The genome sequence of the grey top shell, Steromphala cineraria (Linnaeus, 1758)
Patrick Adkins, Robert Mrowicki, Joanna Harley, Nova Mieszkowska

TL;DR
This paper provides the genome sequence of the grey top shell, Steromphala cineraria, assembled into 18 chromosomal pseudomolecules.
Contribution
The novel contribution is the high-quality genome assembly of Steromphala cineraria, with most of it scaffolded into chromosomal pseudomolecules.
Findings
The genome assembly spans 1,270 megabases.
99.23% of the assembly is organized into 18 chromosomal pseudomolecules.
Abstract
We present a genome assembly from an individual Steromphala cineraria (the grey topshell; Mollusca; Gastropoda; Trochida; Trochidae). The genome sequence is 1,270 megabases in span. The majority of the assembly (99.23%) is scaffolded into 18 chromosomal pseudomolecules.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1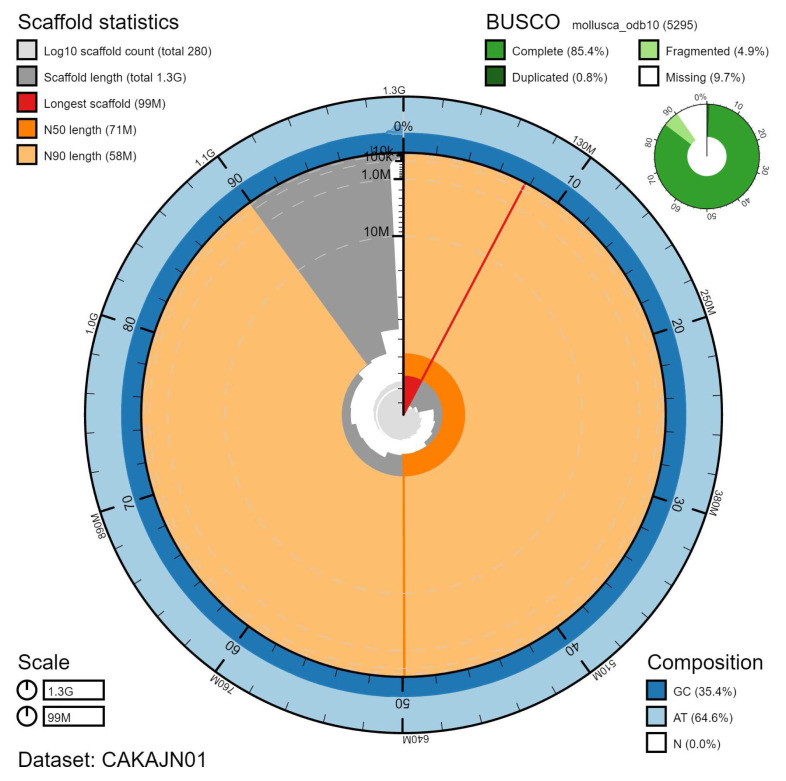 Figure 2
Figure 2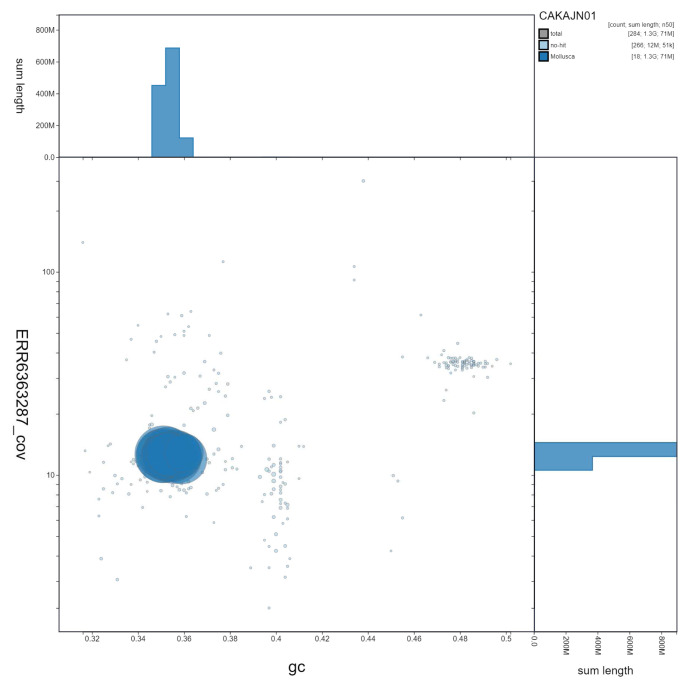 Figure 3
Figure 3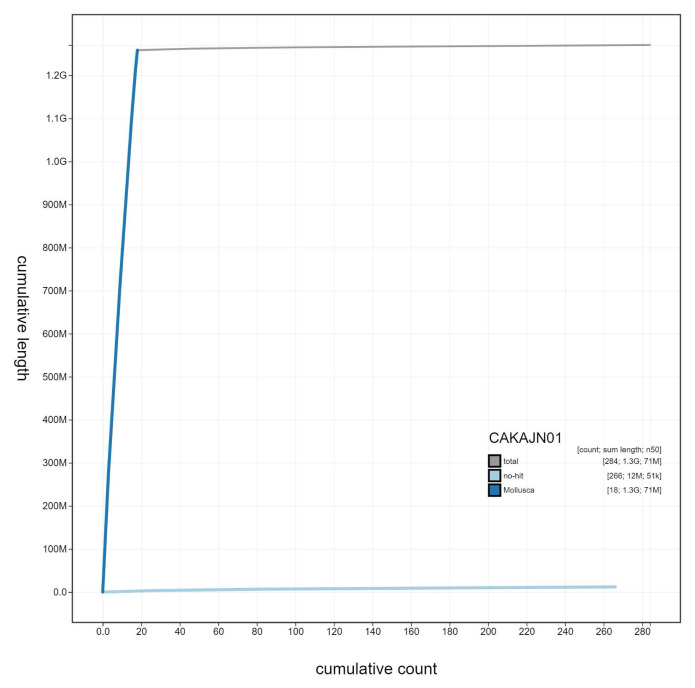 Figure 4
Figure 4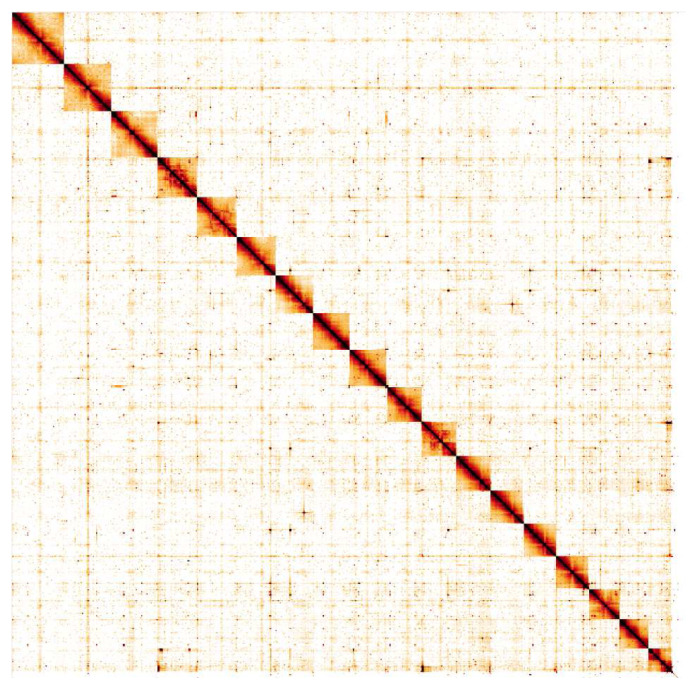 Figure 5
Figure 5|
| |
|---|---|
| Assembly identifier | xgSteCine2.1 |
| Species |
|
| Specimen | xgSteCine2 |
| NCBI taxonomy ID | NCBI:txid216125 |
| BioProject | PRJEB45667 |
| BioSample ID | SAMEA7536348 |
| Isolate information | Muscle |
|
| |
