Cell-type-specific representation of spatial context in the rat prefrontal cortex
Hans Brünner, Hoseok Kim, Sofie Ährlund-Richter, Josina Anna van Lunteren, Ana Paula Crestani, Konstantinos Meletis, Marie Carlén

TL;DR
The study shows how specific neurons in the rat prefrontal cortex encode spatial information and movement during goal-directed behavior.
Contribution
The research introduces a PV-Cre rat line for studying inhibitory neuron function in spatial encoding.
Findings
PV interneurons in the medial prefrontal cortex prominently encode spatial context.
Gamma band oscillations correlate with movement toward reward but not spatial information.
The PV-Cre rat line allows manipulation of PV neurons in behaving animals.
Abstract
The ability to represent one’s own position in relation to cues, goals, or threats is crucial to successful goal-directed behavior. Using optotagging in knock-in rats expressing Cre recombinase in parvalbumin (PV) neurons (PV-Cre rats), we demonstrate cell-type-specific encoding of spatial and movement variables in the medial prefrontal cortex (mPFC) during goal-directed reward seeking. Single neurons encoded the conjunction of the animal’s spatial position and the run direction, referred to as the spatial context. The spatial context was most prominently represented by the inhibitory PV interneurons. Movement toward the reward was signified by increased local field potential (LFP) oscillations in the gamma band but this LFP signature was not related to the spatial information in the neuronal firing. The results highlight how spatial information is incorporated into cognitive operations…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
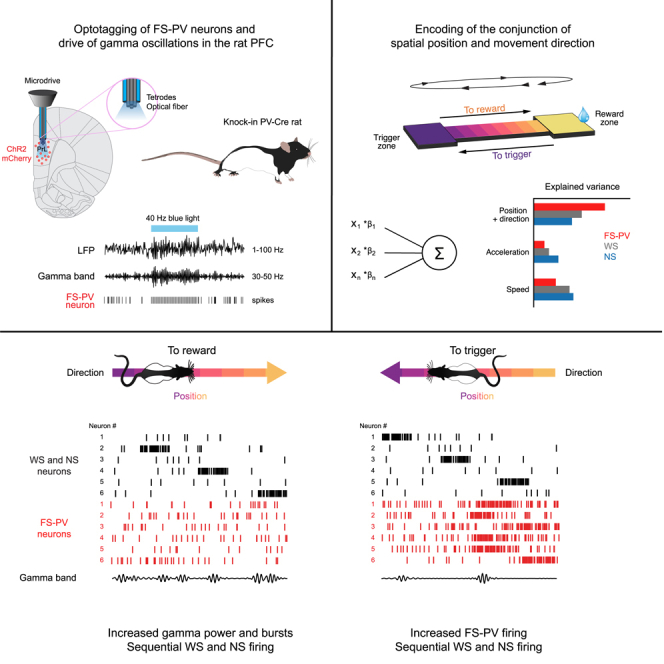 Figure 1
Figure 1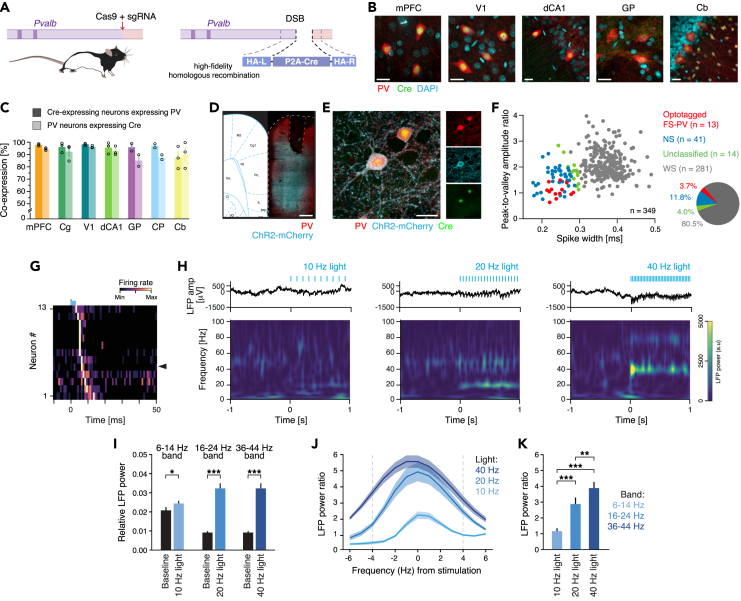 Figure 2
Figure 2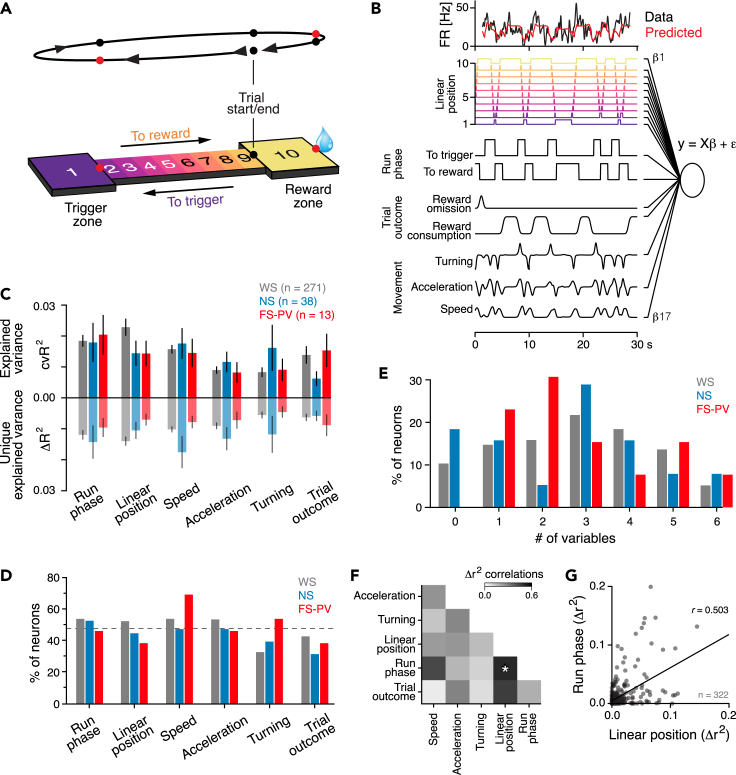 Figure 3
Figure 3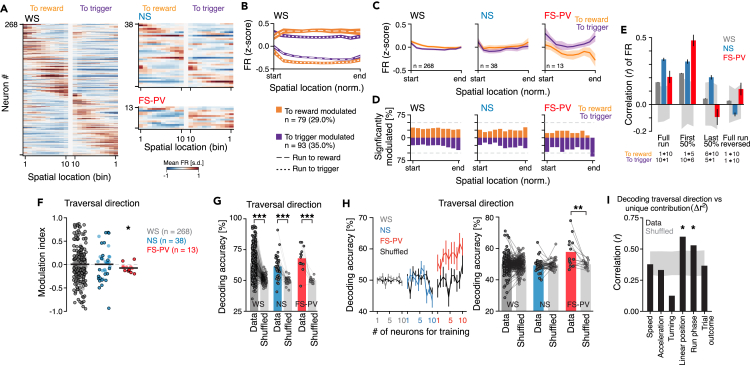 Figure 4
Figure 4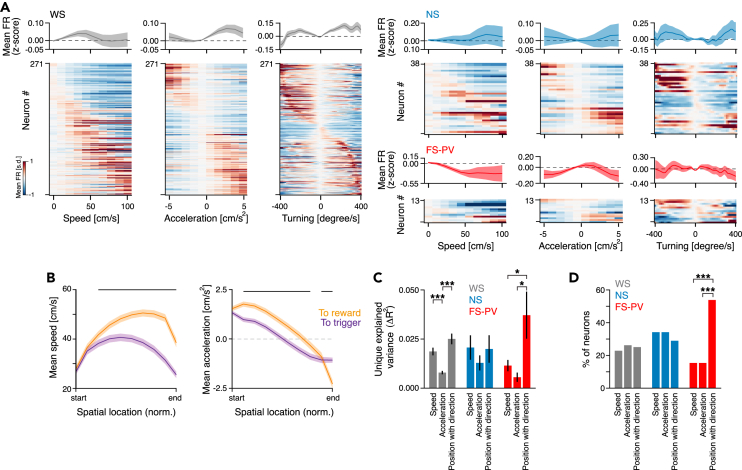 Figure 5
Figure 5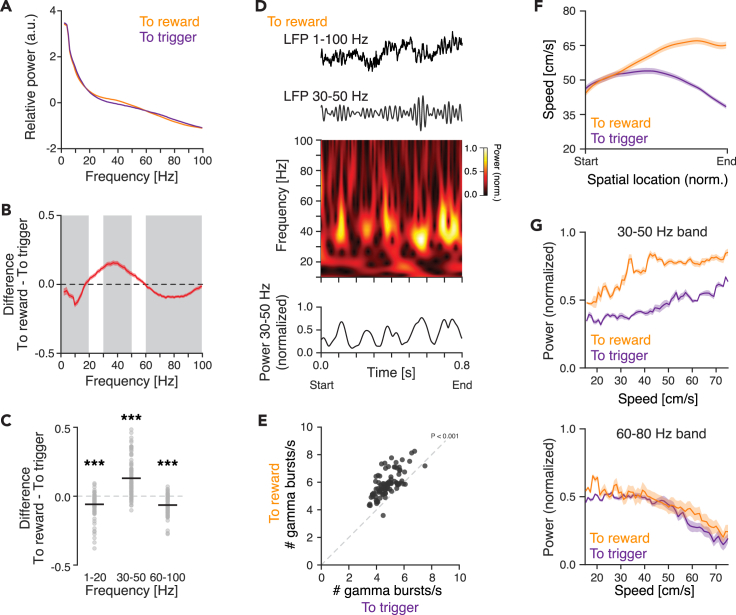 Figure 6
Figure 6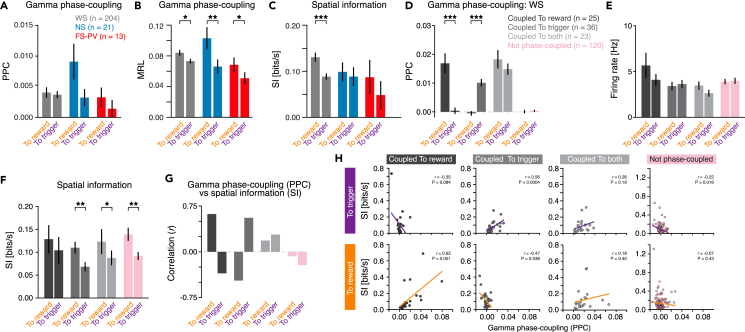 Figure 7
Figure 7Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMemory and Neural Mechanisms · Neuroscience and Neuropharmacology Research · Neural dynamics and brain function
Introduction
Navigating the environment is fundamental to animals' acquisition of sustenance. Navigation decisions incorporate the own localization and place-associated objects, rewards, and threats. The importance of the hippocampal formation in spatial processing has been firmly established, but spatial activation patterns are also widespread in cortical regions.1^,^2^,^3^,^4^,^5 This suggests that spatial variables are diversely utilized in brain functioning, although the specificities remain to be established. The mPFC serves as a hub for integrating different information streams subserving cognitive computations.6 Many cognitive tasks engaging the mPFC require the incorporation of spatial contingencies, and single mPFC neurons have been shown to merge spatial information with other relevant task variables, for example, goals, past choices, or task rules.7^,^8^,^9 Thus, the context and/or relevance of a location appears to be represented in the mPFC.10
Cortical information processing involves dynamic interactions between excitatory and inhibitory neurons organized in network motifs (for review see11^,^12). Interneurons expressing the calcium-binding protein parvalbumin (PV) constitute the largest inhibitory population in both the cortex and the hippocampus.11^,^12 This population is characterized by fast, non-adapting firing and synaptic targeting of perisomatic sites close to the site of action potential (AP) generation in local excitatory neurons. This gives PV interneurons strong influence over the excitatory output and the ability to coordinate local population activities at both behavioral timescales (seconds) and oscillation timescales (milliseconds [ms] to hundreds of ms).13 Over the past 40 years, a wealth of theoretical and experimental research has provided evidence for the central role of fast-spiking PV interneurons (FS-PV neurons) in gamma oscillations (30–80 Hz),14 ubiquitous rhythms that typically emerge in brain regions rich in inhibitory interneurons providing perisomatic inhibition. These transient rhythms appear spontaneously during sleep and waking states but also during distinct processing.15^,^16 Hippocampal gamma rhythms are associated with spatial sequences and movement variables17^,^18 and although gamma rhythms in the PFC have been intensively studied in relation to cognitive operations, the involvement in spatial processing in the PFC is largely unexplored.
