α-Spectrin regulates cell shape changes during disassembly of microtubule-driven protrusions in Drosophila wings
Ngan Vi Tran, Martti P. Montanari, Dmitri Lubenets, Léa Louise Fischbach, Hanna Antson, Yasushi Okada, Yukitaka Ishimoto, Tambet Tõnissoo, Osamu Shimmi

TL;DR
This paper explores how α-Spectrin helps cells change shape during wing development in fruit flies.
Contribution
The study reveals α-Spectrin's role in regulating microtubule and actomyosin networks during cell shape changes in Drosophila wings.
Findings
α-Spectrin regulates microtubule and actomyosin networks during protrusion disassembly.
Apical relaxation of wing epithelia requires α-Spectrin during protrusion disassembly.
α-Spectrin contributes to 3D tissue morphogenesis in Drosophila pupal wings.
Abstract
The dynamics of microtubule-mediated protrusions, termed Interplanar Amida Network (IPAN) in Drosophila pupal wing, involve cell shape changes. The molecular mechanisms underlying these processes are yet to be fully understood. This study delineates the stages of cell shape alterations during the disassembly of microtubule protrusions and underscores the pivotal role of α-Spectrin in driving these changes by regulating both the microtubule and actomyosin networks. Our findings also demonstrate that α-Spectrin is required for the apical relaxation of wing epithelia during protrusion disassembly, indicating its substantial contribution to the robustness of 3D tissue morphogenesis.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
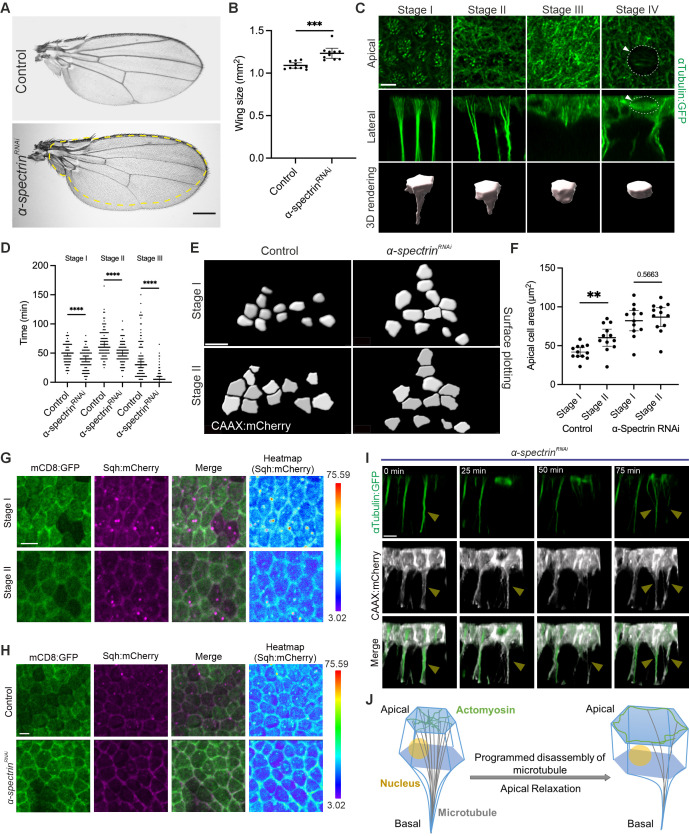 Figure 1
Figure 1|
STRAIN |
GENOTYPE |
AVAILABLE FROM |
|
|
|
Bloomington Drosophila Stock Center (BDSC) #3041 |
|
|
|
BDSC #25754 |
|
|
|
BDSC #42801 (Fig.1 A, B)
BDSC #56932 (
|
|
|
|
BDSC #7017 |
|
|
|
BDSC #59024 |
|
|
|
BDSC #59021 |
|
|
|
BDSC #32186 |
|
|
|
From C. Gonzalez
|
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCellular Mechanics and Interactions · Erythrocyte Function and Pathophysiology · Nuclear Structure and Function
Description
Wing development in
- Drosophila* is a classical model for elucidating genetic regulation of tissue morphogenesis. The larval wing imaginal disc, a monolayer epithelium, is widely utilized to study the integration of diverse signaling molecules in tissue growth and pattern formation (Tripathi and Irvine, 2022) . During metamorphosis, this monolayer transforms into a bilayered primordial wing, comprising dorsal and ventral epithelia. The developmental process within the first 24 hours after puparium formation (APF) unfolds into three distinct phases. Initially, the wing disc evolves into a bilayered structure during the first phase (0-10h APF, first apposition). In the subsequent phase (10-20h APF, inflation), the two epithelia separate and later re-appose during the third phase (at 20h APF, second apposition) (Fristrom et al., 1993; Gui et al., 2019; Montanari et al., 2022) . Notably, cell proliferation in the pupal wing primarily occurs during the inflation stage (Etournay et al., 2016; Milan et al., 1996) . Our latest research highlights the critical function of a microtubule-mediated protrusion network, which we have termed IPAN in driving proliferation within 3D epithelial tissues (Tran et al., 2024) . The IPAN sustains the connection between the two-layered epithelia of the inflated pupal wing and then loses cell-cell contacts through the degeneration of microtubules, leading to coordinated mitoses.