| PacificBiosciences SEQUEL II | ERR6939216, ERR6939217 |
| 10X Genomics Illumina | ERR6363284-ERR6363287 |
| Hi-C Illumina | ERR6363289 |
| PolyA RNA-Seq Illumina | ERR6688409 |
|
| |
| Assembly accession | GCA_916613615.1 |
|
| GCA_916613985.1 |
| Span (Mb) | 1,270 |
| Number of contigs | 842 |
| Contig N50 length (Mb) | 6.2 |
| Number of scaffolds | 283 |
| Scaffold N50 length (Mb) | 70.7 |
| Longest scaffold (Mb) | 98.8 |
| BUSCO
| C:85.4%[S:84.6%,D:0.8%],
|
| INSDC accession | Chromosome | Size (Mb) | GC% |
|---|---|---|---|
| 1 | 98.78 | 35.1 | |
| 2 | 90.22 | 35.1 | |
| 3 | 87.49 | 35.6 | |
| 4 | 76.76 | 35.9 | |
| 5 | 75.56 | 35.5 | |
| 6 | 74.15 | 35.3 | |
| 7 | 70.75 | 35.0 | |
| 8 | 71.35 | 35.3 | |
| 9 | 70.17 | 35.7 | |
| 10 | 66.46 | 35.2 | |
| 11 | 66.15 | 35.5 | |
| 12 | 65.37 | 35.3 | |
| 13 | 62.64 | 35.1 | |
| 14 | 62.10 | 35.2 | |
| 15 | 61.92 | 35.4 | |
| 16 | 57.77 | 35.8 | |
| 17 | 56.52 | 35.3 | |
| 18 | 44.44 | 36.0 | |
| MT | 0.02 | 31.6 | |
| - | Unplaced | 11.89 | 39.8 |
| Software tool | Version | Source |
|---|---|---|
| Hifiasm | 0.15 |
|
| purge_dups | 1.2.5 |
|
| SALSA2 | 3.0 |
|
| longranger align | 2.2.2 |
|
| freebayes | v1.3.1 |
|
| MitoHiFi | 2 |
|
| gEVAL | N/A |
|
| HiGlass | 1.11.6 |
|
| PretextView | 0.2.x |
|
| BlobToolKit | 2.6.4 |
|
- —Wellcome Trust
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGenomics and Phylogenetic Studies · Identification and Quantification in Food · Genetic diversity and population structure
Species taxonomy
Eukaryota; Metazoa; Spiralia; Lophotrochozoa; Mollusca; Gastropoda; Vetigastropoda; Trochida; Trochoidea; Trochidae; Cantharidinae; Steromphala; Steromphala cineraria (Linnaeus, 1758) (NCBI:txid216125).
Background
Steromphala cineraria (Linnaeus, 1758), commonly called the grey topshell, is a gastropod common to rocky shores in the UK. It typically occurs among boulders and cobbles on the lowshore and sub-tidally, where it grazes among Fucus and Laminaria species. Intertidally, it is most common on the lower shore, but can also be found in pools higher on the shore. Sub-tidally it extends to depths of 130 m, although it is most common in the kelp forests between 30 m and low water spring tide ( Fretter & Graham, 1976). Its geographical distribution ranges from southern Portugal and north to the White Sea in northern Russia, becoming rarer at its range edges as thermal limits are approached ( Nekhaev, 2013).
An important grazing species, S. cineraria is distinguished from other species of trochids by its bluntly conical shell and grey/yellowish finely striped patterning on the shell. In smaller shells, the umbilicus is large, becoming smaller and elliptical with age and in large shells sometimes becoming overgrown by the columellar lip ( Fretter & Graham, 1976).
