Grappling with grapevine guardians: Unraveling the dynamic dance of VviWRKY10 and VviWRKY30 in powdery mildew defense
Ritu Singh

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
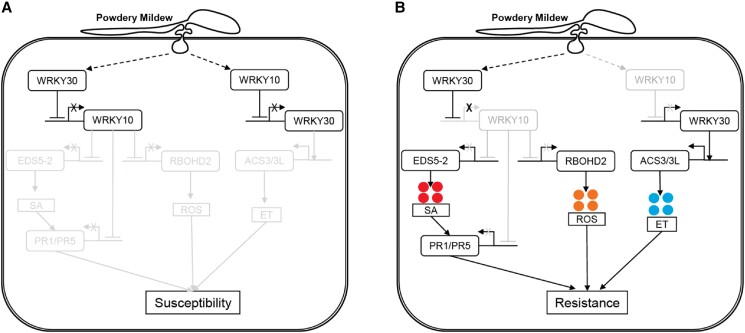 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant-Microbe Interactions and Immunity · Plant Gene Expression Analysis · Powdery Mildew Fungal Diseases
Plants face constant threats from various microbial pathogens, posing a significant risk to global food security. To counter these threats, plants have evolved sophisticated defense mechanisms primarily involving physical and chemical strategies to protect themselves against these microbes (Jones and Dangl 2006; Ramirez-Prado et al. 2018). In addition to these preformed physical/chemical barriers, plants actively detect pathogens and trigger defense signaling networks, restricting the further growth and spread of pathogens. Transcription factors (TFs) play a central role in these signaling networks by governing the expression of defense genes. Exploring TF regulation during plant-pathogenic fungi interactions opens avenues to discover novel virulence factors and enhance our understanding of the regulatory networks associated with pathogen evolution. Moreover, targeting TFs themselves can be a strategy for disease control.
Grapevine (Vitis vinifera) is one of the most economically important fruits globally. However, its production is severely hampered by Erysiphe necator, an obligate biotrophic fungal pathogen that causes powdery mildew disease. Although previous research identified AtWRKY18 and AtWRKY40 as negative regulators of powdery mildew resistance in Arabidopsis (Xu et al. 2006, Pandey et al. 2010), the specific role of WRKY TFs in grapevine remained elusive.
In the recent Plant Physiology issue, Zhou and colleagues (2024) explored the function of VviWRKY10 and VviWRKY30 (orthologs of AtWRKY18 and AtWRKY40) in grapevine powdery mildew resistance. The authors used the CRISPR-Cas9 method to generate single and double mutant lines in the powdery mildew–susceptible Cabernet Sauvignon grape cultivar. They successfully obtained 53 wrky10 mutants and 15 wrky10wrky30 mutants, with no wrky30 mutant plants. Upon infection with E. necator NAFU1, wrky10 plants displayed strong resistance, whereas wrky10wrky30 showed moderate resistance compared to wild-type plants. Further, trypan blue, 3,3'-diaminobenzidine, and aniline blue staining revealed enhanced hypersensitive response–like cell death, H_2_O_2_, and callose accumulation, respectively, in wrky10 lines, moderate in wrky10wrky30, and none in wild-type.
Because no wrky30 CRISPR-Cas9 mutants were obtained, the authors transiently overexpressed VviWRKY30 along with VviWRKY10 in Cabernet Sauvignon leaves through Agrobacterium-mediated transformation. Leaves overexpressed with VviWRKY30 showed increased resistance against powdery mildew, whereas leaves overexpressed with VviWRKY10 exhibited more susceptibility compared to the wild type, indicating their opposite roles in powdery mildew resistance. Further, real-time expression analysis identified that VviWRKY10 was induced at the early stage of infection (peaking at 24 hpi), whereas VviWRKY30 was induced at a later time (peaking at 48 hpi), suggesting their distinct roles in the transcriptional regulation of host defense at different infection stages.
WRKY TFs can modulate the levels of salicylic acid (SA) and ethylene (ET) hormones during plant–pathogen interactions (Dang et al. 2014; Birkenbihl et al. 2017). Zhou et al. (2024) selected key genes involved in SA and ET hormone signaling and response and measured their expressions in E. necator-inoculated mutants and wild-type plant leaves. The result indicated significantly increased expression of most of these genes in both wrky10wrky30 and wrky10, with a more specific upregulation in wrky10 lines. To further validate this, the authors quantified SA and ET levels in leaf tissues of wild-type and mutant lines, which confirmed higher SA levels in both mutants and elevated ET in wrky10. This suggests that VviWRKY10 and VviWRKY30 influence grapevine powdery mildew resistance by altering SA and ET.
