Callicarpayongshunensis (Lamiaceae): A new species from Hunan, China

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1 Figure 2
Figure 2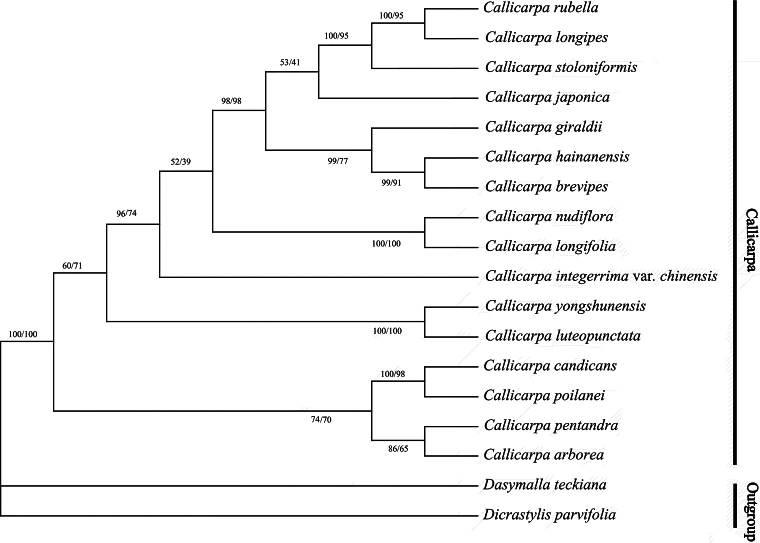 Figure 3
Figure 3 Figure 4
Figure 4| Taxon | ETS | ITS |
|
|
|
|
|
|---|---|---|---|---|---|---|---|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| # | # |
|
|
|
|
|
|
| # |
|
|
|
|
|
|
| Characters |
|
|
|
|
|---|---|---|---|---|
| Life form | procumbent shrub | erect shrub | erect shrub | erect shrub |
| Leaf habit | evergreen | deciduous | deciduous | evergreen |
| Leaf size | 7.5–16 × 3.5–5.5 cm | 7–16 × 2–5 cm | 5–17 × 2–10 cm | 8–20 × 2–6 cm |
| nodes with a transverse scar | absent | absent | absent | present |
| Calyx length | ca. 0.7 mm | ca. 0.7 mm | ca. 1.5 mm | ca. 1 mm |
| Corolla colour | light red to light purple | purple | purple | pale purple |
| Corolla length | ca. 1.3 mm | ca. 1.4 mm | ca. 3 mm | ca. 2 mm |
| Fruit diameter | ca. 2 mm | ca. 1 mm | 2.5–4 mm | ca. 1.5 mm |
| Fruit colour | white | red ( | purple | white |
| Mature fruit overlay | glabrous with yellow glands | glabrous with yellow glands | glabrescent without glands | stellate pubescent with sparse yellow glands |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant and Fungal Species Descriptions · Plant Diversity and Evolution · Phytochemistry and Biological Activities
Introduction
The genus Callicarpa L. was established by Linnaeus (1753), but the family to which it belongs was not defined at that time. Brown (1810) assigned the genus in the family Verbenaceae, which, for decades, was widely recognised by most taxonomists (Endlicher 1836; Schauer 1847; Briquet 1895; Moldenke 1936; Fang 1982; Chen and Michael 1994). However, significant adjustment and delimitation of Lamiaceae and Verbenaceae have been proposed in the past three decades, based on morphological features, especially inflorescence characteristics and molecular systematics (Cantino et al. 1992; Wagstaff and Olmstead 1997; Olmstead et al. 2001; Harley et al. 2004; Ma et al. 2015; Zhao et al. 2021). Consequently, the genus Callicarpa was re-assigned into the Lamiaceae family along with the subfamily Viticoideae and some groups that originally belonged to the family Verbenaceae. Recent molecular phylogenetic studies have further indicated that the genus Callicarpa belongs to the Lamiaceae family, suggesting that it is sister to the endemic Australian subfamily Prostantheroideae, with both groups being early diverging lineages in the family (Mint Evolutionary Genomics Consortium 2018; Zhao et al. 2021). Species delimitation within the genus is typically based on traits, such as life form, number of inflorescence branches, glandular colour, filament length, mode of anther opening, fruit colour and size, as well as presence or absence of stellate hairs (Fang 1982; Chen and Michael 1994).