To understand the molecular mechanisms underlying pupal wing development, we carried out in vivo RNAi screening using a pupal wing-specific approach. We selected candidate genes based on the adult wing phenotypes that exhibited larger or smaller sizes than the control, or showed inflated structures. Knockdown of α-spectrin revealed it one of the candidate genes that resulted in a larger adult wing size. Spectrin proteins function with multiple interacting partners, including microtubules and actin filaments (Deng et al., 2020; Fletcher et al., 2015; Forest et al., 2018; Khanal et al., 2016) . When α-spectrin was conditionally knocked down in wing epithelia using the nub-Gal4 driver (Forest et al., 2018) , an increase in wing tissue size was observed ( Fig. 1A, B ). Therefore, we hypothesize that α-Spectrin may be critical in regulating IPAN-mediated proliferation.
Since the dynamics of microtubule protrusions involve cell shape changes (Ko et al., 2019; Matis, 2020; Plochocka et al., 2021; Singh et al., 2018) , we clarify the changes in cellular structures over time and designate the following stage numbers ( Fig. 1C ):
Stage I : The majority of αTubulin is utilized for membrane protrusions, with ~30 microtubule foci per cell found medio-apically (Tran et al., 2024) .
Stage II : Vertical microtubule projections are partially degenerated, and the number of microtubule foci decreases to 10 or fewer per cell.
Stage III : Vertical microtubule projections are mostly degenerated; accordingly, basal protrusions are lost, and a lateral fiber of microtubules is observed medio-apically and cortically.
Stage IV : αTubulin is utilized to form the mitotic spindle, and mitotic cell rounding occurs (dotted line) (Gibson et al., 2006) .
To understand the time-dependent alterations in cells, we selected 25 cells from time-lapse imaging, repeated across four independent datasets, and analysed the cellular stages. We measured the duration of stages I through III by tracking individual cells ( Fig. 1D ). We then analyzed the cellular stages of conditional knockdown in * α-spectrin ^RNAi^ * wings. We found that the degeneration of stage I microtubule protrusions occurred faster in * α-spectrin ^RNAi^ * wings than in control wings. When comparing the lifetimes of stages II and III, both were significantly shorter in * α-spectrin ^RNAi^ * wings than in control wings ( Fig. 1D ). These data suggest that α-Spectrin-mediated cell shape changes are one of the means to regulate IPAN-mediated proliferation by controlling the temporal sequence of gradual changes in microtubule structures from vertical protrusions to mitotic spindles.