Much knowledge on spatial processing, and learning and memory, has been derived from studies in freely moving rats,19^,^20^,^21^,^22 which tolerate more dense and bulky recording technologies than mice. Rats are considered to better reflect human physiology, disease, and behavior than mice,23 but the utility of this model animal has decreased in response to the rapid expansion of the genetic and experimental toolbox for studies in mice. We here introduce PV-Cre rats, a knock-in rat line expressing Cre recombinase in PV neurons throughout the cerebrum and cerebellum. This tool enables ex vivo or in vivo identification and manipulation of select PV populations in rats. To characterize the involvement of FS-PV neurons in prefrontal spatial processing, we employed chronic electrophysiological recordings and optotagging in the prelimbic cortex (PrL) of adult PV-Cre rats engaging in a self-paced, asymmetric reward-seeking task on a linear track. One end of the track was attached to a reward platform and the other end to a trigger platform devoid of reward. To trigger reward delivery in the reward platform, the animals needed to visit the trigger platform, leading the animals to ambulate across the track. Generalized linear models (GLMs) identified significant encoding of both spatial and movement variables in FS-PV neurons, other narrow-spiking (NS) neurons, and wide-spiking (WS) putative excitatory neurons, respectively. The animal’s spatial position on the track was prominently represented in the neuronal activity of all three populations, with subpopulations of single neurons encoding both the animal’s spatial position on the track and if the animal was moving toward or away from the reward. The context of the track traversal was particularly evident in the activities of the FS-PV neurons. Furthermore, the traversals to reward versus (vs.) to trigger were associated with significant differences in the LFP activity in the gamma band (30–50 Hz). However, we did not find support for local gamma oscillations organizing spatial representation in the PrL.
Results
Specific targeting of PV neurons in PV-Cre rats
To allow study and manipulation of PV neurons in rats, we used CRISPR/Cas technology to generate a knock-in rat line with Cre expression in PV neurons (Figure 1A; STAR methods). We used immunohistochemistry as a first step to confirm the targeting of Cre to neurons expressing PV, focusing on a set of brain regions enriched with different types of PV neurons (cortical, subcortical, and cerebellum; Figures 1B, S1A, and S1B). 95.7 ± 0.8% (11858/12368) of the Cre-expressing neurons expressed PV across the seven brain regions, confirming high specificity. Immunohistochemistry also confirmed highly efficient targeting of Cre to the PV populations, with 93.6 ± 1.4% (11858/12641) of the analyzed PV neurons expressing Cre (Figure 1C). The Cre expression was specific and efficient throughout the cortical layers (Figure S1C). PV is a calcium-binding protein, and its expression varies across neurons and brain regions in an activity-dependent manner.24^,^25 This biological variable together with the need for antigen retrieval for Cre detection likely contributed to an underestimation of the overlap between PV and Cre expression. To address functional Cre recombination, an adeno-associated virus (AAV) with Cre-dependent expression of ChR2-mCherry was injected into the mPFC of adult PV-Cre rats (N = 3). This confirmed efficient recombination in PV neurons (94.34 ± 0.79% [742/790] of the opsin-expressing neurons expressed PV; data not shown but see further below).Figure 1. Generation and characterization of the PV-Cre rat line(A) Schematic illustration of the genetic strategy for generation of the PV-Cre knock-in rats. Cre was introduced into the 3′ UTR of the Parvalbumin (Pvalb) locus using a 2A self-cleaving peptide sequence (P2A) between Pvalb and Cre.(B) Example of immunohistochemical detection of Cre (green) in PV-expressing neurons (red) in the cerebrum and cerebellum of adult PV-Cre rats. Nuclei counterstained with DAPI (cyan). For split channels, see Figure S1B.(C) The specificity and efficiency, respectively, of targeting of Cre to PV neurons in seven different brain regions. Dark bars: specificity, i.e., the fraction of Cre-expressing neurons also expressing PV: mPFC (PrL + IL): 97.88 ± 0.54%, Cg (Cg1 + Cg2): 96.09 ± 2.34%, V1: 98.25 ± 0.49%, dCA1: 95.61 ± 2.69%, GP: 95.92 ± 3.06%, CP: 97.12 ± 1.63%, Cb: 87.44 ± 5.23%. Light bars: efficiency, i.e., the fraction of PV neurons expressing Cre: mPFC: 94.39 ± 0.63%, Cg: 92.33 ± 3.81%, V1: 96.25 ± 0.59%, dCA1: 92.75 ± 2.12%, GPe 85.11 ± 4.70%, CP: 87.84 ± 1.94%, Cb: 90.20 ± 6.11.(D) Coronal section from a PV-Cre rat with ChR2-mCherry expression (cyan) in mPFC PV neurons (red). Orange dashed line: outline of the position of optical fiber.(E) Close-up of recombined mPFC PV neurons (red) with ChR2-mCherry expression (cyan) and nuclear expression of Cre (green).(F) Left: plotting and classification of recorded mPFC neurons (n = 349) based on spike waveform properties (spike width and peak-to-valley ratio (PVR), respectively). Three neuronal cell types were classified: WS (gray, n = 281): spike width: 372 ± 35 μs, PVR: 2.1 ± 0.5, FR: 3.6 ± 3.2 Hz; NS (blue, n = 41): spike width: 239 ± 30 μs, PVR: 1.6 ± 0.4, FR: 4.3 ± 6.0 Hz; FS-PV (red, n = 13): spike width: 242 ± 23 μs, PVR: 1.1 ± 0.3, FR: 13.0 ± 6.6 Hz. The light-activated neurons (red, n = 13) display AP characteristics typical of inhibitory FS-PV interneurons, with short spike width and a low PVR. Right: pie chart with the proportions of classified neurons.(G) Averaged and normalized (Min-Max) PSTH aligned to light (3 ms; blue bar) onset for all FS-PV neurons (n = 13). Black arrowhead: the FS-PV neuron in Figure S2H.(H–K) The LFP resonance in response to activation of mPFC FS-PV neurons at 10, 20, or 40 Hz, respectively.(H) Example single LFP traces during baseline (−1 to 0 s) and light stimulation (0–1 s) at 10 (left), 20 (middle), or 40 Hz (right). Top: raw LFP (0.1–500 Hz). Bottom: spectrogram (2–100 Hz). All three stimulation frequencies evoked a response in the LFP visible as an increase in power at the stimulation frequency.(I) Comparison of the relative LFP power (band: ±4 Hz the light stimulation frequency) between baseline (black) and in response to light application (blues).(J) The relative LFP power ratio (Plight/Pbaseline [mean]) in response to activation of mPFC FS-PV neurons at 10, 20, or 40 Hz, respectively. Gray dashed lines: ±4 Hz the light stimulation frequency, i.e., the frequency bands analyzed in (I) and (K).(K) Comparison of the relative LFP power ratio (band: ±4 Hz the light stimulation frequency) between 10, 20, and 40 Hz activation of mPFC FS-PV neurons; 40 Hz activation results in significantly more LFP resonance than activation at 10 or 20 HzScale bars: 20 μm (B, E), 500 μm (D). Cas9, CRISPR-associated protein 9; sgRNA, single-guide RNA; DSB, double-strand break; HA-L/HA-R, left and right homology arm, respectively; P2A, self-cleaving peptide; mPFC, medial prefrontal cortex; Cb, cerebellum; Cg, cingulate cortex area; CP, caudate putamen (striatum); dCA1, dorsal CA1; GP, globus pallidus; V1, primary visual cortex.Mean ± SEM (C, I–K); ∗p < 0.05, ∗∗p < 0.01, ∗∗∗p < 0.001. In (I, K) we adjusted the p values using the Bonferroni correction procedure.See also Figures S1 and S2.
PV neurons in the PFC of adult rats, mice, and monkeys display a typical FS phenotype with narrow APs and minor or no adaptation.26^,^27 To characterize the intrinsic electrophysiological properties of the genetically labeled PV neurons in PV-Cre rats, an AAV with Cre-dependent expression of ChR2-mCherry was injected into the mPFC (N = 8 PV-Cre rats), and whole-cell patch-clamp electrophysiology of labeled neurons (n = 37) was conducted ex vivo (Figure S1D). In line with previous reports, rheobase current injections elicited narrow APs (0.25 ± 0.05 ms), with a large and short afterhyperpolarization typical of inhibitory FS interneurons (Figures S1E and S1F). The input resistance (92 ± 24 MΩ; Figure S1F) was low, similar to the reported values for cortical FS neurons in mice.27 An AP train evoked at twice the rheobase current revealed typical FS characteristics, with a high AP firing rate (FR; 161 ± 51 Hz) and limited adaptation (102 ± 15%) in response to a 1 s (s) current pulse (Figures S1E and S1G), which is in accordance with earlier studies in both mice and rats.26^,^27 All other intrinsic electrophysiological properties confirmed the FS nature of the recorded neurons, including a short membrane time constant and a fast AP downstroke (Figure S1H). In addition, neurons labeled with biocytin (n = 6) displayed morphologies typical of cortical PV-expressing basket cells, with multipolar neurites28 (Figure S1I). No significant laminar differences were observed across the recorded population (Figures S1D and S1F–H). Based on this characterization, we in the current study refer to the inhibitory mPFC interneurons expressing PV as FS-PV neurons.