The genus Callicarpa harbours approximately 140 species and it is distributed in the tropical to temperate Asia, America, Australia, Pacific Islands and Madagascar (Chang 1951; Fang 1982; Chen and Michael 1994; Bramley 2011, 2013; Bramley et al. 2019). A total of 48 species in the genus Callicarpa were catalogued in the Flora of China (Chen and Michael 1994). Later, four new species, including C.hainanensis Z. H. Ma & D. X. Zhang (Ma and Zhang 2012), C.pararubella Qiang Wang (Wang 2019), C.stoloniformis X. X. Su, Z. H. Ma & B. Chen (Ma et al. 2023) and C.liuliana Z. H. Chen, W. Y. Xie & F. Chen (Xie et al. 2023), were reported and a new combination C.subglabra (C. Pei) L. X. Ye & B. Y. Ding was proposed (Ye et al. 2018). Thus, to date, a total of 53 Callicarpa species have been reported to be distributed in China.
For a long time, we have been puzzled by a species of Callicarpa with a quite large population cultivated in Wuhan Botanical Garden, Chinese Academy of Sciences. The species has attracted considerable attention due to features, such as procumbent branches, evergreen leaves, white fruits, scarless nodes and stellate pubescence free fruits. These features make the species distinguishable from any other previously-characterised Callicarpa species. In order to further analyse this species, we reviewed the original records and revisited its original source area in March 2023 in Yongshun, Hunan, China. Species specimens were also collected in December 2023 from Hupingshan, Shimen, Hunan, China. Careful literature review combined with comprehensive morphological and molecular characterisation obviously identified this plant as a new species in the genus Callicarpa from China.
Materials and methods
All available specimens in the genus Callicarpa from Hunan Province were retrieved from the China Virtual Herbarium (http://www.cvh.ac.cn/) and JSTOR Global Factory (https://plants.jstor.org/) and examined. Observation and morphological description of the new species was conducted, based on living plants cultivated in Wuhan Botanical Garden and individuals obtained from their type localities. The microstructure of flowers, fruits and glands were observed and photographed using a Nikon SMZ25 stereomicroscope (Nikon, Japan), while additional images were taken with a Canon 90D digital camera (Nikon, Japan). The morphological traits of the new species were described according to previous reports (Fang 1982; Chen and Michael 1994).
Sixteen Chinese Callicarpa species (Ma et al. 2023) were selected as representatives for phylogenetic analysis, with Dasymallateckiana (F. Muell.) B. J. Conn & Henwood and Dicrastylisparvifolia F. Muell. being used as outgroups. Genomic DNA was purified from the fresh plant leaves using the cetyltrimethylammonium bromide (CTAB) method (Doyle and Doyle 1987), then sequenced on an Illunima Novoseq platform in Novogene (https://cn.novogene.com/) to obtain a 5G raw data of paired end reads. The complete nuclear ribosomal sequence and complete chloroplast genome sequence was assembled using Getorganelle1.7.7.0 (Jin et al. 2020) (kmer = 127). Two nuclear DNA markers, ITS and ETS, from the nuclear ribosomal sequence and five chloroplast markers, including matK, rpl32-trnL, trnH-psbA, psbJ-petA and trnS-trnG intergenic spacer from the chloroplast genome sequence were used to construct the phylogenic relationships of the plant species. The GenBank accession numbers of sequences used in this study are listed in Table 1. All sequences were aligned with MAFFT 7.520 (Rozewicki et al. 2019) and the seven aligned DNA markers were concatenated in Phylosuite 1.2.3 (Zhang et al. 2020). The substitution model was assessed in Phylosuite 1.2.3 (Zhang et al. 2020) using ModelFinder (Kalyaanamoorthy et al. 2017). Bayesian Inference (BI) was carried out using MrBayes (Ronquist et al. 2012) in Phylosuite v.1.2.3 (Zhang et al. 2020). The number of generations run by the Markov chains was set to 2,000,000, with sampling every 1,000 generations and a burn-in of 0.25. Four Markov chains with two runs were performed. The Maximum Likelihood (ML) phylogenetic trees were inferred with IQ-TREE 2.2 (Minh et al. 2020, parameters: -m TEST, -bb 5000 and -bcor 0.90).
