Functional analysis of epilepsy-linked pathogenic variants of the Munc18-1 gene in the inhibitory nervous system of Caenorhabditis elegans
Keiko Gengyo-Ando, Akane Osawa-Noguchi, Hideki Ando, Junichi Nakai

TL;DR
This study explores how harmful mutations in the Munc18-1 gene cause epilepsy by examining their effects on nerve cells in a worm model.
Contribution
The study reveals how Munc18-1 pathogenic variants affect inhibitory neurons and correlate with seizure severity in a model organism.
Findings
Three Munc18-1 mutant proteins form aggregates in GABAergic motoneurons, reducing their axonal expression.
Axonal expression defects correlate with pentylenetetrazol-induced convulsions in the worm model.
Mutant protein instability appears to determine the severity of epileptic phenotypes.
Abstract
Heterozygous de novo mutations in Munc18-1, which is essential for neurotransmitter release, cause early infantile epileptic encephalopathy. Munc18-1-linked epilepsy is currently an untreatable disorder and its precise disease mechanism remains elusive. Here, we investigated how Munc18-1 pathogenic variants affect inhibitory neurons using Caenorhabditis elegans . Expression analysis revealed that three missense mutant proteins form aggregates in the cell body of gamma-aminobutyric-acid (GABA)-ergic motoneurons, resulting in a strong reduction of their expression in axons. Their defects of axonal expression correlated closely with pentylenetetrazol-induced convulsions, suggesting that the degree of instability of each mutant protein account for the severity of the epileptic phenotypes.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
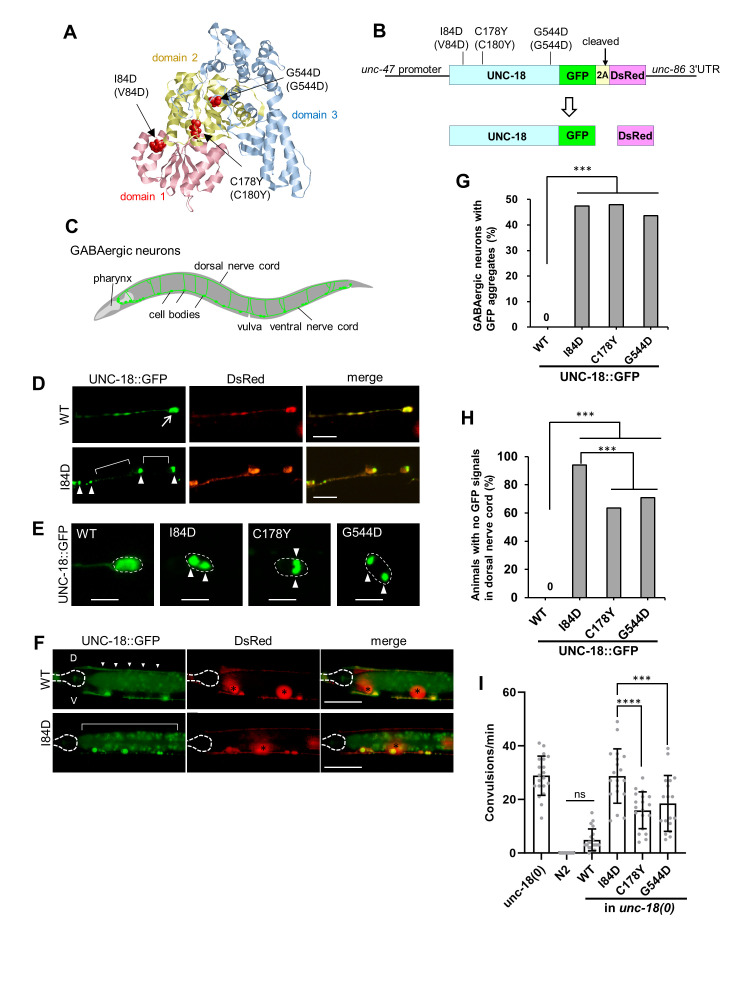 Figure 1
Figure 1|
Strain |
Genotype |
Source |
|
|
|
This study |
|
|
|
This study |
|
|
|
This study |
|
|
|
This study |
|
|
|
This study |
|
|
|
This study |
|
|
|
This study |
|
|
|
This study |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsUbiquitin and proteasome pathways · S100 Proteins and Annexins
Description
Early infantile epileptic encephalopathy (EIEE) with suppression burst, also known as Ohtahara syndrome, is the earliest form of age-dependent epileptic encephalopathy (Yamatogi and Ohtahara, 2002) . Recent high-throughput sequencing studies have identified a number of heterozygous de novo mutations in the Munc18-1 gene in patients with EIEE type 4 (EIEE4), a severe form of epilepsy associated with developmental delay and intellectual disability (Saitsu et al., 2008; Deprez et al . , 2010; Pavone et al., 2012). Currently, seizures in EIEE4 are largely resistant to antiepileptic drugs and there is no targeted therapy for EIEE4. Munc18-1 (also known as STXBP1) encodes a neuronal Sec1/Munc18 protein that regulates synaptic vesicle exocytosis in both vertebrates and invertebrates (Gengyo-Ando et al., 1993; Hata, Slaughter and Südhof, 1993; Harrison et al., 1994). Imbalance between the inhibitory tone and neural excitation, especially perturbation of the inhibitory tone, is strongly suspected as the cause of epilepsy, but how the epilepsy-linked Munc18-1 variants affect inhibitory neurons in vivo is poorly understood. Here, we investigated the effects of pathogenic Munc18-1 missense mutations on protein expression and GABAergic neurotransmission in the C. elegans inhibitory nervous system.