Optotagging of FS-PV neurons
To characterize mPFC neuronal and network activities associated with spatial processing, we paired chronic extracellular recordings with optotagging of FS-PV neurons29^,^30 in a cohort of adult PV-Cre rats (N = 10) engaging in a self-paced reward-seeking task.29^,^31 To render mPFC FS-PV neurons light-sensitive, an AAV with Cre-dependent expression of ChR2-mCherry was injected (Figures 1D and 1E). Behavioral training (STAR methods) started one week after viral injection. After the rats had properly learned the task (>50 trials in 1 h for three consecutive days), a microdrive holding four movable tetrodes and an optical fiber was implanted (Figures S2A and S2B). Although all rats could perform the task after the implantation, detailed post-hoc analysis of the behavior warranted exclusion of three rats, yielding a final dataset with extracellular recordings from 97 behavioral sessions (N = 7 PV-Cre rats; Figures S2C and S2D; STAR methods). Optotagging of FS-PV neurons was conducted directly before a subset of the sessions (n = 35). The optotagging and all other optogenetic experiments employed 473 nm blue light (5–10 mW), 1–3 ms light pulses for 1 s, and 5–10 light applications. A total of 349 units were recorded, and we used a Gaussian mixture model (GMM) to cluster the recorded units into WS or NS units, based on the spike width and peak-to-valley ratio32 (Figure S2E). This yielded 281 WS, 54 NS, and 14 unclassified units, respectively, corresponding to ∼80% WS, putative pyramidal neurons, and ∼15% NS, putative inhibitory interneurons (Figure 1F), which is in agreement with earlier studies of the rat PFC.33 13 NS units responded with AP firing in direct response to light application (Figure 1G), identifying them as FS-PV neurons. Using stimulus-associated spike latency test (SALT) in combination with a spike-shape correlation measure,29 we confirmed that the 13 optotagged units were directly light-driven (Figures S2F and S2G). The FS-PV neurons displayed a narrow spike width, low peak-to-valley ratio, and high FR (Figure 1F), paralleling defining features of cortical inhibitory FS-PV interneurons in mice,30 and as in mice, light activation of the FS-PV neurons resulted in inhibition of concurrently recorded WS units in the local network (Figure S2H). Some NS neurons displayed waveforms like the optotagged FS-PV neurons, presumably reflecting non-tagged FS-PV neurons (Figure S2I). A substantial fraction of the NS (non FS-PV) population should be constituted by somatostatin-expressing interneurons.34
FS-PV activation in the rat mPFC generates gamma oscillations
We previously showed that activation of cortical FS-PV neurons in the mouse barrel cortex selectively amplifies oscillations in the gamma band.35 To investigate how activation of FS-PV neurons in the rat mPFC resonates in the LFP, we in a subset of the optotagging sessions (n = 22, N = 3 PV-Cre rats) activated the FS-PV neurons with 10, 20, or 40 Hz blue light, respectively (Figure 1H). All three stimulations induced LFP resonance around (±4 Hz) the respective stimulation frequency (Figures 1I and 1J) and also harmonics occurring in integer multiplications of the stimulated frequency (Figures S2J and S2K). To compare the LFP resonance evoked by the different stimulation frequencies, we computed the LFP power ratio (Power_light_/Power_baseline_ at the stimulation frequency ±4 Hz) and found that 40 Hz stimulation of FS-PV neurons resulted in significantly more LFP resonance than the activation at lower frequencies, consistent with specific resonance in the gamma range at the network level35^,^36^,^37 (Figure 1K).
Encoding of spatial, movement, and task variables in the rat PrL
The behavioral setup used for the self-paced reward-seeking task consisted of two platforms connected by a linear track; one platform held a reward spout (reward platform), and for the reward to be delivered, the rat needed to visit the other platform (the trigger platform; Figure 2A). The task design provides the opportunity to study single trials holding two track traversals where the context and goal of the traversals differ—traversals “to reward” have reward consumption as the goal, whereas traversals “to trigger” have the goal to trigger reward availability. To quantify neuronal encoding, we used generalized linear models (GLMs) to fit the individual neuron’s activity with behavioral and task variables as regressors38^,^39^,^40 (Figure 2B). Both categorical variables (linear position [in one of 10 spatial bins; Figure 2A], run phase [to reward, or to trigger; Figure S3A], trial outcome, trial time, trial type, trial history), and continuous variables (speed, acceleration, turning) were included (Video S1; STAR methods). The performance of the models, reflecting neuronal encoding, was quantified by calculation of the explained variance (cvR^2^; squared Pearson’s correlation [r^2^] between the predicted and actual test data), obtained by 5-fold cross-validation. As a first step, we identified which variables were encoded by the PrL neurons and therefore should be included in the models. To this end, we fitted the activity of the individual neurons using models that incorporated only a single variable, and we thereafter ranked the variables based on their ability to predict the neuronal activity (Figure S3B). The variables were subsequently added one by one, in descending order, until all variables were included in the models. This revealed a lack of significant encoding of trial time, trial history, and trial type, as the inclusion of these variables did not significantly improve model performance—these variables were therefore not included in the final (full) models (Figure S3C). Across all recorded neurons, the animals’ linear position was the variable best predicting neuronal activity (2.5 ± 0.50%), followed by the run phase (2.0 ± 0.37%) and the running speed (1.6 ± 0.26%; Figure S3B). On average, the full models encompassing six variables explained 7.0 ± 1.1% (95% CI) of the variance (Figure S3C), comparable to previous reports.40^,^41Figure 2GLMs reveal conjunct encoding of behavioral and task variables in single PrL neurons(A) Schematic of the self-paced reward seeking task. Top: outline of a single trial, with indication of the traversal direction (arrow heads) and timestamps (colored dots). Red dots: beam break detectors; black dots: timestamps extracted from video. Bottom: illustration of the track with definition of the 10 spatial locations (bins) and indication of the traversal directions. In each trial, the rat must visit the trigger zone for reward to be delivered in the reward zone.(B) Schematic of the GLM approach used to quantify the individual neurons’ encoding of behavioral and task variables. Top: activity of an example neuron (black) and the activity predicted by the full model (red). Bottom: scoring of the behavioral and task variables included in the full model.(C–E) Quantification of the correlation between neuronal activity and behavioral and task variables.(C) Encoding of model variables by individual neurons was quantified using two approaches. Top: average explained variance (cvR^2^, 5-fold cross-validated) of single variable models. Bottom: average unique explained variance (Δr^2^) of the variables.(D) The fractions of neurons with significant encoding (p < 0.01, permutation test) of the different behavioral and task variables. Dashed line: mean across all variables (47 ± 4.4%).(E) Number of variables significantly encoded by each neuron. Many neurons encode more than one variable.(F) Pearson’s correlation between the unique explained variance (Δr^2^) for all combinations of variables. There is a significant correlation between the animal’s position on the track (linear position) and the direction of the track traversal (run phase; p < 0.05, Holm-Bonferroni correction as compared with shuffled data).(G) Scatterplot of the unique explained variance (Δr^2^) for the linear position (x axis) vs. the run phase (y axis) for all individual neurons (n = 322), revealing a significant correlation.Mean ± SEM (C); ∗p < 0.05.See also Figure S3.
Video S1. GLM approach, related to Figure 2. Movie (35 s, 3 trials) of animal behavior with scoring of task and behavioral variables used in the full GLM and activity from an FS-PV neuron (Black lines: Spike times, red line: firing rate [Hz]). The full GLM consisted of the categorical variables linear position, run phase, and trial outcome and the continuous variables speed, acceleration, and turning. Orange dots: tracking of the animal’s ears and tail base generated from DeepLabCut.
We observed that the explained variance of the full models was lower than predicted, i.e., lower than the sum of the explained variance of the individual variables (Figure S3C). This indicates an overlap in the predictive power of some variables, i.e., certain variables contained redundant information. This suggests that cvR^2^ is a suboptimal measure of variable encoding. To optimally capture each variable’s unique contribution (ΔR^2^) to the models, we created reduced models, by removing single variables and quantifying the reduction in cvR^2^ relative to the full models38^,^39^,^40 (Figure S3D). As expected, the removal of any of the variables reduced the explained variance (cvR^2^) of the full models, with linear position, the run phase, and the running speed, causing the largest reductions (Figures S3D and S3E). Overall, the animal’s linear position was the best predictor of the activity of the PrL neurons (Figures S3B and S3E).
Cell-type-specific encoding of behavioral and task variables in the rat PrL
Our observations of spatial and movement encoding in the PrL corroborate previous findings in mice5^,^40 and rats.42 However, to the best of our knowledge, the activities of FS-PV neurons have not been accounted for. We therefore as the next step separately characterized the encoding by the FS-PV, WS, and NS populations. Both the explained variance (cvR^2^) of the single variable models and the unique contribution (ΔR^2^) derived from the reduced models indicated a lack of significant differences in the encoding strength of the individual variables between the three neuronal populations (p > 0.05; t test, Figure 2C). Comparable fractions of WS, NS, and FS-PV neurons encoded the individual variables, and overall, the behavioral and task variables were significantly encoded by 47 ± 4.4% of the PrL neurons (Figure 2D). A minority of the WS, and the NS, neurons did not encode any of the variables (WS: 28/271; 10.3%, NS: 7/38; 18.4%; Figure 2E). The majority of the neurons (238/322; 73.9%) encoded more than one variable, indicative of mixed selectivity.43^,^44 To investigate this further, we computed the pairwise correlation between the encoding strength of all variables, which revealed a significant linear relationship between the unique contribution (ΔR^2^) of the animal’s linear position on the track and the run phase (p < 0.05 permutation test, Holm-Bonferroni correction; Figures 2F and 2G). This indicates that subpopulations of PrL neurons encode the rat’s spatial location on the track and the traversal direction (to vs. away from the reward), i.e., PrL neurons display context-dependent spatial representations (see further below).