Taxonomic treatment
Callicarpa
yongshunensis
Taxon classificationPlantaeLamialesLamiaceae
Wen B. Xu, Xiao D. Li & Yan Ling Liu, sp nov.
A49F4F31-1BC0-5BFE-A5E9-7470BC42A8DE
urn:lsid:ipni.org:names:77340494-1
Type.
China. Hunan Province, Yongshun County, Shanmuhe Forestry Farm, growing under the forest by the edge of a valley, 29.16517°N, 109.84132°E, 676 m alt., 16 March 2023(fr.), W.B. Xu & X.W. Li 230155 (holotype: HIB, barcode no. 0250267, isotype: HIB, barcode no. 0250265).
Images of Callicarpayongshunensis Wen B. Xu, Xiao D. Li & Yan Ling Liu A inflorescence in ventral view B inflorescence in lateral view C fruit branch in lateral view D infructescence in ventral view E individual in the fruiting period of wild populations F roots developed from the node area of fruit branches G glands on the abaxial surface of the leaf H leaf, adaxial surface I leaf, abaxial surface J typical species natural habitat. Photos by Wen-Bin XU and Shu-Hui WANG.
Diagnosis.
C.yongshunensis is morphologically similar to C.luteopunctata H.T. Chang, C.giraldii Hesse ex Rehd. and C.longifolia Lamk. (Table 2), but differs from C.luteopunctatain and C.giraldii in being a procumbent shrub (vs. erect shrubs) with evergreen leaf (vs. deciduous leaf) and white fruit (vs. red or purple fruit). Similarly, unlike C.longifolia, it has procumbent shrubs (vs. erect shrubs), no transverse scar in the nodes (vs. nodes with a transverse scar) and glabrous mature fruits covered with yellow glands (vs. mature fruit covered with stellate pubescence).
The microstructure of flowers and fruits of CallicarpayongshunensisA flower B longitudinally dehiscing anther C calyx and attached glands D petals and attached glands E young fruit and attached glands F bifid stigma G petiole nodes without transverse scar. Photos by Shu-Hui WANG.
Description.
Procumbent shrub without an obvious main stem, up to 1.3 m high, branches slender, often base close to the ground and apex rising, sometimes roots growing from nodes when the nodes reaching the ground, nodes without transverse scar. Branchlets slightly 4-angled, densely covered with yellowish-brown stellate indumentum and yellow glands. Leaf with petiole 1.5–2.5 cm long, blade ovate-elliptic to oblong-elliptic, 7.5–16 × 3.5–5.5 cm, base narrowly cuneate and decurrent, margin serrate, apex acuminate, both surfaces subglabrous (with occasional hairs) and densely covered with small yellowish glands. Cymes 2–3 cm in diameter, 4–5-bifurcated, densely covered with stellate hairs and yellow glands; peduncle 4–7 mm. Calyx cup-shaped, 0.6–0.7 mm long, glabrous, sparsely yellow glandular, margin obscurely dentate and apex subtruncate. Corolla light red to light purple, 1.1–1.4 mm long, petals 4, elliptical, glabrous, slightly covered yellow glands outside. Stamens strongly exserted, usually twice as long as corolla. Style nearly as long as stamens, stigma bifid. Fruit white, 1.7–2.2 mm in diam., glabrous with sparsely yellow glands.
Distribution and habitat.
Currently, C.yongshunensis has been found in Yongshun and Shimen Counties in Hunan Province, China. It occurs in forests with weak light, at an elevation of 600–680 m. The companion species mainly include Litseaelongata (Wall. ex Nees) Benth. et Hook. f., Camelliacostei Levl., Camelliaoleifera Abel., Dichroafebrifuga Lour., Maesajaponica (Thunb.) Moritzi. ex Zoll. and Monachosorumhenryi Christ.
Phenology.