C. elegans UNC-18 (Munc18-1 homologue) was originally isolated by paralyzed phenotype (Brenner, 1974) and is essential for neurotransmitter release including acetylcholine and GABA (Hosono et al., 1992) . We investigated here the three pathogenic Munc18-1 mutations, V84D, C180Y and G544D (I84D, C178Y, G544D for UNC-18 ; Figure 1A ), which were first identified in 2008 in patients with Ohtahara syndrome (Saitsu et al., 2008) . To evaluate the expression of these mutant proteins, we used a 2A peptide-mediated co-expression system, which allows the expression of multiple proteins within a single open reading frame through a co-translational cleavage event (Luke et al . , 2008; Ahier and Jarriault, 2014). We generated transgenic lines expressing GFP-tagged wild-type (WT) or mutant UNC-18 analogous to the pathogenic Munc18-1 mutant with DsRed under the * unc-47 * (a vesicular GABA transporter) promoter ( Figure 1B ). We chose the * unc-47 * promoter so that transgene products would be specifically expressed in GABAergic neurons, i.e., the inhibitory nervous system of C. elegans ( Figure 1C ). We injected these GFP-tagged UNC-18 constructs following our routine protocol in which coelomocyte RFP maker was co-injected. GFP-tagged UNC-18 (WT) showed a soluble distribution pattern throughout the GABAergic motoneurons in the ventral/dorsal nerve cords and cell bodies, similar to soluble DsRed ( Figure 1D upper panels ). On the other hand, GFP-tagged mutant UNC-18 (I84D) exhibited a punctate pattern that differed from the soluble distribution ( Figure 1D lower panels ). This observation is consistent with previous reports showing the aggregation of Munc18-1 mutant proteins in cultured cells and animal models (Chai et al., 2016; Guiberson et al., 2018) . We found that in all three missense mutants, approximately 50% of GABAergic cell bodies in the ventral nerve cord contain aggregates of mutant proteins (n=98-110 from 11-13 animals, Figure 1E and G ). Furthermore, a strong reduction of their expression in the axons was observed in all transgenic strains expressing the mutant proteins ( Figure 1F ). In the dorsal nerve cord, I84D (V84D in Munc18-1) showed the most reduced expression, with no GFP signal detectable in 94.1% of animals ( Figure 1H ).
In order to examine the functional defects of the pathogenic mutants, we generated transgenic lines introducing wild-type (WT) or mutant (I84D, C178Y and G544D) * unc-18 * gene in an * unc-18 ( e81 ) * null mutant background (hereafter referred to as * unc-18 (0) * ) and performed pentylenetetrazole (PTZ)-induced seizure assay. PTZ is a GABA A receptor antagonist and is commonly used to assess defects in GABAergic signaling in animal models, including *C. elegans * (Bessa, Maciel and Rodrigues, 2013) . PTZ treatment induced rapid head seizures in * unc-18 * null mutant animals ( Figure 1I ), while it had no seizure-inducing effect in wild-type N2 animals as described (Williams et al., 2004) . The increased sensitivity to PTZ observed in * unc-18 * *(0) * was completely rescued by * unc-18 * (WT) expression; WT controls ( * unc-18 (0); unc-18 (WT) * ) showed slight convulsions, but not significantly different from wild-type N2 (p=0.29). In contrast, all three mutants showed significantly higher sensitivity to PTZ than the WT control (WT, 4.9±4.1, n=20; I84D, 30.5±12.6, n=20; C178Y, 15.9±6.8, n=19; G544D, 18.5±10.4, n=18; I84D and G544D to WT, p<0.0001; C178Y to WT, p<0.001, Figure 1I ), suggesting that the pathogenic mutants have reduced rescue activity for presynaptic GABA release defects compared to WT. Notably, as in the expression analysis above ( Figure 1H ), I84D (V84D in Munc18-1) again had the most severe effect as compared to the other mutants (I84D to C178Y, p<0.0001; I84D to G544D, p<0.001).