Context-dependent spatial encoding in the rat PrL
The GLM showed that neurons in the rat PrL encode the spatial context, and to reveal the associated neuronal activities, we analyzed the firing patterns in relation to both the animal’s spatial location and the direction of the track traversal. The WS neurons overall fired sequentially along the 20 bins of the track (10 bins to trigger + 10 bins to reward), i.e., individual neurons fired in a few consecutive bins in only one of the track directions (Figures 3A, 3B, and S4A). For the FS-PV neurons, the spatial context was reflected by elevated FR in traversals to the trigger zone (Figures 3A–3C and S4A). The FS-PV activities differed particularly between the two platforms, with >60% of the FS-PV neurons displaying significantly increased activity in the trigger zone (p < 0.05 permutation test; STAR methods) and no FS-PV neurons showing increased activity in the reward zone (Figure 3D). Analysis of the correlation of the FRs between the two traversal directions confirmed the presence of traversal-specific activity patterns and dissimilar FS-PV activities particularly in the end of the traversals (Figure 3E). The traversal-specific firing patterns were consistent (spatial consistency) between odd and even trials, refuting that the patterns reflect random fluctuations in the activity5^,^40 (Figures S4B and S4C). Across the recorded population, 63.0% (n = 201/319) of the neurons displayed significantly different mean activities in traversals to reward vs. traversals to trigger (p < 0.05, t test; WS: 64.2%, n = 172/268, NS: 55.3%, n = 21/38, FS-PV: 61.5%, n = 8/13; Figure 3F). On the population level, the activity in the two traversals was significantly different only for the FS-PV population (Figure 3F). To ensure that the finding of traversal-specific activity patterns was not attributed to the averaging of activities across many traversals, we used SVMs to decode the direction of individual traversals from the neuronal activity (STAR methods). We found that the direction of individual traversals could be predicted with high and significant accuracy based on the neuronal activity from either of the three cell types (WS: 66 ± 1.5%, NS: 64 ± 3.7%, PV: 69 ± 6.0%; Figure 3G).Figure 3. Encoding of spatial context(A) The trial-averaged FR (z-scored) of all individual neurons, split by cell type and traversal direction. The neurons are peak sorted by the FR in traversals to reward and plotted in the same order for both traversal directions.(B) The mean activity (z-scored) of WS neurons significantly modulated by the animal’s traversal direction (p < 0.05, Wilcoxon rank-sum test). Different WS populations are activated in traversals to reward vs. to trigger. The two populations show opposing activity patterns during the respective traversals.(C) The mean activity (z-scored) of the three cell types in the two traversal directions. The FS-PV population displays higher activity in traversals to the trigger than in traversal to the reward, with particularly dissimilar activities in the end of the traversals (i.e., when the animals reach the respective platform).(D) The fraction of neurons with significantly modulated activity (p < 0.05, permutation test) at each of the 10 track positions (bins) in the two track traversals. Orange: increased activity in traversals to reward; purple: increased activity in traversals to trigger. Gray dashed lines: 50%.(E) Comparison of the FR in the two traversal directions. The correlation (r) between the FR in traversals to reward vs. to trigger was computed for each neuron [numbers as in (C)]. The FR of all cell types was most similar in the first half of the traversals, and least similar at the end of the traversals, and this was particularly true for the FS-PV neurons. First 50%: WS: 0.23 ± 0.002, NS: 0.34 ± 0.017, FS-PV: 0.48 ± 0.043; Full run: WS: 0.17 ± 0.002, NS: 0.29 ± 0.013, FS-PV: 0.21 ± 0.046), Last 50%: WS: 0.05 ± 0.003, NS: 0.15 ± 0.017, PV: −0.09 ± 0.059; Full run reversed: WS: 0.0 ± 0.002, NS: −0.1 ± 0.015 and PV: 0.12 ± 0.047. Gray area: 95% confidence interval of shuffled data.(F) The modulation index (MI) of the traversal direction for the three cell types. Positive values: increased activity in traversals to reward, negative values: increased activity in traversals to trigger. Mean MI (black line): WS, n = 268: 0.008 ± 0.32, p = 0.68; NS, n = 38: 0.027 ± 0.309, p = 0.61; FS-PV, n = 13: −0.068 ± 0.086, p = 0.018; t test. The FS-PV population has significantly higher activity in traversals to trigger than in traversals to reward. Black circles; significantly modulated neurons (p < 0.05, Wilcoxon rank-sum test; to trigger: WS: n = 93, NS: n = 13, FS-PV: n = 6; to reward: WS: n = 79, NS: n = 8, FS-PV: n = 2).(G) For all three neuron types, SVMs could predict the animals’ traversals direction above the chance level, from the activity of individual neurons when the SVMs were trained on each neuron's individual activity (WS: p = 6.0 × 10^−58^, NS: p = 7.5 × 10^−9^, PV: p = 2.8 × 10^−5^, vs. control data with shuffled traversal direction labels (light gray bars), paired t test). This indicates that all three neuronal cell types encode the traversal direction. One dot = one neuron, numbers as in (F).(H) Left: SVMs could predict the animals’ traversal direction above the chance level from the activity of individual FS-PV neurons when the SVMs were trained on data from 1 to 10 randomly selected, different, FS-PV neurons. Black traces: predictions from SVMs trained on control data with shuffled traversal direction labels. Right: the mean decoding accuracy for all individual neurons (SVM training as in left), demonstrating significantly increased decoding accuracy for only the FS-PV population (FS-PV: p = 0.004 WS: p = 0.172, NS: p = 0.705, vs. control data with shuffled traversal direction labels; paired t test). This suggests more homogeneous firing patterns across the FS-PV population than within the WS, or the NS, population. One dot = one neuron, numbers as in (F).(I) Correlation between the decoding accuracy (G) and the unique contribution of the variables used in the full model (Figure 2C), revealing a significant correlation specifically for the linear position and the run phase*.* Gray area: 95% confidence interval of shuffled data.Mean ± SEM (B, C, E, H left); ∗p < 0.05, ∗∗p < 0.01, ∗∗∗p < 0.001.See also Figure S4.
To address how homogeneously the neurons within a cell type represented the two traversal directions, we analyzed if linear support vector machines (SVMs) trained on data from 1 to 10 randomly selected neurons of a cell type could predict the direction of individual traversals based on the activity of a single, different, neuron of the same cell type. We found that the traversal direction could be predicted above the chance level from the activity of FS-PV neurons but not from WS, or NS, neurons (Figure 3H). This indicates that the spatial context is represented by similar activity patterns across the FS-PV population, which aligns with the finding of quite uniform FS-PV activities in both traversals to trigger (high FR) and traversals to reward (low FR; Figure 3A).
The compiled conclusion of the SVM analyses is that although only the FS-PV population displays homogeneous activities, the traversal direction can yet be decoded from either of the three populations. Neurons that display high decoding accuracy for the traversal direction would be predicted to show strong encoding of the run phase (GLM, Figure 2C). To confirm this, we analyzed the correlation between the decoding accuracy for the traversal direction and the unique contribution of all variables in the full model. As expected, the decoding accuracy for the traversal direction correlated significantly with the run phase, and also with the linear position, but not with the other variables in the full model (Figure 3I). On the whole, our analyses identify neurons in the PrL that do not only encode the animals’ actual location in space (“place cells”) or a specific location relative to the start or stop of the movement trajectory (i.e., a specific “trajectory phase”)31 but rather the context of the movement in space, i.e., the spatial context.
Context encoding in relation to movement variables
Signals related to movement are considered ubiquitous and can confound the correlation of neuronal activity to cognitive and other task-related processes.45^,^46 As we observed that PrL neurons encode movement variables (Figure 2), we therefore next moved our attention to the animals’ movement on the track (speed, acceleration, and turning) and the encoding of these continuous variables. We found the whole continuum of movement speeds (0–100 cm/s) to be reflected in the activity of the WS population, with some WS neurons being tuned to a narrower range of speeds than others (Figure 4A). In contrast, the WS activities associated with acceleration were strikingly dichotomized, with absolutely most WS neurons being tuned to either acceleration or deceleration (Figure 4A). Also, the WS responses to ipsi- vs. contralateral turning on the platforms were dichotomized (Figure 4A). Importantly, the animals’ traversals to reward were faster and had a different acceleration pattern than traversals to trigger (Figure 4B), and it was therefore a possibility that the neuronal activities interpreted as encoding of spatial context (Figure 3) would be better explained by variables related to the animals’ movement on the track. To address this, we used a GLM to quantify the encoding strength of speed, acceleration, and spatial context, respectively, during the animals’ running on the track (Figure S3A; STAR methods). Encoding of spatial context was evaluated by generation of a new variable, position with direction, which captures both the animal’s position and direction on the track. Reduced models were used for quantification of the unique explained variance of the speed, acceleration, and position with direction variables (Figure S3STAR methods). This identified that the position with direction variable explained significantly more of the variance than both the speed and acceleration variables only for the FS-PV population (Figure 4C), i.e., the spatial context was encoded significantly more strongly than the movement variables by this specific cell type. Although position with direction was the strongest predictor also for the WS activity, this variable did not explain significantly more of the variance than the speed variable. In essence, although spatial context is encoded by the WS population, some activity differences in the two track traversals are attributed to speed encoding. In support of this, similar fractions of WS neurons encoded speed and position with direction, respectively (Figure 4D). Overall the analyses show that although spatial context is encoded by all three PrL cell types, this encoding is particularly pronounced in the FS-PV population.Figure 4. Encoding of spatial context in relation to movement variables(A) Top: For each cell type, the mean firing rate (z-scored) as a function of speed (cm/s, bin size = 10 cm/s; left), acceleration (cm/s^2^, bin size = 1 cm/s^2^; middle), and turning (degrees/s—head angular velocity ipsilaterally [positive values] or contralaterally [negative values] to the recorded hemisphere, bin size = 10°/s; right). Bottom: the trial-averaged FR (z-scored) for all individual neurons. The neurons are peak sorted in the individual panels.(B) Mean speed (left) and mean acceleration (right) in traversals to reward and trigger, from all sessions (n = 97). Black horizontal lines: significant difference between the two traversal directions (p < 0.05, paired t test).(C) The unique explained variance (Δr^2^) of the variables speed, acceleration, and position with direction, respectively. Only the activity of the FS-PV neurons is significantly better explained by position with direction than by speed or acceleration.(D) The fraction of neurons with significant encoding (p < 0.01, permutation test) of the variables speed, acceleration, and position with direction, respectively. A significantly larger proportion of the FS-PV neurons encode the variable position with direction than the variables speed and acceleration*,* respectively (p < 0.01, binomial test).Mean ± SEM (A–C); ∗p < 0.05, ∗∗∗p < 0.001; paired t test (B, C). Start/end = bin 1 or 10 depending on the traversal direction (B; see Figure 2A).