Interestingly, we found that apical cell size fluctuates during cell shape changes while microtubule protrusions are disassembled. When we measured the apical cell area by tracking individual cells between stage I and stage II ( Fig. 1E ), we observed that the apical area is smaller in stage I and then increases in stage II, a process termed apical relaxation ( Fig. 1E, F ) (Godard et al., 2020) . When α-Spectrin was conditionally knocked down, apical areas are larger size than in control in stage I, and do not show significant expansion from stage I to II ( Fig. 1E, F ). Since it has been argued that medioapical Myosin II localization is tightly linked to the apical constriction of epithelial cells (Booth et al., 2014; Gillard et al., 2021; Le and Chung, 2021; Toddie-Moore et al., 2022) , we imaged live membrane-bound mCD8:GFP and Spaghetti squash (Sqh):mCherry (myosin II light chain)-expressing pupal wings in stage I and II. We found that substantial amounts of Sqh:mCherry were observed medioapically and cortically in stage I. Most Sqh:mCherry was then detected cortically, but not medioapically, in stage II, corroborating apically constricted cells relaxing from stage I to stage II ( Fig. 1G ). We further attempted to investigate the spatial distribution of Sqh:mCherry in α-spectrin knockdown tissue ( Fig. 1H ). Our data suggest that α-Spectrin is required for apical relaxation between stage I and stage II, likely mediated through the regulation of MyoII localization.
Moreover, while observing time-lapse imaging of αTubulin:GFP and CAAX:mCherry in both control and * α-spectrin ^RNAi^ * wings, we noticed that the dynamics of cell shapes are affected in * α-spectrin ^RNAi^ * wings. When microtubule protrusions are disassembled, membrane protrusions are also disassembled simultaneously in control tissue. In contrast, membrane protrusions are occasionally observed even after microtubule protrusion disassembly and the mitotic phase in * α-spectrin ^RNAi^ * wings ( Fig. 1I ). Strikingly, microtubule protrusions are often regenerated after mitosis if membrane protrusions remained intact ( Fig. 1I ).
These results indicate that α-Spectrin plays an important role in regulating cell shape changes related to the timing of stages of the IPAN structure and the robustness of IPAN structure dynamics. Therefore, α-Spectrin is a crucial factor in supporting tissue homeostasis during IPAN-mediated morphogenesis (Fig 1J).
Methods
** Drosophila genetics **
Crosses were set up and maintained at room temperature (~22ºC) to generate pupal wings for imaging RNAi knockdown transgene expression phenotypes. Pupae from crosses with RNAi transgene-bearing flies were transferred to 29⁰C 16 hours before collecting white pupae. These white pupae were then kept at 29⁰C until imaging commenced at 10.5h after puparium formation (APF) (equivalent to 13.5h APF at 25⁰C). Head eversion served as a developmental marker to assess the suitability of pupae for live imaging. Male pupae were selected for imaging due to their slightly smaller size, and to maximize αTubulin:GFP expression, benefiting from dosage compensation (the ubi>αTubulin:GFP transgene is located on the X-chromosome).
Full genotypes
Control: * w, ubi>αTubulin:GFP; nub-Gal4, ubi>cnn:RFP /+; tub>Gal80 ^ts^ /+ *
α-spectrin ^RNAi^ : * w, ubi>αTubulin:GFP; nub-Gal4, ubi>cnn:RFP/+; tub>Gal80 ^ts^ / UAS-α-spectrin ^RNAi^ *
Figure 1C : w, ubi>αTubulin:G FP (X)
Control: * w, ubi>αTubulin:GFP; ubi>cnn:RFP/+; tub>Gal80 ^ts^ /+ *
α-spectrin ^RNAi^ * : * w, ubi>αTubulin:GFP; nub-Gal4, ubi>cnn:RFP/UAS-α-spectrin ^RNAi^ ; tub>Gal80 ^ts^ /+ *
Control: * w, ubi>αTubulin:GFP; nub-Gal4>UAS-CAAX:mCherry/+; tub>Gal80 ^ts^ /+ *
α-spectrin ^RNAi^ : * w, ubi>αTubulin:GFP; nub-Gal4>UAS-CAAX:mCherry/UAS-α-spectrin ^RNAi^ ; tub>Gal80 ^ts^ /+ *
Figure 1G : * w; ap-Gal4>UAS-mCD8:GFP/+; tub>Gal80 ^ts^ / Sqh:mCherry *
Control: * w; ap-Gal4>UAS-mCD8:GFP/+; tub>Gal80 ^ts^ / Sqh:mCherry *
α-spectrin ^RNAi^ : * w; ap-Gal4>UAS-mCD8:GFP/ UAS-α-spectrin ^RNAi^ ; tub>Gal80 ^ts^ / Sqh:mCherry *
Figure 1I : * w, ubi>αTubulin:GFP; nub-Gal4>UAS-CAAX:mCherry/UAS-α-spectrin ^RNAi^ ; tub>Gal80 ^ts^ /+ *
Preparing pupal wings for live imaging & Live imaging of pupal wings
The live imaging protocol has been described previously (Tran et al., 2024) .