As S. cineraria is found across a large range of latitudes, it is exposed to a wide range of thermal environments in temperature, both due to time of year and geographical distribution. It is important to understand how populations may change in response to climate change, especially in its southern and northern range limits, and the knock-on effects this may have on macroalgae due to changes in grazing populations ( Mieszkowska et al., 2007). A high quality genome sequence for this species will allow future studies to understand more about the mechanisms driving the observed response of this species to a changing climate.
Genome sequence report
The genome was sequenced from a single S. cineraria ( Figure 1) collected from Mount Batten, Devon, UK (latitude 50.36084, longitude -4.12833). A total of 42-fold coverage in Pacific Biosciences single-molecule long reads and 35-fold coverage in 10X Genomics read clouds were generated. Primary assembly contigs were scaffolded with chromosome conformation Hi-C data. Manual assembly curation corrected 408 missing/misjoins and removed 70 haplotypic duplications, reducing the assembly size by 2.51% and the scaffold number by 53.835%, and increasing the scaffold N50 by 123.17%.
Image of the xgSteCine2 specimen.( A) Image taken of the specimen prior to collection. ( B– D) Image of the shell of the specimen following preservation and processing.
The final assembly has a total length of 1,270 Mb in 283 sequence scaffolds with a scaffold N50 of 70.7 Mb ( Table 1). Of the assembly sequence, 99.23% was assigned to 18 chromosomal-level scaffolds (numbered by sequence length) ( Figure 2– Figure 5; Table 2). Large inversions between sister chromatids can be seen on chromosome 5 at 29.7–60.7 Mb and chromosome 11 at 17.7–39.7 Mb. Possible inversions are also seen on chromosome 11 at Mb 3.4–39.4 and 18–66 Mb. The assembly has a BUSCO v5.1.2 ( Manni et al., 2021) completeness of 85.4% (single 84.6%, duplicated 0.8%) using the mollusca_odb10 reference set (n=5295). However, we believe that this relatively low BUSCO score is a result of limitations with the current mollusca_odb10 geneset. Using the metazoa_odb10 reference set (n=954), the assembly has a completeness of 97.6% (single 97.0%, duplicated 0.6%), which we believe is evidence of high completeness. While not fully phased, the assembly deposited is of one haplotype. Contigs corresponding to the second haplotype have also been deposited.
Table 1.: Genome data for Steromphala cineraria, xgSteCine2.1.
Genome assembly of Steromphala cineraria, xgSteCine2.1: metrics.The BlobToolKit Snailplot shows N50 metrics and BUSCO gene completeness. The main plot is divided into 1,000 size-ordered bins around the circumference with each bin representing 0.1% of the 1,270,504,078 bp assembly. The distribution of scaffold lengths is shown in dark grey with the plot radius scaled to the longest scaffold present in the assembly (98,775,408 bp, shown in red). Orange and pale-orange arcs show the N50 and N90 scaffold lengths (70,748,747 and 57,769,559 bp), respectively. The pale grey spiral shows the cumulative scaffold count on a log scale with white scale lines showing successive orders of magnitude. The blue and pale-blue area around the outside of the plot shows the distribution of GC, AT and N percentages in the same bins as the inner plot. A summary of complete, fragmented, duplicated and missing BUSCO genes in the mollusca_odb10 set is shown in the top right. An interactive version of this figure is available at https://blobtoolkit.genomehubs.org/view/Steromphala%20cineraria/dataset/CAKAJN01/snail.
Genome assembly of Steromphala cineraria, xgSteCine2.1. GC coverage.BlobToolKit GC-coverage plot. Scaffolds are coloured by phylum. Circles are sized in proportion to scaffold length. Histograms show the distribution of scaffold length sum along each axis. An interactive version of this figure is available at https://blobtoolkit.genomehubs.org/view/Steromphala%20cineraria/dataset/CAKAJN01/blob.