Prefrontal gamma oscillations in relation to spatial processing
LFP oscillations reflect network activity patterns and are ubiquitous signatures of operating circuits. In line with this, oscillations are used for readout of computations and behavioral variables, in the PFC often with a focus on gamma oscillations due to the strong correlation between prefrontal gamma activity and cognition.30^,^47^,^48 Building on these concepts, we next investigated the modulation of the LFP activity during the animals’ running on the track. Spectral analysis of the LFP oscillations revealed differences between the traversal directions, with the power in the gamma band being significantly higher in traversals to reward than to trigger (Figures 5A–5C, S5A, and S5B). Previous studies have reported that gamma power increases occur in brief bursts and that findings of sustained, continuous gamma power increases are a result of averaging, e.g., over trials.49^,^50^,^51 In agreement with this, single-trial analysis demonstrated a pattern of bursts of increased 30–50 Hz gamma power rather than a continuous elevation (Figures 5D and S5D), with the rate of gamma bursts being significantly higher in traversals to reward than in traversals to trigger (paired t test, p < 0.001; Figure 5E). In the hippocampus, the expression of gamma oscillations has been correlated to the speed of movement,17^,^52 and as the speed differed in the two traversal directions (Figures 4B, 5F, and S5E), we investigated the relationship between speed and 30–50 Hz gamma power. We found an overall positive correlation, but importantly, accounting for the differential running speeds in the two directions did not abolish the difference in 30–50 Hz gamma power between the two traversal directions (Figures 5G, S5F, and S5G). Thus, the prefrontal computations involved in the asymmetrical reward-seeking task were reflected by modulation of the 30–50 Hz gamma band activity.Figure 5. Modulation of PrL gamma oscillations by reward-seeking behavior(A–G) Analysis of the modulation of the LFP activity during the animals’ running on the track, i.e., bins 2–9 or 9–2 (see Figure 2A). Only temporally homogeneous track traversals (0.5–1.5 s) are included in the analysis.(A–C) Traversals to reward are characterized by increased gamma oscillations.(A) The relative LFP power (1–100 Hz) in traversals to trigger vs. to reward.(B) The difference in relative power in traversals to trigger vs. to reward. Gray bars: frequency bands with statistics in (C).(C) The difference in relative power in three frequency bands, between traversals to trigger and traversals to reward. Positive values: increased relative power in traversals to reward; negative values: increased relative power in traversals to trigger. Black lines: mean, gray circles: individual recording sites.(D and E) Traversals to reward are signified by an increased rate of 30–50 Hz gamma bursts.(D) Example LFP traces (0.8 s) from a single traversal to reward. From the top: raw LFP; band-pass filtered LFP (30–50 Hz); spectrogram (10–100 Hz); extraction of the mean power in the 30–50 Hz gamma band.(E) Detection of individual gamma bouts during traversals to reward (y axis) and to trigger (x axis), respectively. Bursts/s: to reward: 5.9 ± 0.1, to trigger: 4.8 ± 0.1; p < 0.001, paired t test.(F and G) The relationship between running speed and power in the LFP spectrum.(F) The running speed and its dynamics differ depending on the direction of the track traversal. The mean speed across the recording sessions (n = 47) is shown; for the speed in individual sessions see Figure S5E.(G) The LFP power in two gamma frequency bands after adjustment for the differential running behavior in the two track traversals. Top: while the 30–50 Hz gamma power overall increases with the running speed, the power is higher in traversals to reward than in traversals to trigger. Bottom: the power in the 60–80 Hz band in contrast decreases with the running speed and does not differ depending on the direction of the track traversal. The analysis was restricted to speed bins encompassed by the speed distribution of both traversal directions (15–75 cm/s, bin size 1 cm/s; Figure S5E), and sessions holding any of the 15–75 cm/s speed bin were included..Forty-seven sessions (A–F); 91 LFP recording sites (A–C, E); LFP recording sites: to trigger n = 87 ± 4, to reward n = 88 ± 6 [mean ± SD; (G)]; one dot = the mean at one recording site (C, E); Start/end = bin 2 or 9, depending on the traversal direction (see Figure 2A); mean ± SEM (A, B, E-G); ∗∗∗p < 0.001..See also Figure S5.
Spatial encoding in the rat hippocampus is signified by neuronal spiking coupled to the phase of ongoing network oscillations in the gamma and theta bands.53^,^54 However, the relationship between local network oscillations and neuronal discharge in prefrontal operations remains to be deciphered, including during spatial processing, and we therefore next investigated the strength of spike phase-coupling during the track traversals. We focused on the gamma band (30–50 Hz) and quantified spike phase-coupling using two different measures: pairwise phase consistency (PPC55) and mean resultant length (MRL), respectively. At the population level, both measures identified increased spike-gamma phase-coupling during traversals to reward for all three PrL cell types, although only the MRL showed statistical differences between the two traversal directions (Figures 6A and 6B). As both the rate of gamma bursts and the spike-gamma phase-coupling were increased in one of the traversal directions (to reward), we wondered if the spatial representation in the PrL neurons also differed between the traversals, i.e., if the animals’ position in space (linear position on the track) was more prominently represented in one of the traversal directions (or not). To quantify the spatially related activity, we computed the spatial information (SI)5 from spatially binned firing rates (STAR methods). This identified significantly higher SI in traversals to reward than in traversals to trigger for the WS neurons (Figure 6C). We next investigated a possible relationship between the gamma phase-coupling and the spatial representation in PrL neurons. We identified four subpopulations of neurons—neurons with significant gamma phase-coupling in (A) traversals to reward, (B) traversals to trigger, (C) both traversal directions, and (D) neurons lacking significant gamma phase-coupling. The low number of FS-PV and NS neurons, respectively, precluded proper analysis of subpopulations within these cell types, and we therefore focused the further analyses on the large WS population (Figure 6D). None of the WS subpopulations displayed significantly different FR in the two traversals (Figure 6E). The increased SI in traversals to reward (Figure 6C) persisted across the four WS subpopulations, i.e., the SI was highest in traversals to reward regardless of the gamma phase-coupling properties of the WS neurons (Figure 6F). This strongly indicates that spike gamma phase-coupling is not an overarching feature of spatial processing in the rat mPFC. In support of this, the levels of SI in traversals to reward and traversals to trigger, respectively, were similar across the four WS populations (Figure 6F). Furthermore, although the WS population with spike-gamma phase-coupling in both traversal directions showed a positive correlation between the PPC and the SI, this correlation was weak and non-significant (Figures 6G and 6H). Two informative observations can be made from the data. First, the spatial information in the firing of the WS neurons was consistently increased when the animals moved toward an expected reward, regardless of the relationship between the spiking and the ongoing gamma cycles (Figure 6F). Second, subpopulations of WS neurons displayed gamma phase-coupling selectively in one of the traversal directions, although the neurons’ SI was high in both directions. Both observations identify that the spatial representation in the WS neurons is not related to their gamma phase-coupled firing. This further suggests that although gamma phase-coupling is not directly related to spatial encoding in the mPFC, gamma phase-coupling is a feature of discrete WS ensembles during goal-directed reward-seeking behavior. It remains to be established what operations the gamma phase-coupling support and what computations the modulations of the gamma oscillations reflect.Figure 6. Spike-gamma phase-coupling of PrL WS neurons does not reflect spatial processing(A) No significant differences in the strength of the spike-gamma phase-coupling (quantified as pairwise phase consistency, PPC) were found between the two traversal directions (WS: p = 0.720, NS: p = 0.061, FS-PV: p = 0.119).(B) The strength of spike-gamma phase-coupling in the two traversal directions, quantified as MRL. This measure identifies that the spiking of neurons of all three cell types is significantly more phase-coupled to local gamma oscillations (30–50 Hz) in traversals to reward than in traversals to trigger (WS: p = 0.01, NS: p = 0.003, FS-PV: p = 0.04). Numbers as in (A).(C) The spatial information (SI) in WS neurons was significantly higher in traversals to reward than in traversals to trigger. WS: p = 1.9 × 10^−5,^ NS: p = 0.671, FS-PV: p = 0.115, paired t test. Numbers as in (A).(D–H) Lack of support for gamma phase-coupling of WS neurons being a feature of spatial encoding in the PrL.(D) WS neurons were divided into four groups, based on their spike-gamma phase-coupling properties.(E) None of the four populations of WS neurons displayed significantly different FR in the two traversals.(F) Regardless of the WS neurons’ spike-gamma phase-coupling properties, the SI was highest in traversals to reward (difference in SI between traversals to reward and to trigger: coupled to reward: p = 0.42, coupled to trigger: p = 0.004, coupled to both: p = 0.04, not phase-coupled: p = 0.001). No significant difference in SI was found between the WS subtypes in traversals to reward (p = 0.75) or to trigger (p = 0.52), respectively (ANOVA).(G and H) Analysis of the relationship (Pearson’s correlation [r]) between the spike-gamma phase-coupling (PPC) and the spatial information (SI) in the two traversal directions.(G) Bar plots of the correlation between PPC and SI in the two traversal directions.(H) Scatterplots of the PPC (x axes) vs. the SI (y axes) in the two traversal directions for all individual neurons in the four WS populations.Mean ± SEM (A–F); ∗p < 0.05, ∗∗p < 0.01, ∗∗∗p < 0.001; paired t test (A–F).
Discussion
We here introduce a knock-in rat line with faithful and specific Cre expression in PV neurons and demonstrate its utility for cell-type-specific experiments ex vivo, and in vivo during behavior, respectively. Our ex vivo experiments demonstrate that mPFC PV neurons in adult rats display properties typical of inhibitory FS interneurons and in many ways mirror cortical FS-PV neurons in adult mice. The optogenetic experiments demonstrated direct light activation of mPFC FS-PV neurons and concurrent silencing of WS, putative pyramidal, neurons in the local circuit, confirming the cell identity of the Cre-expressing neurons.
The theoretical and experimental support for the importance of FS-PV activities in the generation of gamma oscillations is ample, and optogenetics has been frequently used in mouse studies to address how gamma rhythmicity relates to information processing and cognition in both health and disease.35^,^36^,^37^,^48 Optogenetic drive of FS-PV neurons in the rat mPFC resulted in specific resonance in the gamma range at the network level, replicating findings in the adult mouse cortex.35^,^36^,^56 Optogenetic manipulation of PV neurons in different brain regions has in the past 15 years found utilities beyond studies of the neurons per se—photostimulation of cortical FS-PV neurons has proven a powerful means for rapid inactivation of cortical microcircuits in mice,57^,^58 and gamma frequency activation of FS-PV neurons in cortex or hippocampus has been used for both restoration and improvement of cognition and/or behavior.30^,^48^,^59 Our data suggest that the PV-Cre rats open up for new research approaches also in this species.
Encoding of spatial context by mPFC FS-PV neurons
mPFC neurons tend to be less spatially selective than hippocampal place cells,5 and the spatial processing is intricately tied to a variety of factors beyond mere spatial location, including goal locations, past choices, the surrounding environment’s context, and task structures.7^,^8^,^9^,^10 Thus, the mPFC appears to emphasize a location in relation to cognitive and behavioral variables relevant to the task at hand. Our study adds to this body of work and demonstrates that although different cell types in the rat PrL encode discrete variables reflecting the animal’s spatial location, movement (speed, acceleration, turning), and if the behavior was rewarded or not (trial outcome), subpopulations of single neurons encode the animal’s spatial position in relation to a rewarded goal. Although we identified the encoding strength of discrete behavioral variables to be similar across the three cell types studied (Figure 2), the spatial context was particularly encoded by the FS-PV population (Figure 4). A recent study identified that neurons in the mouse anterior cingulate cortex are tuned to a specific trajectory phase, i.e., a spatial location relative to the start and end of a traversal on a linear track, regardless of the animal’s direction on the track.31 Importantly, in this study, the animals could collect a reward on both ends of the track, whereas in our study reward was only available in one of the platforms. Together the studies suggest that the asymmetric presence of a reward site is what drives the differential spatial encoding of the two track traversals in our study. Supporting this, recent studies using a spatial working memory task have shown that prefrontal pyramidal firing is more influenced by the animal’s direction (inbound vs. outbound in relation to reward) than by the task phase (sample vs. choice).7^,^40
We found that the WS held more SI during traversals to reward than during traversals to trigger, suggesting stronger spatial representation when the rats ran toward the reward. In line with this, the spatial information in the spiking of putative pyramidal neurons in the PFC has been found to be higher when mice approach a reward site compared with when moving away from it, a difference that was abolished when the reward was removed.7 We show here that the traversals to the reward site are associated with increased power and bursts of gamma oscillations in the PrL, i.e., the asymmetric reward seeking is associated with modulation of the LFP activity in the gamma band. As gamma rhythms are suggested to temporally organize spiking, we enquired about the relationship between the WS and the gamma cycle phases and if spike-gamma phase-coupling was a feature of spatial processing in the PrL. Our results do not support that local gamma oscillations organize spiking activities part of spatial encoding in the PrL and instead suggest that the strong modulation of the gamma band activity is related to other prefrontal operations part of reward-seeking behavior. Future work could employ a longer linear track for ambulating traversals, optimized for investigation of spike-phase relationships to lower LFP frequencies, including theta frequencies. The generated PV-Cre line presents opportunities for detailed enquiries in rats about the role of inhibitory PV neurons in population activities and their relationship to theta and gamma rhythms, across the cortex and hippocampal formation.