Flowers in cultivated plants at the Wuhan Botanical Garden appeared between May and July, while fruits were observed in both wild and cultivated plants at the Botanical Garden from October to March of the following year.
Etymology.
The type specimen of this species was collected from Yongshun County, Hunan Province. Thus, we chose the specific epithet “yongshunensis” for this species. Its Chinese name is “永顺紫珠”.
Conservation assessment.
The on-going field investigation only identified few populations within narrow altitude ranges. More fieldwork is still warranted to better understand this taxon. According to the guidelines of IUCN Red List Categories and Criteria (IUCN Standards and Petitions Committee 2022), C.yongshunensis is assessed as data deficient (DD).
Additional specimens examined.
C.yongshunensis (Paratypes): China. Hunan Province: Shimen County, Hupingshan National Nature Reserve, Xiahuanglian River, growing by the stream in the valley, 603 m alt., 20 December 2023 (fr.), Zhenfa Chen 231202 (HIB, barcode nos. 0250264 & 0250266, CSFI, barcode no. 080033). Hubei Province: Cultivated in Wuhan Botanical Garden, Chinese Academy of Sciences, 15 m alt., 20 June 2022 (fl.), W. B. Xu 20230302 (HIB, barcode no. 0233004).
C.luteopunctata: China. Sichuan Province: E. H. Wilson 5100 (IBSC); Emei Mountain, 10 July 1937, Chong-Shu Qian 6024 (PE); Leshan City, Mount Emei, 7 July 1941, Wen-Pei Fang 17200 (PE); Leshan City, Mount Emei, shishungou, 4 November 1952, Ji-Hua Xiong, Xiu-Shi Zhang & Xing-Lin Jiang 33464 (PE); Emei Mountain, near Hongchunping, 1000 m alt., 20 August 1957, Guang-Hui Yang 56774 (PE); Xuyong County, Shuiwei Town, Guandou Village, evergreen broad-leaved forest, 105°38'16"E, 28°16'39"N, 790 m alt., 8 June 2013, Xin-Fen Gao, Yun-Dong Gao & Wen-Bin Ju HGX11767 (CDBI); ibid., growing by the stream, under the forest, 105°23'44"E, 28°09'20"N, 14 September 2013, Wen-Bin Ju & Heng-Ning Deng HGX13448 (CDBI). Yunnan Province: Yongshan County, growing in a ravine, 1400 m alt., 13 July 1932, H. T. Tsai 51132 (LBG); Jinping County, Mengla, Tuomazhai, 900 m alt., 28 June 2009, Yunnan expedition team YN-ET400 (PE).
C.giraldii: China. Hunan Province: Yuanling County, Jiaomuxi, found at the mountain top, 1000 m alt., 23 June 1988, Wuling team 533 (IBSC); Zhijiang County, muyexi, in ravine, 90 m alt., 3 October 1988, Wuling team 1773 (IBSC); Xinning County, Ziyun Mountain, 1150 m alt., 20 October 1962, Lin-Han Liu 15240 (IBSC); Changsha, Lushan, wetland, 400–500 m alt., 2 July 1929, Shu-Zhi Xin S.H.23 (IBSC); Chengbu County, Dalao Mountain, occurring under dense forest at the mountaintop, 1500 m alt., 12 July 1959, Pei-Xiang Tan 63685 (IBSC); Sangzhi County, Shadiping, Luojiatai, valley dense forest with a slope of 20°, 1500 m alt., 22, June 1958, Lin-Han Liu 9056 (PE). Hubei Province: Yingshan County, growing near Miaoergang, subtropical coniferous broad-leaved mixed forest, roadside, 116°02'31.47"E, 30°58'41.70"N, 991 m alt., 10 August 2018, Xin-Xin Zhu et al. ZXX18584 (KUN). Jiangxi Province: Jiujiang County, mingshan, obtained under the broad-leaved forest in the valley, by the stream, 200 m alt., 3 June 1993, Ce-Ming Tan 93192 (PE). Guangdong Province: Ruyuan County, Wuzhi Mountain, Shikeng, 750 m alt., 8 August 1983, Nian-He Xia & Nian Liu 148 (IBSC).
