Novel Ultrastructural Insights into the Clear-Cell Carcinoma of the Pancreas: A Case Report
Valentina Giansante, Luca Di Angelo, Chiara Calabrese, Paolo De Sanctis, Paolo Regi, Filippo Maria Martelli, Gianmarco Stati, Rossano Lattanzio, Saverio Alberti, Emanuela Guerra, Roberta Di Pietro

TL;DR
This case report provides new ultrastructural insights into a rare and aggressive form of pancreatic cancer called clear-cell carcinoma.
Contribution
The study reveals that disrupted mitochondria are linked to the clear-cell nature of this cancer subtype.
Findings
The tumor cells showed a clear-cell appearance with abundant intracytoplasmic vacuoles.
Transmission electron microscopy revealed a massive presence of structurally disrupted mitochondria.
Negative staining for NDUFA4L2 suggests a specific molecular characteristic of the tumor.
Abstract
Pancreatic cancer, most frequently as ductal adenocarcinoma (PDAC), is the third leading cause of cancer death. Clear-cell primary adenocarcinoma of the pancreas (CCCP) is a rare, aggressive, still poorly characterized subtype of PDAC. We report here a case of a 65-year-old male presenting with pancreatic neoplasia. A histochemical examination of the tumor showed large cells with clear and abundant intracytoplasmic vacuoles. The clear-cell foamy appearance was not related to the hyperproduction