Genome assembly of Steromphala cineraria, xgSteCine2.1: cumulative sequence.BlobToolKit cumulative sequence plot. The grey line shows cumulative length for all scaffolds. Coloured lines show cumulative lengths of scaffolds assigned to each phylum using the buscogenes taxrule. An interactive version of this figure is available at https://blobtoolkit.genomehubs.org/view/Steromphala%20cineraria/dataset/CAKAJN01/cumulative.
Genome assembly of Steromphala cineraria, xgSteCine2.1: Hi-C contact map.Hi-C contact map of the xgSteCine2.1 assembly, visualised in HiGlass. Chromosomes are shown in size order from left to right and top to bottom.
Table 2.: Chromosomal pseudomolecules in the genome assembly of Steromphala cineraria, xgSteCine2.1.
Methods
Sample acquisition and nucleic acid extraction
A single S. cineraria specimen (xgSteCine2) was collected from Mount Batten, Devon, UK (latitude 50.36084, longitude -4.12833) by Rob Mrowicki (Natural History Museum), Patrick Adkins and Joanna Harley (both Marine Biological Association), by hand. The samples were identified by the same individual and snap-frozen in liquid nitrogen.
DNA was extracted at the Tree of Life laboratory, Wellcome Sanger Institute. The xgSteCine2 sample was weighed and dissected on dry ice with tissue set aside for Hi-C and RNA sequencing. Muscle tissue was cryogenically disrupted to a fine powder using a Covaris cryoPREP Automated Dry Pulveriser, receiving multiple impacts. Fragment size analysis of 0.01–0.5 ng of DNA was then performed using an Agilent FemtoPulse. High molecular weight (HMW) DNA was extracted using the Qiagen MagAttract HMW DNA extraction kit. Low molecular weight DNA was removed from a 200-ng aliquot of extracted DNA using 0.8X AMpure XP purification kit prior to 10X Chromium sequencing; a minimum of 50 ng DNA was submitted for 10X sequencing. HMW DNA was sheared into an average fragment size between 12–20 kb in a Megaruptor 3 system with speed setting 30. Sheared DNA was purified by solid-phase reversible immobilisation using AMPure PB beads with a 1.8X ratio of beads to sample to remove the shorter fragments and concentrate the DNA sample. The concentration of the sheared and purified DNA was assessed using a Nanodrop spectrophotometer and Qubit Fluorometer and Qubit dsDNA High Sensitivity Assay kit. Fragment size distribution was evaluated by running the sample on the FemtoPulse system.
RNA was extracted from muscle tissue in the Tree of Life Laboratory at the WSI using TRIzol, according to the manufacturer’s instructions. RNA was then eluted in 50 μl RNAse-free water and its concentration RNA assessed using a Nanodrop spectrophotometer and Qubit Fluorometer using the Qubit RNA Broad-Range (BR) Assay kit. Analysis of the integrity of the RNA was done using Agilent RNA 6000 Pico Kit and Eukaryotic Total RNA assay.
Sequencing
Pacific Biosciences HiFi circular consensus and 10X Genomics read cloud sequencing libraries were constructed according to the manufacturers’ instructions. Sequencing was performed by the Scientific Operations core at the Wellcome Sanger Institute on Pacific Biosciences SEQUEL II and Illumina NovaSeq 6000 instruments. Hi-C data were generated from additional muscle tissue of xgSteCine2 using the Arima v2.0 kit and sequenced on an Illumina NovaSeq 6000 instrument.
Genome assembly
Assembly was carried out with Hifiasm ( Cheng et al., 2021). Haplotypic duplication was identified and removed with purge_dups ( Guan et al., 2020). One round of polishing was performed by aligning 10X Genomics read data to the assembly with longranger align, calling variants with freebayes ( Garrison & Marth, 2012). The assembly was then scaffolded with Hi-C data ( Rao et al., 2014) using SALSA ( Ghurye et al., 2019). The mitochondrial genome was assembled with MitoHiFi ( Uliano-Silva et al., 2021), which performed annotation using MitoFinder ( Allio et al., 2020). The assembly was checked for contamination and corrected using the gEVAL system ( Chow et al., 2016) as described previously ( Howe et al., 2021). Manual curation ( Howe et al., 2021) was performed using gEVAL, HiGlass ( Kerpedjiev et al., 2018) and Pretext. The genome was analysed within the BlobToolKit environment ( Challis et al., 2020). Table 3 contains a list of all software tool versions used, where appropriate.
