Plastid translation as a developmental checkpoint? Plastid ribosomal protein EMB27 is required for maize embryogenesis
Janlo M Robil

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
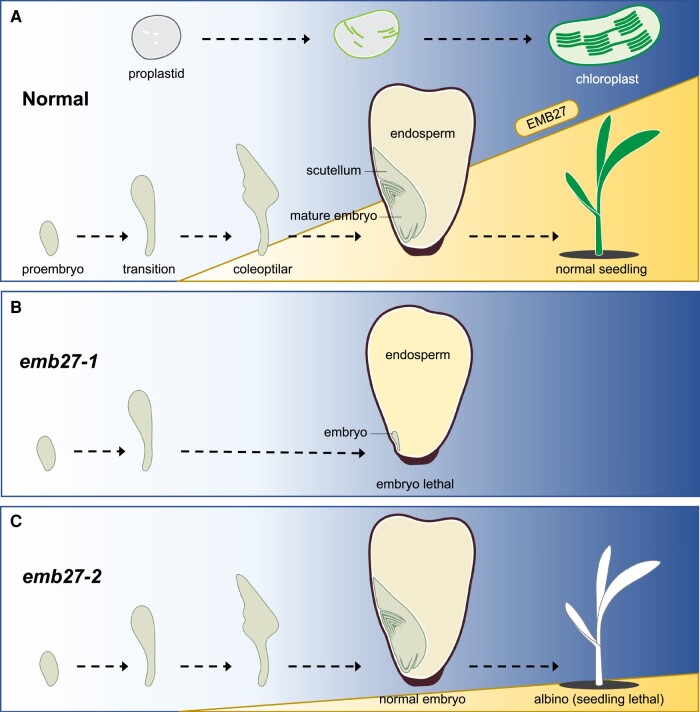 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPhotosynthetic Processes and Mechanisms · Plant Molecular Biology Research · Plant nutrient uptake and metabolism
Plant embryos undergo crucial development to establish themselves for photosynthesis. It begins at the early stages of embryogenesis with the formation of plastids in cells of nascent ground and epidermal tissues (West and Harada 1993). Genetic screens in Arabidopsis and maize have revealed embryo lethality in numerous chloroplast-defective mutants (Meinke 2020). However, the observed phenotypic variations across different species and genetic backgrounds complicate our understanding of the relationship between these 2 defects (Parker et al. 2014). Therefore, how chloroplast biogenesis influences plant embryogenesis remains unclear.
In this issue of Plant Physiology, Liu et al. (2024) explore the roles of EMB27, a plastid ribosomal protein S13 (RPS13), in maize development. Through genetic experiments and biochemical assays, the authors provide evidence that EMB27 is involved in plastid protein translation, required in both plastid and embryo development (Fig. 1A). The functional analysis of EMB27 offers some insights into the yet-to-be-resolved link between chloroplast biogenesis and plant embryogenesis.
The investigation started with the isolation of the embryo defective27-1 (emb27-1) mutant from a transposon insertion population. A mature maize kernel is composed of an embryo that includes a scutellum, which resembles a shield-like cotyledon, all of which are enclosed within the pericarp alongside the persistent endosperm (Fig. 1A). emb27-1 exhibits a premature arrest in embryo development, whereas the development of the endosperm remains unaffected (Fig. 1B). EMB27-1 encodes a highly conserved RPS13, a key component of the small subunit of the plastid ribosome. Interestingly, the authors identified a weaker allele, emb27-2, exhibiting normal embryo development but displaying albinism and seedling lethality due to its inability to produce functional chloroplasts (Fig. 1C). The authors leveraged both the strong and weak emb27 mutant alleles to decouple the processes of embryo and chloroplast development.
The divergent phenotypes of the 2 emb27 alleles led the authors to consider whether a threshold level of EMB27 expression is required for normal embryo development. To test this hypothesis, they quantified EMB27 transcript levels in the embryos of both mutants and detected no transcript in emb27-1 but at least 10% of normal levels in emb27-2. This result suggests that emb27-1 is a null allele while emb27-2 is of partial loss of function, which could explain the normal embryo development in emb27-2. Hence, the authors concluded that emb27-2 expresses enough EMB27 during embryogenesis to reach a developmental threshold but not enough to sustain chloroplast biogenesis throughout seedling growth.
The authors then tested the correlation between EMB27 expression and embryo development progression. Maize early embryo development has 3 distinct morphological phases: the ovoid proembryo, the club-shaped transition, and the spade-shaped coleoptilar stage (Vernoud et al. 2005). However, the null mutant emb27-1 exhibits developmental arrest only at the transition stage. Therefore, the authors crossed emb27-1/+ with emb27-2/+ to generate trans-heterozygotes that would arrest at different stages of embryo development. As predicted, the trans-heterozygote progeny displayed a spectrum of phenotypes, ranging from albino seedlings to defective embryos with varying severity. The authors quantified EMB27 transcript levels in these embryos and uncovered a negative correlation between the EMB27 expression level and the severity of the phenotype. Therefore, this result supports the conclusion that sufficient EMB27 expression is progressively required throughout maize embryogenesis.