WRKY TFs specifically bind to W-box (TTGACC/T) elements on the promoters of the target genes (Rushton et al. 2010). Chromatin immunoprecipitation-quantitative real-time PCR assays and electrophoretic mobility shift assays analysis demonstrated that VviWRKY10 and VviWRKY30 could directly bind to the promoters of key genes in SA and ET pathways, such as VviEDS5-2, VviPR1, VviPR5, VviACS3/ACS3L, and VviRBOHD2. Further, dual-luciferase assays suggest that VviWRKY10 inhibited the transcription of SA- and ET-related genes, whereas VviWRKY30 significantly enhanced the transcription of ET-related genes. Moreover, VviWRKY10 and VviWRKY30 can mutually inhibit each other by binding to each other's promoters.
In summary, the study highlights the distinctive roles of VviWRKY10 and VviWRKY30 in powdery mildew resistance by modulating SA and ET hormones (Figure). VviWRKY10 suppresses the SA-dependent defense responses upon powdery mildew infection, whereas VviWRKY30 promotes ET production for limiting powdery mildew growth at a later stage of nfection. Furthermore, VviWRKY10 and VviWRKY30 can inhibit each other to inhibit ET synthesis and SA and ROS production, respectively. The mutual inhibition of VviWRKY10 and VviWRKY30 and their distinct functions might be crucial for balanced defenses through regulating multiple hormonal pathways against powdery mildew and potentially other pathogens in grapevines. Additionally, these genes appear to be linked to plant growth, as mutants displayed reduced height and growth compared to the wild type, highlighting the intricate relationship between growth and defense. These genes stand out as valuable tools for studying this delicate balance.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Birkenbihl RP , Kracher B, Roccaro M, Somssich IE. Induced genome-wide binding of three Arabidopsis WRKY transcription factors during early MAMP-triggered immunity. Plant Cell.2017:29(1):20–38. 10.1105/tpc.16.0068128011690 PMC 5304350 · doi ↗ · pubmed ↗
- 2Dang F , Wang Y, She J, Lei Y, Liu Z, Eulgem T, Lai Y, Lin J, Yu L, Lei D, et al Overexpression of Ca WRKY 27, a subgroup I Ie WRKY transcription factor of Capsicum annuum, positively regulates tobacco resistance to Ralstonia solanacearum infection. Physiol Plant.2014:150(3):397–411. 10.1111/ppl.1209324032447 · doi ↗ · pubmed ↗
- 3Jones JD , Dangl JL. The plant immune system. Nature. 2006:444(7117):323–329. 10.1038/nature 0528617108957 · doi ↗ · pubmed ↗
- 4Pandey SP , Roccaro M, Schön M, Logemann E, Somssich IE. Transcriptional reprogramming regulated by WRKY 18 and WRKY 40 facilitates powdery mildew infection of Arabidopsis. Plant J.2010:64(6):912–923. 10.1111/j.1365-313X.2010.04387.x 21143673 · doi ↗ · pubmed ↗
- 5Ramirez-Prado JS , Abulfaraj AA, Rayapuram N, Benhamed M, Hirt H. Plant immunity: from signaling to epigenetic control of defense. Trends Plant Sci.2018:23(9):833–844. 10.1016/j.tplants.2018.06.00429970339 · doi ↗ · pubmed ↗
- 6Rushton PJ , Somssich IE, Ringler P, Shen QJ. WRKY transcription factors. Trends Plant Sci.2010:15(5):247–258. 10.1016/j.tplants.2010.02.00620304701 · doi ↗ · pubmed ↗
- 7Xu X , Chen C, Fan B, Chen Z. Physical and functional interactions between pathogen-induced Arabidopsis WRKY 18, WRKY 40, and WRKY 60 transcription factors. Plant Cell.2006:18(5):1310–1326. 10.1105/tpc.105.03752316603654 PMC 1456877 · doi ↗ · pubmed ↗
- 8Zhou M , Wang HY, Yu XN, Cui K, Hu Y, Xiao S, Wen YQ. Vvi WRKY 10 and Vvi WRKY 30 co-regulate powdery mildew resistance through modulating SA and ET-based defenses in grapevine. Plant Physiol.2024;kiae 080. 10.1093/plphys/kiae 08038366578 · doi ↗ · pubmed ↗