In this study, we showed that the disease-associated Munc18-1 mutant proteins formed aggregates in the cell body of GABAergic motoneurons and that their expression was severely reduced in axons. This result is consistent with the previous findings that the haploinsufficiency of Munc18-1 is one of the major causes for EIEE4 (Saitsu et al., 2010) . Importantly, we observed a close correlation between defects of axonal expression of mutant proteins and the GABA synaptic transmission phenotype. I84D (V84D in Munc18-1) was found to be the most severe mutation in both aspects, which is consistent with previous reports that V84D cause strong phenotypes in both mice and a humanized *C. elegans * model (Kovačević et al., 2018; Zhu et al., 2020). Recently, Munc18-1 haploinsufficiency has also been shown in organotypic slice cultures of the human subplate region; reduction of Munc18-1 by shRNA interference resulted in a reduction of glutamatergic synapse, with a compensatory increase in GABAergic synapse (McLeod et al, 2022) . Thus, Munc18-1 could have roles regulating the number of at least the two types of synapses and, in any case, requires a certain level of expression to exert its proper function. The difference in severity between Munc18-1 mutations may be caused by the degree of protein instability and/or the ability of WT and mutant Munc18-1 to co-aggregate. Recent studies suggest that EIEE4 is caused by a dominant-negative effect of mutant Munc18-1, i.e. by co-aggregation of mutant Munc18-1 with wild-type Munc18-1 or other synaptic proteins (Chai et al., 2016; Guiberson et al., 2018; Lanoue et al., 2019) . The system developed here will be used to further address this question.
Methods
** C. elegans strains and maintenance: ** C. elegans strains were cultured on Nematode Growth Medium (NGM) seeded with E. coli OP50-1 as described (Brenner, 1974) . The wild-type strains Bristol N2 and * unc-18 ( e81 ) * were obtained from the Caenorhabditis Genetics Center. The transgenic strains generated in this study are shown in Reagents.
**Molecular cloning: ** * unc-18 * genomic constructs containing * unc-18 * mutations orthologous to pathogenic Munc18-1 missense mutations were generated as follows. Briefly, amino acid substitutions I84D, C178Y and G544D (V84D, C180Y and G544D in Munc18-1) were introduced into the * unc-18 * genomic clone PE10 (Hosono et al., 1992) by mutating the respective codons using the QuickChange Lightning Site-Directed Mutagenesis Kit (Agilent). For the 2A peptide co-expression system, the *unc-18 (WT)::GFP::T2A::DsRed * fusion cDNA was generated by sewing PCR and cloned into the pFX_EGFPT vector (Gengyo-Ando et al., 2006) with the * unc-47 * promoter (1.2 kb). Enhanced GFP (EGFP) was used for the GFP. I84D, C178Y and G544D were introduced into * unc-18 (WT) * cDNA as described above. All constructs were verified by DNA sequencing.
**Generation of transgenic strains: ** DNA construct (20 ng/µl) and red fluorescent protein (RFP) markers (50 ng/µl) were co-injected into the gonads of N2 hermaphrodites using a standard protocol (Mello et al., 1991) . Stable transgenic lines carrying extrachromosomal arrays were obtained at the F2 generation and maintained. Integration of extrachromosomal arrays was performed by a UV irradiation method (Mitani, 1995) . We used transgenic lines with almost equivalent DsRed intensity between strains in our experiments. Integrated lines were backcrossed four times to N2 and then replaced by crossing into the * unc-18 ( e81 ) * null mutant background. The transgenic strains generated in this study are shown in the reagents.
**Expression analysis: ** We examined aggregates of UNC-18::GFP proteins in the cell bodies where DsRed was expressed. For each strain, 98-110 GABAergic cell bodies in the ventral nerve cord from 10-13 animals were examined and the percentage of cell bodies with aggregates was calculated. We observed UNC-18::GFP in the dorsal nerve cord anterior to the vulva where DsRed was expressed. For each strain, 101-113 animals were examined and the percentage of animals with no detectable GFP signal was calculated.
Imaging: L4 animals were mounted on 2% agarose pads and immobilized with 50 mM sodium azide. A Nikon confocal microscope (A1R, Nikon) with a 20x objective (Plan Apo Lambda, Nikon) and 5x digital zoom or an Olympus microscope (BX50, Olympus) with a 100x objective (UPlan Apo, Olympus) was used to image aggregates in the cell bodies. UNC-18 mutant aggregates were identified as GFP aggregates with a minimum diameter size of 0.5 µm. Images were captured with a Nikon digital camera (DS-Qi1Mc, Nikon). For imaging dorsal nerve cord, a Nikon microscope (ECRIPSE Ni, Nikon) with a 40x objective (Plan Apo Lambda, Nikon) was used. Images were captured with a DP28 digital camera (Olympus).