of mucins. Ultrastructural characterization with transmission electron microscopy revealed the massive presence of mitochondria in the clear-cell cytoplasm. The mitochondria showed disordered cristae and various degrees of loss of structural integrity. Immunohistochemistry staining for NADH dehydrogenase [ubiquinone] 1 alpha subcomplex, 4-like 2 (NDUFA4L2) proved specifically negative for the…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
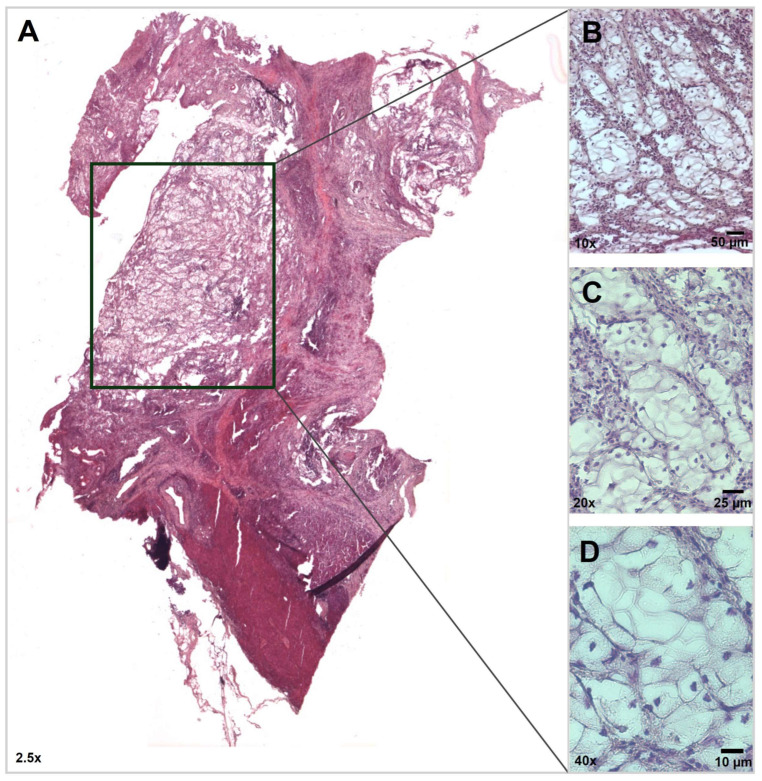 Figure 1
Figure 1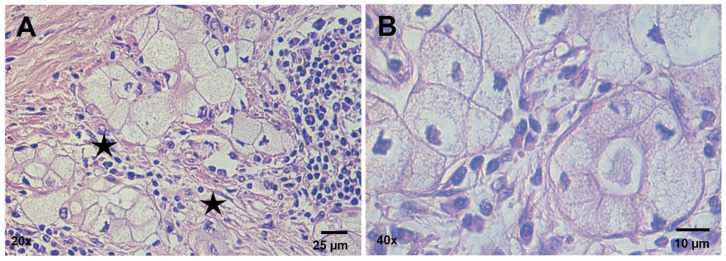 Figure 2
Figure 2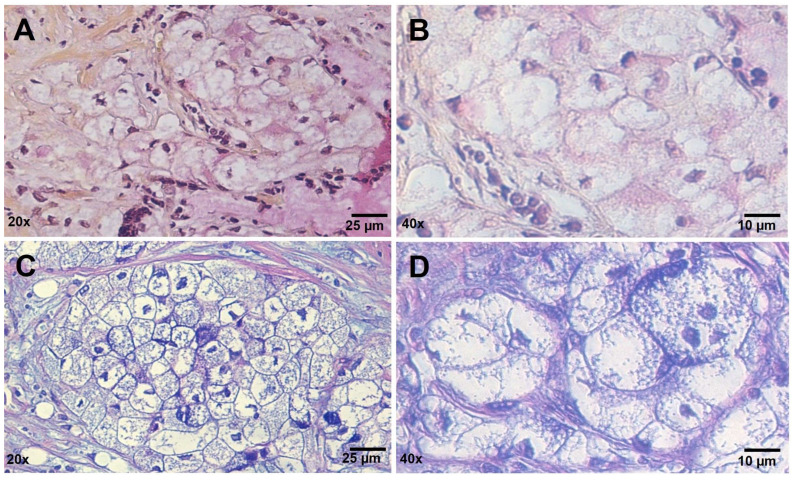 Figure 3
Figure 3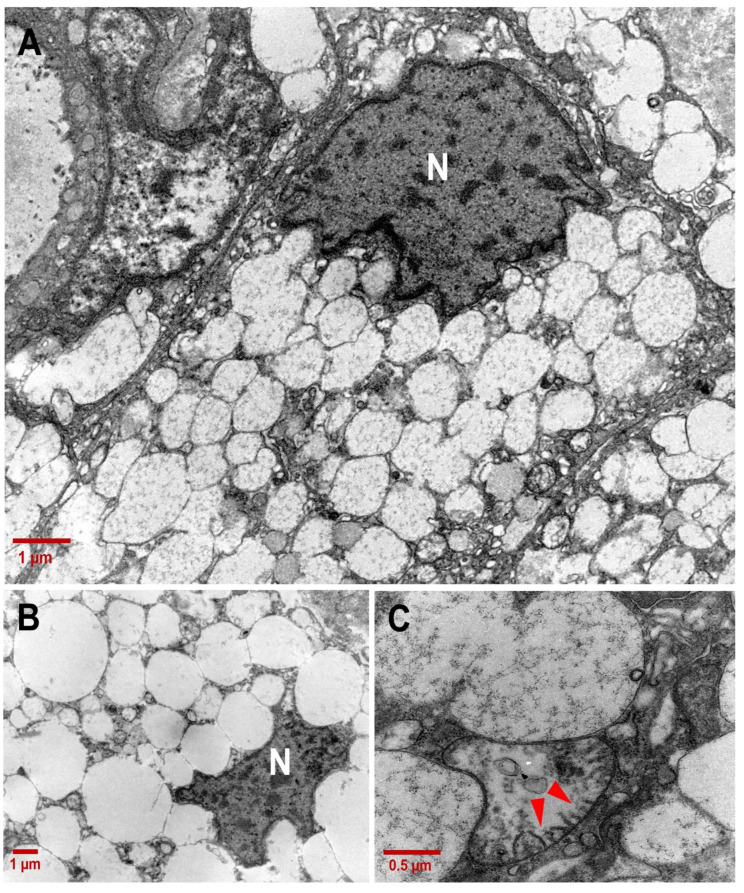 Figure 4