Limitations of the study
By use of optotagging in PV-Cre knock-in rats, we demonstrate that PrL FS-PV neurons encode the spatial context, which is supported by a range of statistical tests. Recording the activity of a larger number of FS-PV neurons could allow the study of FS-PV neuronal subpopulations, divided by features such as gamma phase-coupling or cross-correlation characteristics. In addition, our analysis of neuronal activity relied on linear methods such as GLMs and SVMs that do not capture nonlinear relationships between behavioral variables and neuronal activity.
STAR★Methods
Key resources table
REAGENT or RESOURCESOURCEIDENTIFIERAntibodiesAnti-Cre (rabbit)Synaptic systemsCat#257 003; RRID :AB_2619968Anti-PV (Guinea pig)SwantCat#GP72; RRID: AB_2665495Anti-PV (Guinea pig)Synaptic systemsCat#195 004; RRID: AB_2156476Anti-RFP (rabbit)RocklandCat# 600-401-379; RRID: AB_2209751Anti-Rabbit (donkey, 488)JacksonCat# 711-545-152; RRID: AB_2313584Anti-Guinea pig (donkey, Cy3)JacksonCat# 706-165-148; RRID: AB_2340460Anti-Guinea pig (donkey, Cy5)JacksonCat# 706-175-148; RRID: AB_2340462Bacterial and virus strainsAAV5-Ef1a-DIO-hChR2(H134R)-mCherryUNC Gene Therapy CenterLot #AV4314BChemicals, peptides and recombinant proteinsDakoTarget Retrieval SolutionDakoS2367Experimental Models: Organisms/StrainsPvalb-P2A-CreSAGE LabsN/ASoftware and algorithmsCheetah acquisition softwareNeuralynxN/AMATLAB 2019aMathworksN/APython 3.8Python Software FoundationN/AOtherDMCdriveKim et al.62N/AflexDriveVoigts et al.61N/AOptic fiberThorlabsFP200ERT
Resource availability
Lead contact
Further information and requests for resources and reagents should be directed to and will be fulfilled by the lead contact, Marie Carlén ([email protected]).
Materials availability
The PV-Cre rat generated in this study is available upon reasonable request to the lead contact.
Data and code availability
- •The data reported in this paper will be shared by the lead contact upon request.
- •This paper does not report original code. All custom MATLAB and python code will be available upon request.
- •Any additional information required to reanalyze the data reported in this paper is available from the lead contact upon request.
Experimental model and study participants details
A total of 24 adult (2–6 months) female and male heterozygous PV-Cre rats were used. 3 PV-Cre rats were used to characterize the expression overlap of PV and Cre. 3 PV-Cre rats were used to characterize viral recombination. 8 PV-Cre rats were used for ex vivo electrophysiological experiments. 10 PV-Cre rats were used for in vivo electrophysiological and behavioral experiments. Animals were maintained on a reversed 12-h light/dark cycle and all experiments were performed during the dark phase. For animals in behavior, the daily amount of food was restricted to 10 g. The animals were weighed daily, and the weight did not subside 85% of the initial free feeding weight. Animals were single housed after microdrive implantation. All procedures were approved and performed in accordance and compliance with the guidelines of the Stockholm Municipal Committee.
Method details
Anatomy
All anatomical measurements and nomenclature are based on Paxinos G and Watson C, The rat brain in stereotaxic coordinates, 6^th^ edition. In the current manuscript, mPFC refers to PrL + IL, Cg to Cg1 + Cg2. dCA1 denotes dorsal CA1 and was determined visually by imaging followed by microscopy.
Generation of the PV-Cre rat line
The PV-Cre rat line (‘Pvalb-P2A-Cre rat model’) was produced by SAGE Labs (Saint Louis, MO, USA) using CRISPR-based technology. Cre was introduced into the 3′ UTR of the Parvalbumin (Pvalb) locus using a 2A self-cleaving peptide sequence between Pvalb and Cre. This strategy was chosen to allow the expression of Cre without disruption of the expression of Pvalb.
Virus injections
Burprenorphine was delivered (I.P 0.05 mg/kg) 30 min before surgery. The animals were deeply anesthetized with isoflurane (1.5–2.5%), and the body temperature maintained at 37°C using a heating pad. The animals were fixed in a stereotaxic frame (Harvard Apparatus, Holliston, MA) and a small craniotomy (0.5 mm) was made unilaterally over the mPFC (AP 3.0–3.3 mm, ML 0.5–0.8 mm). The virus (AAV5-Ef1a-DIO-hChR2(H134R)-mCherry, UNC Gene Therapy Center, #AV4314B, 4x10^12^ virus molecules/ml) was delivered by a glass capillary attached to a motorized Quintessential Stereotaxic Injector (Stoelting, Wood Dale, IL, USA) at rate of 0.05–0.1 μL min^−1^. Injection in the mPFC: half the volume (0.5 μL) was injected at DV -2.0 mm. The capillary was thereafter held in place for 5 min before being slowly lowered to DV -3.0 mm, at which the rest of the virus (0.5 μL) was injected. The pipette was thereafter held in place for 10 min before being slowly retracted from the brain. The incision was closed with stitches (Ethicon, USA), and postoperative caprofren (5 mg/kg) was administered every 24 h for 48 h.
Tissue processing
General procedure
For perfusions, the animals were deeply anesthetized with pentobarbital and transcardially perfused with 1x PBS followed by 4% paraformaldehyde (PFA) in 1x PBS. The perfused brain was removed from the skull and postfixed in 4% paraformaldehyde in 1x PBS at 4°C for 16 h. The brains were thoroughly washed in 1x PBS and thereafter sectioned (50 μm thickness) using a vibratome (Leica VT1000, Leica Microsystems, Nussloch GmbH, Germany).
Immunohistochemistry
PV and Cre co-expression across brain regions and the cortical depth
Vibratome cut (free floating) tissue was heated to 70°C in 1x Daco Target Retrieval solution (DakoTarget Retrieval Solution, pH 6.1, Dako, code. S2367) for 15 min and left to cool at room temperature (RT). The sections were transferred to 1x TBST (0.3% Triton X- in 1x TBS) for 1 h, blocked with 10% normal donkey serum in 1x TBST for 1 h, and thereafter incubated with primary antibodies (1:250 Cre-recombinase, rabbit, Synaptic Systems, cat. no. 257 003, and 1:1000 Parvalbumin, guinea pig, Swant, code no. GP72, or Parvalbumin, guinea pig, Synaptic Systems, cat. no. 195 004) in 1x TBST at RT for 12–24 h. The sections were thereafter washed three times in 1x TBST and incubated with a species-specific fluorophore conjugated secondary antibody in 1x TBST for 3–5 h (1:500 donkey anti-rabbit, DyLight 488, Jackson, 711-545-152 and 1:500 donkey anti-guinea pig, Cy3 or Cy 5, Jackson, 706-165-148/706-175-148). The sections were thereafter transferred to 1x TBST with 1:50000 DAPI for 3 min, and consecutively washed with 1x TBST, 1x TBS and 1x PBS (10 min each). The sections were mounted on glass slides (Superfrost Plus, Thermo Scientific) and coverslipped (Thermo Scientific) using 50:50 Glycerol:1x PBS.
PV and ChR2-mCherry co-expression in the mPFC
Immunohistochemistry and mounting of sections were performed as described above. The same PV antibodies were used. Primary antibody for detection of ChR2-mCherry: RFP: 1:500 RFP, rabbit, Rockland code 600-401-379.
Imaging, and quantifications
For scoring of the co-expression of Cre and PV across brain regions (mPFC, Cg, V1, dCA1, CB; N = 3 PV-Cre rats, CP, GPe; N = 2 PV-Cre rats) and also for the scoring of the co-expression of Cre and PV across the cortical layers (mPFC, Cg, V1; N = 3 PV-Cre rats) tiled z stack images were acquired at 20x (3-4 sections per brain region for each rat), using a Zeiss (LSM800) confocal microscope with a motorized stage. The co-expression of Cre and PV was quantified using a custom script in R. The position of labeled neurons across cortical layers was determined using a custom MATLAB (MathWorks) script utilizing cell and brain surface coordinates manually mapped in ImageJ (Cell counter). Representative high-resolution z stack images (20–30 focal planes) of neurons co-expressing PV and Cre were acquired at 40x or 63x with the same Zeiss (LSM800) microscope.
Scoring of co-expression of PV and ChR2-mCherrry was done by microscopy of the mounted sections (3-4 sections/rat, N = 3 PV-Cre rats) using a Leica DM6000B fluorescent microscope.
Patch clamp recordings
For characterization of the neurophysiological properties of cortical Cre expressing neurons, an AAV with Cre-dependent expression (AAV5-Ef1a-DIO-hChR2(H134R)-mCherry (UNC Gene Therapy Center, #AV4314B) was targeted to the mPFC of adult (2–3 months) heterozygote PV-Cre rats (N = 8; 1 μL unilateral injection, 4x10^12^ virus molecules/ml, for details see Viral injections). 2–3 w after viral injection, 250 μm thick coronal slices were vibratome cut (VT1200S, Leica, Wetzlar, Germany) in ice-cold sucrose solution, containing (in mM): 126 NaCl, 2.5 KCl, 1.25 NaH_2_PO_4_, 24 NaHCO, 25 glucose, 75 sucrose, 1 L-ascorbic acid, 3 Na-pyruvate, 1 CaCl_2_, 4 MgCl_2_. The slices were placed in an extracellular solution, containing (in mM): 126 NaCl, 2.5 KCl, 1.25 NaH_2_PO_4_, 26 NaHCO_3_, 20 glucose, 1.5 CaCl_2_, 1.5 MgCl_2_ and incubated at 34°C for 30 min, and thereafter kept at RT. For recording, the slices were superfused with extracellular solution kept at 33–35°C. Neurons were visualized using a DIC microscope (Scientifica, Uckfield, UK) with a 60x objective (Olympus, Tokyo, Japan), and fluorescent neurons identified using a pE-4000 LED light source (CoolLED, Andover, UK). Patch pipettes (resistance 5–10 MΩ, pulled on a P-87 Flaming/Brown micropipette puller, Sutter Instruments, Novato, CA, USA) were filled with low chloride internal solution containing (in mM): 130 K-gluconate, 5 Na_2_-posphocreatine, 1.5 MgCl_2_, 10 HEPES, 5 Mg-ATP, 0.35 Na-GTP, 1 EGTA, 8 biocytin. Signals were recorded with an Axon MultiCalmp 700B amplifier and digitized at 20 kHz with an Axon Digidata 1550B digitizer (Molecular Devices, San Jose, CA, USA). Access resistance and pipette capacitance were compensated. To assess passive and active membrane properties neurons recorded in current clamp mode were held at a membrane potential of −70 mV. Near-threshold current steps were applied to determine the rheobase current, then 1 s current steps proportional to the rheobase current were applied. An additional small hyperpolarizing step (30 pA) with a duration of 300 ms was used to determine the time constant. To assess the presence of ChR2, a 2 ms blue light stimulation of ChR2 was performed at a holding potential of −70 mV. Electrical properties were extracted using a custom written MATLAB (MathWorks, Natick, MA, USA) script. The AP characteristics were determined from the first spike that occurred at rheobase current stimulation. FR and adaptation were determined from a current injection of approximately twice the rheobase current. After recording, slices were fixed overnight in 1x PBS containing 4% PFA. Slices were then washed in 1x PBS, followed by three washes in 0.1M PB containing 0.3% Triton X-(PBT). The slices were incubated at 4°C for one day in PBT with 0.002% Streptavidin conjugated to Alexa Fluor 633 (ThermoFisher Scientific, Waltham, MA, USA), then washed two times in PBT and two times in PB. The slices were mounted and imaged with a LSM800 confocal microscope (Zeiss, Oberkochen, Germany) using a 20x objective.