Ethics/compliance issues
The materials that have contributed to this genome note have been supplied by a Darwin Tree of Life Partner. The submission of materials by a Darwin Tree of Life Partner is subject to the Darwin Tree of Life Project Sampling Code of Practice. By agreeing with and signing up to the Sampling Code of Practice, the Darwin Tree of Life Partner agrees they will meet the legal and ethical requirements and standards set out within this document in respect of all samples acquired for, and supplied to, the Darwin Tree of Life Project. Each transfer of samples is further undertaken according to a Research Collaboration Agreement or Material Transfer Agreement entered into by the Darwin Tree of Life Partner, Genome Research Limited (operating as the Wellcome Sanger Institute), and in some circumstances other Darwin Tree of Life collaborators.
Data availability
European Nucleotide Archive: Steromphala cineraria (grey top shell). Accession number PRJEB45667; https://identifiers.org/ena.embl/PRJEB45667.
The genome sequence is released openly for reuse. The S. cineraria genome sequencing initiative is part of the Darwin Tree of Life (DToL) project. All raw sequence data and the assembly have been deposited in INSDC databases. The genome will be annotated with the RNA-Seq data and presented through the Ensembl pipeline at the European Bioinformatics Institute. Raw data and assembly accession identifiers are reported in Table 1.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Allio R Schomaker-Bastos A Romiguier J : Mito Finder: Efficient Automated Large-Scale Extraction of Mitogenomic Data in Target Enrichment Phylogenomics. Mol Ecol Resour. 2020;20(4):892–905. 10.1111/1755-0998.13160 32243090 PMC 7497042 · doi ↗ · pubmed ↗
- 2Challis R Richards E Rajan J : Blob Tool Kit - Interactive Quality Assessment of Genome Assemblies. G 3 (Bethesda). 2020;10(4):1361–74. 10.1534/g 3.119.400908 32071071 PMC 7144090 · doi ↗ · pubmed ↗
- 3Cheng H Concepcion GT Feng X : Haplotype-Resolved de Novo Assembly Using Phased Assembly Graphs with Hifiasm. Nat Methods. 2021;18(2):170–75. 10.1038/s 41592-020-01056-5 33526886 PMC 7961889 · doi ↗ · pubmed ↗
- 4Chow W Brugger K Caccamo M : g EVAL - a Web-Based Browser for Evaluating Genome Assemblies. Bioinformatics. 2016;32(16):2508–10. 10.1093/bioinformatics/btw 159 27153597 PMC 4978925 · doi ↗ · pubmed ↗
- 5Fretter V Graham A : The Prosobranch Molluscs of Britain and Denmark: Part 2. Trochacea. The Journal of Molluscan Studies. 1976;42. Reference Source
- 6Garrison E Marth G : Haplotype-Based Variant Detection from Short-Read Sequencing.ar Xiv: 1207.3907,2012. Reference Source
- 7Ghurye J Rhie A Walenz BP : Integrating Hi-C Links with Assembly Graphs for Chromosome-Scale Assembly. P Lo S Comput Biol. 2019;15(8):e 1007273. 10.1371/journal.pcbi.1007273 31433799 PMC 6719893 · doi ↗ · pubmed ↗
- 8Guan D Mc Carthy SA Wood J : Identifying and Removing Haplotypic Duplication in Primary Genome Assemblies. Bioinformatics. 2020;36(9):2896–98. 10.1093/bioinformatics/btaa 025 31971576 PMC 7203741 · doi ↗ · pubmed ↗