Figure 4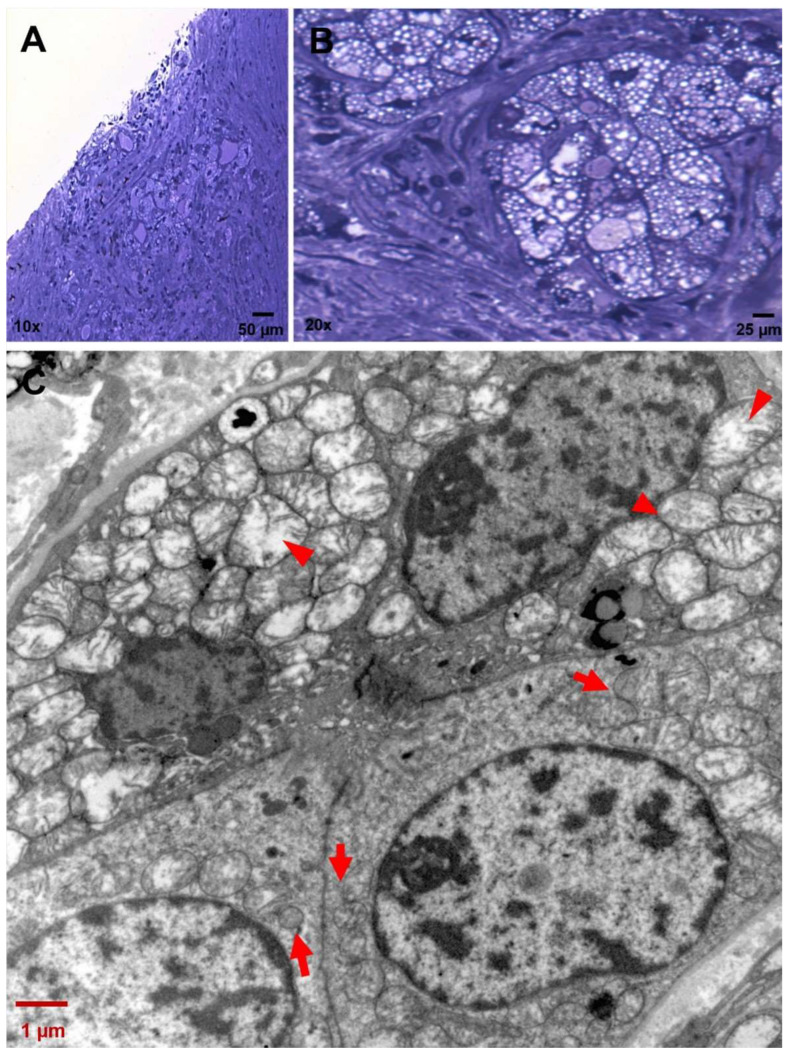 Figure 5
Figure 5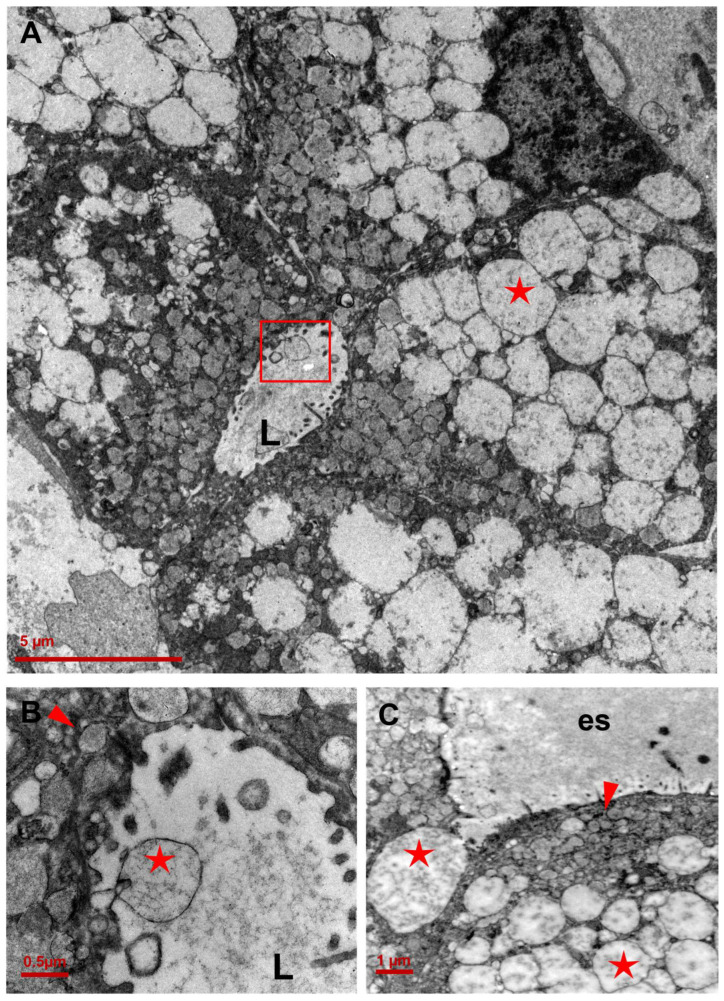 Figure 6
Figure 6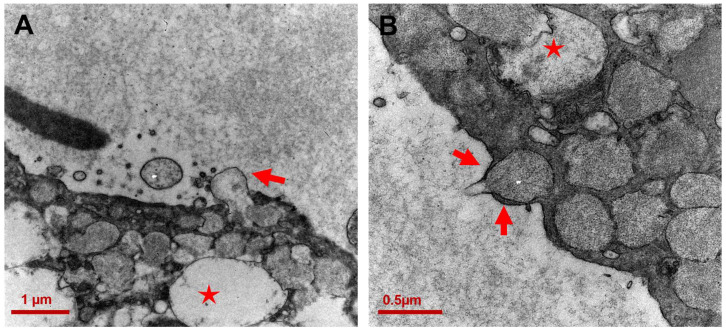 Figure 7
Figure 7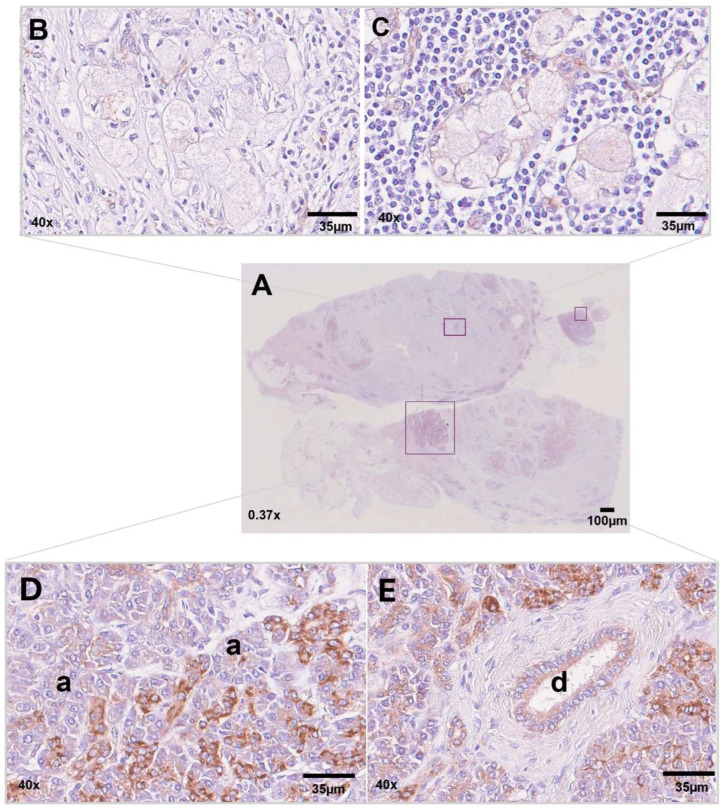 Figure 8
Figure 8- —PhD grant in Translational Medicine awarded
- —Italian Ministry of University and Research (MUR) funds
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPancreatic and Hepatic Oncology Research · Pancreatic function and diabetes · Neuroendocrine Tumor Research Advances
1. Introduction
Pancreatic cancer (PC) is rapidly becoming the third leading cause of cancer death [1], thus representing an urgent, unmet medical need.
