * Enpp2 ^tm1Mmas^ * ) were generated by use of homologous recombination in 129/Ola-derived ES cells (Koike et al., 2009) . The mutant mice were backcrossed to C57BL/6N, 129SvJ, BALB/cA, C3H/HeJ, and DBA/2J, which were purchased from CLEA Japan. Offspring were genotyped by use of PCR using the primers 5’-CTGCTGAAACTTAATGCACTGGAC-3’ ( Enpp2 forward), 5’-TGTGTAAGTCAGGGAACAACTCTG-3’ ( Enpp2 reverse), and 5’-TGCTCCAGACTGCCTTGGGAAAAG-3’ ( neo ). Noon of the day when a vaginal plug was observed was taken as embryonic day 0.5 (E0.5). Embryos were taken after mice were sacrificed by means of cervical dislocation.
** *In situ * hybridization **
C57BL/6N embryos were fixed with 4% paraformaldehyde (PFA) in phosphate-buffered saline (PBS). After the brains were incubated in 30% sucrose/PBS at 4°C overnight and embedded in OCT compound (Sakura Finetek Japan), 10-µm-thick slices were cut by use of a cryostat CM1850 (Leica). The slices were treated with 1 µg/mL proteinase K in PBS with 0.1% Tween-20 (PBT) at 37°C for 5 min, washed and fixed with 4% PFA, and hybridized with 1 µg/mL digoxigenin (DIG)-labeled antisense RNA probe (nt 678–1323 of mouse *Enpp2 * cDNA; GenBank accession number NM015744 ) in a hybridization solution (50% formamide, 5× SSC pH 4.5, 1% SDS, 50 µg/mL heparin, 50 µg/mL yeast RNA) at 65°C for 16 h. The slides were washed with 50% formamide, 5× SSC, 1% SDS at 65°C for 30 min, and with 50% formamide, 2× SSC at 65°C for 30 min 3 times, and then incubated with an alkaline phosphatase-conjugated anti-DIG antibody (Sigma-Aldrich) at 4°C overnight. After washing with Tris-buffered saline with 0.1% Tween-20, signals were detected by use of BM purple (Sigma-Aldrich) in the presence of 2 mM levamisole (Sigma-Aldrich) at room temperature for 1–5 days.
LacZ staining
Enpp2 ^+/+^ * or * Enpp2 ^+/-^ * embryos were fixed with 2% PFA and 0.2% glutaraldehyde in PBS. After rinsing with PBS 3 times, LacZ staining was carried out by incubation of the embryos in PBS containing 5 mM K 4 Fe(CN) 6 , 5 mM K 3 Fe(CN) 6 , 1 mg/ml X-gal, 2 mM MgCl 2 , 0.02% Nonidet P40, and 0.01% Na deoxycholate.
Histology
For histological examination, mouse embryos were fixed with 4% PFA in PBS. Subsequently, the fixed embryos were processed and embedded in paraffin by use of a tissue processor ASP200 (Leica). Slices of 4-µm thickness were then cut with a microtome RM2145 (Leica) and stained with hematoxylin-eosin (Muto Pure Chemicals).
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Brett KE Ferraro ZM Yockell-Lelievre J Gruslin A Adamo KB 2014912 Maternal-fetal nutrient transport in pregnancy pathologies: the role of the placenta.Int J Mol Sci 159161531618510.3390/ijms 15091615325222554 PMC 4200776 · doi ↗ · pubmed ↗
- 2D'Souza K Nzirorera C Cowie AM Varghese GP Trivedi P Eichmann TO Biswas D Touaibia M Morris AJ Aidinis V Kane DA Pulinilkunnil T Kienesberger PC 201882 Autotaxin-LPA signaling contributes to obesity-induced insulin resistance in muscle and impairs mitochondrial metabolism.J Lipid Res 59100022-22751805181710.1194/jlr.M 08200830072447 PMC 6168304 · doi ↗ · pubmed ↗
- 3Dusaulcy R Rancoule C Grès S Wanecq E Colom A GuignéC van Meeteren LA Moolenaar WH Valet P Saulnier-Blache JS 2011318 Adipose-specific disruption of autotaxin enhances nutritional fattening and reduces plasma lysophosphatidic acid.J Lipid Res 5260022-22751247125510.1194/jlr.M 01498521421848 PMC 3090245 · doi ↗ · pubmed ↗
- 4Findlater GS Mc Dougall RD Kaufman MH 199381 Eyelid development, fusion and subsequent reopening in the mouse.J Anat 183 ( Pt 1)Pt 10021-87821211298270467 PMC 1259860 · pubmed ↗
- 5Kano K Aoki J Hla T 20211123 Lysophospholipid Mediators in Health and Disease.Annu Rev Pathol 171553-400645948310.1146/annurev-pathol-050420-02592934813354 PMC 9641500 · doi ↗ · pubmed ↗
- 6Koike S Keino-Masu K Ohto T Sugiyama F Takahashi S Masu M 2009105 Autotaxin/lysophospholipase D-mediated lysophosphatidic acid signaling is required to form distinctive large lysosomes in the visceral endoderm cells of the mouse yolk sac.J Biol Chem 284480021-9258335613357010.1074/jbc.M 109.01271619808661 PMC 2785199 · doi ↗ · pubmed ↗
- 7Luquain C Sciorra VA Morris AJ 200371 Lysophosphatidic acid signaling: how a small lipid does big things.Trends Biochem Sci 2870968-000437738310.1016/S 0968-0004(03)00139-712878005 · doi ↗ · pubmed ↗
- 8Mc Intyre TM Pontsler AV Silva AR St Hilaire A Xu Y Hinshaw JC Zimmerman GA Hama K Aoki J Arai H Prestwich GD 20021226 Identification of an intracellular receptor for lysophosphatidic acid (LPA): LPA is a transcellular PPA Rgamma agonist.Proc Natl Acad Sci U S A 10010027-842413113610.1073/pnas.013585510012502787 PMC 140905 · doi ↗ · pubmed ↗