The chloroplast defects in albino emb27-2 seedlings underscore the role of EMB27 in chloroplast biogenesis. Considering that EMB27 is integral to the ribosomal small subunit, the authors probed the impact of its mutation on the formation of the subunit and the synthesis of chloroplast proteins. They found a significant reduction in the small subunit-associated 16s rRNA, possibly due to its degradation from failing to form a stable complex with the subunit. Additionally, membrane-intrinsic chloroplast proteins, including photosystems and ATP synthase, were undetectable. These findings point to compromised ribosomal subunit assembly and plastid protein translation in the emb27-2 mutant, thus signifying the essential role of EMB27 in these processes during seedling development in maize.
Because some plastid transcripts require plastid-encoded proteins for splicing, the authors investigated whether this process was affected in both emb27-1 and emb27-2 mutants. RT-PCR analysis revealed altered splicing of introns in 4 plastid transcripts in emb27-1 but not in emb27-2. This finding suggests that splicing of plastid transcripts is an early function of EMB27 and potentially contributes to the observed embryogenesis defects. Furthermore, this splicing impairment is shared with other maize mutants exhibiting both plastid translation and embryogenesis defects (Zhang et al. 2013; Xu et al. 2021), supporting the general requirement for specific plastid transcript splicing by plastid-encoded proteins during embryogenesis.
Previous studies on maize have found that mutants with plastid-translation defects exhibit either embryo-defective or albino seedling phenotypes, depending on the genetic background (Zhang et al. 2013; Li et al. 2015; Xu et al. 2021). In the present study, Liu et al. (2024) similarly observed this phenomenon in emb27 mutants. When heterozygous emb27-1/+ from a near-isogenic background was crossed with a different inbred line, the hybrid F2 progeny segregated both embryo-defective and albino mutants. This finding suggests that the genetic background influences the expressivity of EMB27, consistent with previous reports of genetic modifiers associated with plastid translation in Arabidopsis (Parker et al. 2014). However, Liu et al. (2024) provide further evidence that, unlike in Arabidopsis, the background effect observed in maize does not stem from the disruption of metabolic processes but rather from the loss of a chloroplast signal essential for embryo development (Shen et al. 2013; Zhang et al. 2013).
Liu et al. (2024) offer new insights into the role of a plastid ribosomal protein in maize embryogenesis. Their study decouples the embryo and chloroplast defects observed in emb27 mutants, providing a valuable roadmap for future investigations into the downstream mechanisms of plastid protein translation. These mechanisms might underpin “retrograde signaling pathways” (Chan et al. 2016) responsible for coordinating a systemic response during embryogenesis. Thus, exploration of these mechanisms could illuminate how chloroplasts act as checkpoints, controlling developmental progression in plant embryos.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Chan KX , Phua SY, Crisp P, Mc Quinn R, Pogson BJ. Learning the languages of the chloroplast: retrograde signaling and beyond. Annu Rev Plant Biol. 2016:67(1):25–53. 10.1146/annurev-arplant-043015-11185426735063 · doi ↗ · pubmed ↗
- 2Li C , Shen Y, Meeley R, Mc Carty DR, Tan BC. Embryo defective 14 encodes a plastid-targeted c GT Pase essential for embryogenesis in maize. Plant J. 2015:84(4):785–799. 10.1111/tpj.1304526771182 · doi ↗ · pubmed ↗
- 3Liu XY , Jiang RC, Ma B, Wang Y, Yang YZ, Xu C, Sun F, Tan BC. Maize requires Embryo defective 27 for embryogenesis and seedling development. Plant Physiol. 2024:195(1):430–445. 10.1093/plphys/kiae 01038198212 · doi ↗ · pubmed ↗
- 4Meinke DW . Genome-wide identification of EMBRYO-DEFECTIVE (EMB) genes required for growth and development in Arabidopsis. New Phytol. 2020:226(2):306–325. 10.1111/nph.1607131334862 · doi ↗ · pubmed ↗
- 5Parker N , Wang Y, Meinke D. Natural variation in sensitivity to a loss of chloroplast translation in Arabidopsis. Plant Physiol. 2014:166(4):2013–2027. 10.1104/pp.114.24905225336520 PMC 4256881 · doi ↗ · pubmed ↗
- 6Shen Y , Li C, Mc Carty DR, Meeley R, Tan BC. Embryo defective 12 encodes the plastid initiation factor 3 and is essential for embryogenesis in maize. Plant J.2013:74(5):792–804. 10.1111/tpj.1216123451851 · doi ↗ · pubmed ↗
- 7Vernoud V , Hajduch M, Khaled AS, Depège N, Rogowsky PM. Maize embryogenesis. Maydica. 2005:50:469–483.
- 8West M , Harada JJ. Embryogenesis in higher plants: an overview. Plant Cell. 1993:5(10):1361–1369. 10.2307/386978812271035 PMC 160368 · doi ↗ · pubmed ↗