The live imaging process was performed using a Leica TCS SP8 STED 3X CW 3D. The raw confocal data was saved as .lif files and later converted to .ims files using Imaris File Converter from Oxford Instruments. All the subsequent steps were completed in Imaris from Oxford Instruments.
Quantifying cell distribution across stages
Samples were live-imaged at 5-minute intervals for four hours (10.5h - 14.5h APF at 29°C) as described previously (Tran et al., 2024) . Cell distribution analysis was performed using αTubulin:GFP data to classify stage I, stage II, stage III, and stage IV in 25 random individual cells per wing. The experiment was repeated four times.
Quantifying apical cell areas
To visualize the apical cell region, CAAX:mCherry was expressed under the control of nub-Gal4 in both control and α-spectrin RNAi. Samples were live-imaged in for two hours (10.5h - 12.5h APF at 29°C). To analyze apical cell areas in control and α-spectrin RNAi samples, the apical surfaces of individual cells in the ROI region were measured during stage I and stage II.
Reagents
**: **
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Booth AJR Blanchard GB Adams RJ Röper K 201469 A dynamic microtubule cytoskeleton directs medial actomyosin function during tube formation.Dev Cell 2951534-580756257610.1016/j.devcel.2014.03.02324914560 PMC 4064686 · doi ↗ · pubmed ↗
- 2Deng H Yang L Wen P Lei H Blount P Pan D 202046 Spectrin couples cell shape, cortical tension, and Hippo signaling in retinal epithelial morphogenesis.J Cell Biol 21940021-952510.1083/jcb.20190701832328630 PMC 7147103 · doi ↗ · pubmed ↗
- 3Etournay R Merkel M PopovićM Brandl H Dye NA Aigouy B Salbreux G Eaton S Jülicher F 2016526 Tissue Miner: A multiscale analysis toolkit to quantify how cellular processes create tissue dynamics.Elife 510.7554/e Life.1433427228153 PMC 4946903 · doi ↗ · pubmed ↗
- 4Fletcher GC Elbediwy A Khanal I Ribeiro PS Tapon N Thompson BJ 2015223 The Spectrin cytoskeleton regulates the Hippo signalling pathway.EMBO J 3470261-418994095410.15252/embj.20148964225712476 PMC 4388601 · doi ↗ · pubmed ↗
- 5Forest E Logeay R Géminard C Kantar D Frayssinoux F Heron-Milhavet L Djiane A 2018111 The apical scaffold big bang binds to spectrins and regulates the growth of Drosophila melanogaster wing discs.J Cell Biol 21730021-95251047106210.1083/jcb.20170510729326287 PMC 5839784 · doi ↗ · pubmed ↗
- 6Fristrom D Wilcox M Fristrom J 199321 The distribution of PS integrins, laminin A and F-actin during key stages in Drosophila wing development.Development 11720950-199150952310.1242/dev.117.2.5098330522 · doi ↗ · pubmed ↗
- 7Gibson MC Patel AB Nagpal R Perrimon N 200689 The emergence of geometric order in proliferating metazoan epithelia.Nature 44271060028-08361038104110.1038/nature 0501416900102 · doi ↗ · pubmed ↗
- 8Gillard G Girdler G Röper K 202172 A release-and-capture mechanism generates an essential non-centrosomal microtubule array during tube budding.Nat Commun 1214096409610.1038/s 41467-021-24332-034215746 PMC 8253823 · doi ↗ · pubmed ↗