**Convulsion assay: ** The PTZ assay was performed basically as described (Williams et al., 2004) . NGM plates containing PTZ (Sigma, P6500) at a final concentration of 5 mg/mL were freshly prepared and seeded with OP50-1 . Synchronized young adult animals were placed in the PTZ assay plate for 1 minute. After 1 minute of exposure, head-bobbing convulsions were recorded over 1 minute at 15 frames/sec using an Olympus DP21 digital camera attached to a stereomicroscope (SZX16, Olympus). The convulsions were counted and the number of convulsions per minute was measured. Approximately twenty animals of each strain were observed and analyzed.
3D model : Prediction of the 3D structure of C. elegans UNC-18 was performed using the I-TASSER server (https://zhanggroup.org/I-TASSER/) (Zhang, 2008) . The domain structure of C. elegans UNC-18 was based on the three-domain structure of Sec1/Munc18 proteins defined by Misura et al. (Misura, Scheller and Weis, 2000) . The structure was visualized with RasMol (version 2.7.5.2) ( http://www.openrasmol.org/ ).
Statistics : Differences in percentage of cell bodies with aggregates and differences in percentage of animals with no GFP signals were analyzed using Fisher's exact test in Excel Tokei (Social Survey Research Information) ( Fig. 1G and H ). Statistical significance in the number of convulsions was analyzed using one-way ANOVA followed by Tukey's multiple comparison test in GraphPad Prism 10.0.3 (GraphPad Software) ( Fig. 1I ).
Reagents
**: **
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ahier Arnaud Jarriault Sophie 201431 Simultaneous Expression of Multiple Proteins Under a Single Promoter in Caenorhabditis elegans via a Versatile 2A-Based Toolkit Genetics 19631943-263160561310.1534/genetics.113.16084624361941 PMC 3948794 · doi ↗ · pubmed ↗
- 2Bessa Carlos Maciel Patrícia Rodrigues Ana João 2013314 Using C. elegans to Decipher the Cellular and Molecular Mechanisms Underlying Neurodevelopmental Disorders Molecular Neurobiology 4830893-764846548910.1007/s 12035-013-8434-623494747 · doi ↗ · pubmed ↗
- 3Brenner S 197451 THE GENETICS OF CAENORHABDITIS ELEGANS Genetics 7711943-2631719410.1093/genetics/77.1.714366476 PMC 1213120 · doi ↗ · pubmed ↗
- 4Chai Ye Jin Sierecki Emma Tomatis Vanesa M. Gormal Rachel S. Giles Nichole Morrow Isabel C. Xia Di Götz Jürgen Parton Robert G. Collins Brett M. Gambin Yann Meunier Frédéric A. 201695 Munc 18-1 is a molecular chaperone for α-synuclein, controlling its self-replicating aggregation Journal of Cell Biology 21460021-952570571810.1083/jcb.20151201627597756 PMC 5021092 · doi ↗ · pubmed ↗
- 5Deprez L. Weckhuysen S. Holmgren P. Suls A. Van Dyck T. Goossens D. Del-Favero J. Jansen A. Verhaert K. Lagae L. Jordanova A. Van Coster R. Yendle S. Berkovic S.F. Scheffer I. Ceulemans B. De Jonghe P. 2010928 Clinical spectrum of early-onset epileptic encephalopathies associated with STXBP 1 mutations Neurology 75130028-38781159116510.1212/wnl.0b 013e 3181 f 4d 7bf 20876469 · doi ↗ · pubmed ↗
- 6Gengyo-Ando Keiko Kamiya Yasuko Yamakawa Ayanori Kodaira Ken-ichi Nishiwaki Kiyoji Miwa Johji Hori Isao Hosono Ryuji 1993101 The C. elegans unc-18 gene encodes a protein expressed in motor neurons Neuron 1140896-627370371110.1016/0896-6273(93)90080-b 8398155 · doi ↗ · pubmed ↗
- 7Gengyo-Ando Keiko Yoshina Sawako Inoue Hideshi Mitani Shohei 2006111 An efficient transgenic system by TA cloning vectors and RN Ai for C. elegans Biochemical and Biophysical Research Communications 34940006-291X 1345135010.1016/j.bbrc.2006.08.18316979594 · doi ↗ · pubmed ↗
- 8Guiberson Noah Guy Lewis Pineda André Abramov Debra Kharel Parinati Carnazza Kathryn E. Wragg Rachel T. Dittman Jeremy S. BurréJacqueline 2018928 Mechanism-based rescue of Munc 18-1 dysfunction in varied encephalopathies by chemical chaperones Nature Communications 912041-172310.1038/s 41467-018-06507-4PMC 616222730266908 · doi ↗ · pubmed ↗