C.longifolia; China. Guangdong Province: Zhaoqing, Dinghu Mountain, near Qingyunshi, 22 July 1979, Hua-Gu Ye 45 (IBSC); Yangshan County, Wuyuan, Yunyong Mountain, in the shade of the valley slopes, 1000 m alt., 6 June 1956, Liang Deng 1313 (PE). Hainan Province: Sanya, Ganshiling Reservoir, 17 October 1987, Ze-Xian Li 2611 (IBSC); Wanning County, xinglong, 14 May 1995, Fu-Wu Xing et al. 6717 (IBSC). Yunnan Province: Xishuangbanna Prefecture, Jinghong City, Mengyang Town, Guanping, by the edge of the forest, 900 m alt., 10 August 1977, Guo-Da Tao 17611 (HITBC); Cangyuan County, Nanla, Mangka, growing in dense forests within valleys, 1050 m alt., 6 July 1974, Yan-Hui Li 12620 (HITBC).
Molecular phylogeny
Similar topology of the Bayesian and ML trees, both displaying monophyly of the Callicarpa was observed (Fig. 3). Overall, three well-supported primary clades were identified with a posterior probability and bootstrap percentages = 1. Notably, C.yongshunensis clustered with C.luteopunctata and both morphological (Fig. 4) and molecular phylogeny evidence revealed that these two species are closely related. The resulting phylogenetic tree could separate C.yongshunensis from C.giraldii and C.longifolia, while C.giraldii clustered with C.hainanensis and C.brevipes; in addition, C.longifolia clustered with C.nudiflora. The results further confirmed previous conclusions that the phylogenetic relationship does not always reflect morphological similarity (Chang 1951). For example, C.brevipes (section Verticirima), C.giraldii (section Eucallicarpa) and C.hainanensis (section Verticirima) constituted a subclade with over 0.90 support, while C.luteopunctata (section Eucallicarpa) and C.stoloniformis (section Verticirima) were tightly clustered with C.longipes (section Eucallicarpa) in the phylogenetic tree.
Phylogenetic relationships amongst Callicarpayongshunensis and other selected species. The numbers near the nodes are Bayesian posterior probabilities and Maximum Likelihood bootstrap percentages, respectively.
Morphological comparison between C.luteopunctata (A–D) and C.yongshunensis (E–H) A, E life form showing erect shrub and procumbent shrub B, F fruit branch C, G fruit colour D, H leaf behaviour in winter showing deciduous and evergreen phenotypes (Date: 27 February 2024). Photos by Wen-Bin XU.
Supplementary Material
XML Treatment for Callicarpa yongshunensis
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bramley GLC (2011) Distribution patterns in Malesian Callicarpa (Lamiaceae).Gardens’ Bulletin (Singapore)63: 287–298.
- 2Bramley GLC (2013) The genus Callicarpa (Lamiaceae) in the Philippines.Kew Bulletin 68(3): 369–418. 10.1007/s 12225-013-9456-y · doi ↗
- 3Bramley G Bongcheewin B Davies Nde Kok RPJ Mabberley DJ Suddee S Walsingham L Wearn JS (2019) Lamiaceae. Flora Malesiana-Series 1.Spermatophyta 23: 1–424.
- 4Briquet J (1895) Verbenaceae. In: Engler A Prantl K (Eds) Die natürlichen Pflanzenfamilien IV, 3a.Wilhelm Engelmann, Leipzig, 132–182.
- 5Brown R (1810) Verbenaceae. In: Brown R Esenbeck CGNV (Eds) Prodromus Florae Novae Hollandiae et Insulae Van-Diemen.Richard Taylor & Co., London, 510–514. 10.5962/bhl.title.3678 · doi ↗
- 6Cantino PD Harley RM Wagstaff SJ (1992) Genera of Labiatae: status and classification. In: Harley RM Reynolds T (Eds) Advances in Labiatae science.Royal Botanic Gardens, Kew, 511–522.
- 7Chang HT (1951) A Review of the Chinese Species of Callicarpa.Acta Phytotaxonomica Sinica 1: 269–312.
- 8Chen SL Michael GG (1994) Verbenaceae. Flora of China Vol. 17.Science Press, Beijing and Missouri Botanical Garden Press, St. Louis, 49 pp.