Behavioral setup
The setup used for the self-paced reward-seeking task consisted of a linear track (width 8 cm, length 50 cm) with a platform (width, and length 25 cm) at each end. The track and platforms held 2 cm high walls, and the setup was situated 1.5 m above the floor. Two infrared beam break sensors (Adafruit, Product ID: 2168) controlled by an Arduino were used, one tracking entrance into the trigger zone (red time stamp, Figure 2A), and one tracking entrance of the rat’s snout into the reward port (red time stamp, Figure 2A). All other time stamps were based on video analysis. A spout delivered liquid reward (50 μL 10% sucrose reward, delivered during 2 s) in the reward port situated in the back wall of the reward platform. Reward delivery was triggered by the beam break in the reward port if a beam break had been registered in the trigger zone after the latest beam break in the reward port (= correct trial). There were no delays between trials. In 15% of the correct trials reward delivery was omitted to address trial outcome history.
To accurately align neural activity and behavior, a second Arduino would send pulses (20 Hz) to trigger a Blackfly video camera (FLIR) and to the Digital Lynx 4SX acquisition system, thereby providing a timestamp for each video frame in the electrophysiological recording.
Behavioral tracking
DeepLabCut (DLC)60 was used for tracking animal location. The base of the tail, the end of the tail, and the left and right ears were manually labeled in frames, sampled from all sessions. The behavioral and movement analysis were based on three of the markers tracked by DLC: the base of the tail, and the center of the head (point between left and right ears). Smoothing of tracking data was accomplished using a two-pass Kalman Filter algorithm. The initial state and model parameters were first estimated using an Expectation Maximization algorithm. A second pass was then performed with a modified observation covariance matrix (multiplied by 5) to reduce the trust in the raw observations, thereby generating smoother estimates.
Behavioral task
Definition of a trial: leaving the reward zone (= trial start; black time stamp in Figure 2A), traversal of the linear track to the trigger zone (traversal ‘to trigger’), turning 180° in the trigger zone, traversal of the linear track back to the reward zone (traversal ‘to reward’), reward consumption/omission, turning 180° in the reward zone, leaving the reward zone (= trial end, black time stamp in Figure 2A). Reward was randomly delivered in 85% of the trials with successful visit to the trigger zone.
Behavioral training, including the temporal relation to virus injection, microdrive implantation, and electrophysiological recordings
Behavioral training started 1 w after virus injection. The daily amount of food was restricted to 10 g from the first training day. The first training session the rats could explore the setup and collect reward in the reward port at any point, i.e., no specific behavior was required for reward delivery. When the rats had learned where to collect reward (>100 collected rewards in 30 min; usually within the first training session), task training started: the rat was put in the setup and learned through exploratory behavior that visit in the trigger zone was required for reward to be delivered in the reward port. No instructive signals were given to guide the behavior. The rats were trained in one session per day, and rats conducting ≥50 trials in 1 h for three consecutive days were considered fully trained and were taken off food restriction. After ≥ three days with free access to food, the microdrive was implanted. After 1 w of recovery from the implantation, the rats were put on food restriction again and three days later task retraining started, with the recording cables attached to the microdrive. The recording sessions started the day after the animal successfully performed 50 trials in 1 h (requiring 1–3 days of retraining) and continued daily until the electrophysiological signal was deemed suboptimal. One day = one session behavior/recording (max 1 h) across the study. A total of 10 rats were used for behavior. However, three rats were excluded from further analysis due to either long trial times (median trial time >8.7 s) or a low number of total trials (<381). Both cutoffs were computed by use of the standardized outlier detection method median absolute deviation (MAD), employing a scale factor of 1.5.
Microdrive construction
Microdrives (flexDrive,61 N = 3, or DMCdrive,62 N = 4) were used for chronic in vivo electrophysiology and optogenetics in behaving adult (3–4 months at experiment start) PV-Cre rats (N = 7 included in analysis). Tetrodes consisted of four twisted fine wires (polyimide insulated ni-chrome wire, 12 μm, Sandvik-Kanthal) that were goldplated to reduce the impedance to 0.2–0.4 MΩ at 1 kHz. Four movable tetrodes were loaded into medical-grade polyimide carrier tubes (0.005 inch OD, Phelps Dodge) in the microdrive. The microdrives were equipped with a Ø200 μm multimode optical fiber (0.5 NA, Thorlabs (FP200ERT)).
Microdrive implantation
Burprenorphine was delivered (I.P 0.05 mg/kg) 30 min before surgery. The animals were deeply anesthetized with isoflurane (1.5–2.5%), and the body temperature maintained at 37°C using a heating pad. The animals were fixed in a stereotaxic frame (Harvard Apparatus, Holliston, MA) and a small craniotomy (1.0–1.5 mm diameter) was made unilaterally over the mPFC (AP 3.0–3.3 mm, ML 0.5–0.8 mm). The microdrive was positioned above the craniotomy with the optical fiber and tetrodes gradually lowered to the PrL (2.5 mm ventral to brain surface), aimed at layer 5/6. Five miniature anchoring screws were used to attach the microdrive to the skull (one contralateral to the microdrive, and two rows of two screws on the anterior and posterior part of the parietal bone). One Teflon coated stainless steel wire (0.005 inch bare, A-M systems) from the electrode interface board (EIB) of the microdrive was connected to the screws for grounding. The microdrive was secured onto the skull using dental adhesive cement (Super Bond C&B, Sun Medical). 3 rats were implanted in the right hemisphere and 4 rats in the left hemisphere. After surgery the animals were single housed. Postoperative carpofen (5 mg/kg) was administered every 24 h for 48 h.
Electrophysiological recordings with optogenetics during freely moving behavior
The neuronal activity (unit activity and LFP) was recorded in a total of 97 sessions (N = 7 PV-Cre rats) using a Digital Lynx 4SX acquisition system and Cheetah data acquisition software. The tetrodes were lowered 40–90 μm after every recording session. Unit signals were amplified with the gain of 10,000, filtered with bandwidth 600-6,000 Hz, digitized at 32 kHz, and stored on a PC. LFPs were acquired from one electrode of each tetrode at a sampling rate of 32 kHz. The signal was band-pass filtered between 0.1 and 500 Hz. Laser light (473 nm, 1–3 ms pulse width, 1 s duration, 5-10 stimulations/frequency, 5–10 mW, 10, 20 and 40 Hz) was delivered through the optical fiber from a DPSS blue laser (Cobolt MLD 473 nm, Cobolt) controlled by custom software written in LabVIEW.
Quantification and statistical analysis
Cell sorting
Single neurons were manually sorted using MClust (written by A.D. Redish). Well-separated neurons were defined by isolation Distance >15, L-ratio <0.2, and the spikes <0.01% with <2 ms.63
Identification of directly light-activated neurons
To identify directly light-activated FS-PV neurons we employed an optical-tagging test (Stimulus Associated spike Latency Test, SALT).29 SALT is used to statistically determine if neurons displaying short-latency spikes with low jitter have significantly changed spike timing in relation to the onset of light pulses. To ensure that the spike sorting was not compromised by light application, the Pearson’s correlation coefficient was used for comparison of waveforms of light-evoked spikes to spontaneous spikes for individual neurons. Using these measures (SALT p-value <0.05; Pearson’s correlation coefficient r > 0.95) 13 units were identified as FS-PV neurons.
Cell classification
The units were first classified into wide-spiking (WS) putative pyramidal neurons and narrow-spiking (NS) putative interneurons based on the distribution of (1) the peak-to-valley ratio (the ratio between the amplitude of the initial peak and the following trough), and (2) the spike-width (time from peak to trough) of each spike waveform, chosen from the tetrode with largest spike amplitude. For objective classification of units, a Gaussian mixture model (GMM) was fitted to the units.64 Units with NS probability >0.95 were classified as NS, and units with NS probability <0.30 as WS. Units not fulfilling either of these criteria were not classified. The waveform similarity index (r): the Pearson’s correlation between the mean spike waveform of each unit (WS, NS, or FS-PV) and the mean light-activated spike waveform.
Spectral analysis of the LFP in response to optogenetic stimulation
LFP signals were down sampled to 1 kHz and bandpass filtered between 1 and 500 Hz. Relative power was calculated for three frequency bands (6–14, 16–24, and 36–44 Hz) during epochs (1 s) of light stimulation at different frequencies (10, 20, or 40 Hz), and during baseline (1 s preceding light stimulation), respectively. Power spectral density was estimated in the frequency range 2–100 Hz using superlets.65 The relative power was thereafter calculated by dividing the power within each frequency band, to the total power in the power spectrum. The relative power ratio is Plight/Pbaseline where Plight is the relative power in a frequency band during light stimulation (±4 Hz around the stimulation frequency) and Pbaseline is the relative power in that band in the absence of light stimulation.35 The harmonics preclude analysis of how higher frequency bands are affected by lower frequency stimulations.
Spectral analysis of the LFP during track traversals
Raw LFP signals (low-pass filtered at 500 Hz, 32 kHz sampling rate) were processed and analyzed using custom software written in MATLAB. The LFP signals were down-sampled to 1 kHz and LFP power spectra were estimated by the Chronux function (‘mtspectrumc’). To investigate the relative power of the LFP during traversals on the linear track, only traversals with similar track duration (0.5–1.5 s, spatial bins 2–9 in Figure 2A) were selected. The power within the frequency band of interest was divided by the total power in the 1–100 Hz band and averaged across trials. To analyze the relationship between LFP power and the traversal direction or running speed (included speed range: 15–75 cm/s), LFP power spectrograms (10–100 Hz) were constructed using the continuous wavelet transform (cwt) function with the analytical Morlet wavelet (‘amor’) in MATLAB and normalized by the peak power. To identify gamma-band bursts, the LFP power was averaged across the gamma band (30–50 Hz) and then gamma bursts were detected by a threshold (75^th^ percentile power) crossing method using ‘findpeaks’ function in MATLAB.