PC aggressiveness determinants are still poorly understood. Most patients are diagnosed at an advanced stage, only 10–15% of cases being suitable for curative surgical resection. Late diagnosis, large size, and early metastasis, together with chemoresistance and aggressive biology, have contributed to PC’s poor prognosis, with an overall survival of less than 5% [2].
Cellular and matrix epithelial–mesenchymal transition (EMT) determinants [3] have been identified as risk factors. Among them is the desmoplastic reaction, a fibrotic response that surrounds cancer cells and that can act as a barrier for chemotherapeutic agents [4]. Stromal cells such as cancer-associated fibroblasts, tumor-associated macrophages, and myeloid-derived suppressor cells contribute to generating a highly immunosuppressive tumor microenvironment (TME) [5].
The KRAS mutation is the most common genetic driver in pancreatic ductal adenocarcinoma (PDAC). The mutation landscape observed in PDAC cells includes loss-of-function mutations in tumor suppressor genes, such as TP53, CDNK2A, and DPC4/SMAD4. Additional mutations are detected in the AT-rich interaction domain 1A (ARID1A), breast cancer gene (BRCA) 1, and BRCA2 [6]. Mutations in BRCA genes can account for familial PC [7].
PDAC accounts for about 90% of all PC cases [8]. Based on histopathological features, PDACs have been subcategorized into seven variants: adenosquamous carcinoma, colloid carcinoma (mucinous non-cystic carcinoma), hepatoid carcinoma, medullary carcinoma, signet-ring-cell carcinoma, undifferentiated carcinoma, and undifferentiated carcinoma with osteoclast-like giant cells [6,9]. Clear-cell carcinoma of the pancreas (CCCP) is a rare morphological pattern of ductal adenocarcinoma that is not yet fully defined. It was first observed by Cubilla et al. in 1980 [10], and only a few cases have been reported since [11]. CCCP presents with tumor cells that have clear cytoplasm and hyperchromatic nuclei. Histochemical characterization using the PAS diastase reaction and mucicarmine staining revealed heterogeneous positivity for mucins [12,13,14]. Immunohistochemistry (IHC) has shown that the clear cells are of ductal origin due to their positivity for markers like cytokeratin (CK) 7, 8, 18, and 19.
Little information is available on the cellular biology of CCCP cells. Furthermore, no study has yet been conducted to explore the ultrastructure features of these neoplastic clear cells. In the present study, we explored CCCP cell ultrastructure and metabolic pathways, to gain insight into CCCP differentiation determinants.
2. Patient Report
In 2019, a 65-year-old man was admitted to the “P. Pederzoli” Hospital in Peschiera del Garda (VR, Italy) for the onset of type 2 diabetes mellitus. Serum biochemistry showed elevated tumor markers, i.e., CEA 2.9 ng/mL (normal 0–7.0 ng/mL) and CA-19.9 15.2 U/mL (normal: 0–37.0 U/mL). Genetic testing did not detect mutated BRCA1 or BRCA2 variants. Abdominal NMR showed a pancreatic mass of 25 × 19 mm at the body–tail boundary. Distal pancreatectomy and splenectomy were performed after 24 h. Surgical pathology analysis of a 25 mm-diameter mass revealed a moderately differentiated PDAC with the presence of clear cells. A tissue sample collected from the neoformation immediately after surgery was processed for transmission electron microscopy (TEM) analysis. No post-operative adjuvant chemotherapy was administered. The patient was discharged from the hospital at day 7 after surgery and was pronounced dead in 2021.