Phase-coupling analysis
Spike phase-coupling to ongoing gamma (30–50 Hz) oscillations was analyzed separately for the two traversal directions. For every spike occurring, the corresponding instantaneous phase of the gamma oscillation was extracted from the Hilbert transform. Only neurons with ≥50 spikes in each traversal were included (WS: n = 204, NS: n = 21, FS-PV: n = 13). The strength of phase-coupling of single neurons to gamma oscillations were quantified using pairwise phase consistency (PPC)55 (Figure 6A) and mean resultant length (MRL; pycircstat. resultant_vector_length) (Figure 6B). Significant gamma phase-coupling was based on Rayleigh test (p < 0.05, pycircstat.rayleigh).
Analysis of single neuron data
Spike data was binned by the recorded frames (50 ms bins; see Behavioral setup) and smoothed with a Gaussian filter of 100 ms (scipy.ndimage.Gaussian_filter1d) for all analysis described below.
Generalized linear models (GLMs)
To assess the relationship between neuronal activity and various behavioral and task-related variables, we employed a Generalized Linear Model (GLM) using a Poisson regressor (sklearn PoissonRegressor). Variables included in the models: categorical variables; linear position (the rat’s 1D position in the behavioral setup, in 1 of 10 bins (Figure 2A); run phase (track traversal to reward zone or trigger zone, respectively), trial outcome (correct trial rewarded or not rewarded, respectively), speed, acceleration, and turning, respectively. Additional variables were analyzed but determined to not be significantly encoded: trial time (fast vs. slow, with cutoff at 3 median absolute deviation (MAD) above median trial time), trial history (previous trial rewarded or not), and trial type (current trial will be rewarded or not). Run phase was scored specifically during running, i.e., does not include periods of immobility, or turning on a platform (Figure S3A). The rat’s turning in the platforms was scored by calculation of the head angular velocity, i.e., the angular difference between the center of the rat’s head and the tail base. The continuous variables speed, acceleration, and turning were z-scored.
In the GLM model the activity of each neuron was represented as a linear combination of the behavioral variables, each scaled by its respective coefficient. To identify which neurons to include in the models, we first fitted the activity of the neurons using models with single variables (Figure S3B). Next, we fitted neuronal activity using models with increasing number of variables, until all variables were included. For each model, we quantified the cvR^2^ from the test data (average of held-out data from 5-fold cross-validation) and the training data (average of used data from 5-fold cross-validation), respectively. Importantly, the cross-validation was conducted with the dataset divided into five continuous, non-overlapping segments, ensuring that each fold comprised a continous portion of the data. The predicted explained variance was computed as the cumulative sum of cvR^2^ from single variable models (Figures S3B and S3C).
A separate GLM was created for investigation of the relationship between encoding of spatial context and movement variables (speed, acceleration; Figure 4). For this an additional variable was generated – position with direction – where position contained 10 bins, with 2 variables for each bin: traversal to reward, and traversal to trigger, yielding 20 predictors, each representing a unique combination of linear position and run phase. The GLMs were evaluated using 5-fold cross validation (Sklearn.model_selection KFold) and the goodness of fit of the prediction was quantified as the squared correlation (Pearson’s correlation (r)) between the predicted and the actual test data (sklearn.model_selection cross_val_score).
Two methods were used to quantify encoding of behavioral variables.38^,^39 (I) We created models consisting of single variables and computed the cross-validated explained variance (cvR^2^). This method can overestimate the encoding of a certain variable if that variable is correlated with other variables. Thus, as a second approach (II) we created models with all behavioral variables (full model), followed by shifting one variable in time (circular shift) to generate reduced models. The decrease in explained variance (Δr^2^) for the reduced models, as compared to the full models, reflects the unique encoding of the shifted variable (i.e., the encoding is not captured by other variables). These two measures, thus, provide upper and lower boundaries, respectively, on a given behavioral variable’s explanatory power.38
To examine if the activity of individual neurons correlated to a particular variable, we created reduced models, by shifting one variable in time (circular shift) and quantified the cvR^2^ as described above. To calculate whether the reduction in cvR^2^ was significant, we computed the cvR^2^ for 1000 reduced models. If the cvR^2^ for the full model was greater than 99% of the cvR^2^ derived from reduced models, the neuron was defined as being significantly correlated with that variable.
Correlations between variable encodings
In Figure 2F we correlated (Pearson’s correlation) the unique explained variance (Δr^2^) for all combinations of variables. As a control, Δr^2^-values for each neuron were randomly shuffled prior to computing the correlation between the Δr^2^-values. This step was repeated 1000 times and p-values were computed, by comparing the true correlations with the shuffled control.
Analysis of spatial activity patterns
To analyze neuronal activity patterns associated with the respective traversal direction, we computed activity vectors as a function of linear position (10 bins, same bins as in GLM). This was done separately for the two traversal directions (Figure 3). Sessions with less than 10 to-reward and 10 to-trigger traversals were excluded, as were traversals with duration >10 s, yielding inclusion of activity from 319 neurons (WS: n = 268, NS: n = 38, FS-PV: n = 13). To identify neurons displaying differential activity patterns dependent on the traversal direction, the ‘traversal to trigger’ activity vector was flipped to match the ‘traversal to reward’ activity vector with respect to the start and the end of the traversal. The difference in activity was calculated for each relative track position (bin). As control dataset, the traversal direction was randomly shuffled and the difference between the two activity vectors computed 1000 times.66 Real activity difference that exceeded 97.5% or fell below 2.5% of the shuffled activity differences was defined as significantly more or less active, respectively.
The direction modulation index (MI, Figure 3F) was calculated for the individual neurons to identify FR differences between traversals to reward and traversals to trigger. , where is the mean rate of the to reward activity vector and is the mean rate of the to trigger activity vector. The MI ranges from −1 to 1, 0 = no modulation, i.e., no FR differences between the traversal directions. Positive values: enhanced spiking in traversals to reward, and negative values: enhanced spiking in traversals to trigger. To identify neurons with significant modulation, the trial-averaged mean activity in traversals to reward and traversals to trigger, respectively, was compared (Wilcoxon rank-sum (scipy.stats.ranksums)).
Spatial consistency
Spatial consistency was computed by generating activity vectors (see above) for odd and even trials, respectively, followed by correlation of the activity vectors for the odd and even trials (Pearson’s correlation). Control data was obtained by calculating spatial consistency for shifted (circular shift) neuronal activities 1000 times.
Decoding analysis
We decoded the rats’ traversal direction based on neuronal activity (Figure 3G). For each neuron, we computed activity vectors, together with a separate binary vector, describing the traversal direction (0 = to reward, 1 = to trigger). Using the traversal direction as the label and the vector of activity as corresponding features, we trained support vector machines (SVMs) with linear kernels. For each neuron, we randomly divided 80% of the traversal into a training set and the remaining 20% into a test set. We repeated this step 20 times for each neuron and report the average accuracy of these 20 iterations as real accuracy. As control, we shuffled the labels prior to training the SVMs. This step was repeated 20 times and the average accuracy of those 20 iterations are reported as shuffled accuracy.
We predicted the rats’ traversal direction based on neuronal activity, using SVMs trained on activity of different neurons (Figure 3H). Specifically, for each neuron (test neuron) we randomly selected different neurons (training neurons) of same cell type (with increasing numbers: 1–10 neurons), to train the SVMs. For each training neuron, we randomly chose 15 to reward and 15 to trigger z-scored activity vectors, and used these activity vectors, together with the direction labels, to train SVMs. We thereafter evaluated the ability of the SVMs to accurately predict the traversal direction of the rat based on the activity of the test neuron. As a control, we randomly shuffled the direction labels prior to training the SVMs. This procedure was repeated so that all neurons were used as test neurons.
Correlation between unique encoding and decoding performance
We correlated the decoding performance with the unique contribution (Δr2, see generalized linear models (GLMs) section) of each behavioral variable in the model (Figure 3I). As control, we generated shuffled data by shuffling the Δr^2^ values for each neuron, prior to computing the correlation between decoding performance and Δr^2^ values. The shuffling was repeated 1000 times.
Spatial information
To quantify the spatially related activity in the spatial locations across the linear track (bin 2-9), we computed the spatial information (SI)5 from spatially binned firing rates, separately for the two traversal directions (Figures 6C and 6F). SI was calculated as , where N is the number of bins, pi is the occupation probability in the i^th^ bin, fi is the mean activity in bin i, and f is the average activity of the neuron across all bins.5
Statistics
Across all figure panels, the significant differences are indicated for all analyses performed (∗p < 0.05, ∗∗p < 0.01, ∗∗∗p < 0.001), i.e., non-significant (ns) differences are not indicated.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Harvey C.D.Coen P.Tank D.W.Choice-specific sequences in parietal cortex during a virtual-navigation decision task Nature 4842012626810.1038/nature 1091822419153 PMC 3321074 · doi ↗ · pubmed ↗
- 2Mao D.Kandler S.Mc Naughton B.L.Bonin V.Sparse orthogonal population representation of spatial context in the retrosplenial cortex Nat. Commun.8201724310.1038/S 41467-017-00180-9PMC 555792728811461 · doi ↗ · pubmed ↗
- 3Long X.Zhang S.J.A novel somatosensory spatial navigation system outside the hippocampal formation Cell Res.31202164966310.1038/S 41422-020-00448-833462427 PMC 8169756 · doi ↗ · pubmed ↗
- 4Mertens P.E.C.Marchesi P.Ruikes T.R.Oude Lohuis M.Krijger Q.Pennartz C.M.A.Lansink C.S.Coherent mapping of position and head direction across auditory and visual cortex Cereb. Cortex 3320237369738510.1093/CERCOR/BHAD 04536967108 PMC 10267650 · doi ↗ · pubmed ↗
- 5Sauer J.F.Folschweiller S.Bartos M.Topographically organized representation of space and context in the medial prefrontal cortex Proc. Natl. Acad. Sci. USA 1192022 e 211730011910.1073/pnas.2117300119 PMC 883319935121665 · doi ↗ · pubmed ↗
- 6Fuster J.M.The Prefrontal Cortex 201510.1016/C 2012-0-06164-9 · doi ↗
- 7Ma X.Zheng C.Chen Y.Pereira F.Li Z.Working memory and reward increase the accuracy of animal location encoding in the medial prefrontal cortex Cereb. Cortex 3320232245225910.1093/CERCOR/BHAC 20535584788 PMC 9977377 · doi ↗ · pubmed ↗
- 8Hyman J.M.Ma L.Balaguer-Ballester E.Durstewitz D.Seamans J.K.Contextual encoding by ensembles of medial prefrontal cortex neurons Proc. Natl. Acad. Sci. USA 10920125086509110.1073/pnas.111441510922421138 PMC 3323965 · doi ↗ · pubmed ↗