3. Results
3.1. Histopathology Findings
An overview of the histochemical characterization of the patient’s tissue sample is shown in Figure 1, with a special focus on the tumor areas (Figure 1B–D). Upon examination of the Hematoxylin and Eosin (H&E) staining of the formalin-fixed, paraffin-embedded (FFPE) pancreatic tumor tissue, neoplastic cells exhibited a prominent, clear-cell feature arranged to form nest-like structures immersed in a desmoplastic stroma (Figure 2A,B). According to the diagnostic criteria of Kim et al. [13], these findings were interpreted as a transitional trend of cancer cells towards the loss of differentiation.
The tumor cells were large, round to oval, and appeared packed with abundant intracytoplasmic vacuoles that provided a characteristic foamy appearance. Nuclei were centrally or peripherally placed and were often hyperchromatic or pleomorphic with irregular borders.
To investigate whether the clear-cell foamy appearance was due to an excess of mucins, mucicarmine and Alcian–PAS stainings were performed (Figure 3). Upon staining, the clear tumor cells showed heterogeneity, with some completely negative and others weakly stained, indicating the scant presence of acid mucin, if any. However, weakly stained cells showed a peculiar texture within the cytoplasm, also seen in previously described case reports but never thoroughly investigated [13].
Thus, to gain insight into the ultrastructure underlying this abundant clear cytoplasm that was not due to the accumulation of mucin vacuoles as in foamy gland adenocarcinomas, we went on to perform TEM and metabolic analysis of CCCP cells.
3.2. Ultrastructure of CCCP Cells
The TEM analysis allowed us to to gather insight into the CCCP cytoplasmic organelle and vesicle distribution and content. While several intracytoplasmic vacuoles appeared devoid of detectable content, several others were characterized by poorly defined tubular structures that resembled mitochondrial cristae (Figure 4). The electron-lucent matrix combined with the irregular conformation of the cristae found inside these vacuoles indicated that they were mitochondria undergoing disruption. Consistent with this, cristae within the vacuoles appeared fragmented, suggesting cristolysis, as frequently observed during mitochondria swelling.
The sequential steps of mitochondria degeneration were observed side by side in CCCP cells, suggesting sequential processes of progressive degeneration. In Figure 5B, four cells are shown with an acinar aspect and containing mitochondria at different morphological stages, from healthy mitochondria with well-preserved cristae to unhealthy mitochondria with completely disrupted cristae.
The diverse dimension, morphology, and electron density between subplasmalemmal vacuoles containing mucin and dysfunctional mitochondria is displayed in Figure 6. Unexpectedly, clear vacuoles were also found in the lumen of transformed acini (Figure 6B).
Consistent with this finding, TEM captured membrane events (an example is shown in Figure 7A) where the cell seems to exocytose the cytoplasmic electron-lucent vacuole across the membrane into the extracellular space. Mucin-containing electron-dense vacuoles were also seen crossing the cell membrane (Figure 7B).
3.3. Immunohistochemistry for NDUFA4L2
Based on our ultrastructural evaluation, intracytoplasmic vacuoles were indicated to correspond to unfunctional mitochondria with broken cristae. NADH dehydrogenase [ubiquinone] 1 alpha subcomplex, 4-like 2 (NDUFA4L2), is a subunit of Complex I of the mitochondrial respiratory chain, which is highly expressed in clear-cell renal carcinoma and involved in the transfer of electrons from NADH to ubiquinone and in the regulation of autophagic turnover of damaged mitochondria [15]. The IHC investigation of NDUFA4L2 expression in the CCCP tissue sample (Figure 8A) showed a weak labeling signal shaping the intracellular texture in a few neoplastic cells (Figure 8B,C), whereas most clear cells did not express NDUFA4L2. In contrast, NDUFA4L2 labeling was detected in non-neoplastic acinar and ductal epithelial cells (Figure 8D,E).
4. Discussion
Although data on incidence and follow-up are currently limited, previous reports have described CCCP as an aggressive histological variant of PDAC, associated with poor patient outcomes [11]. In this report, we applied ultrastructural TEM analysis to a CCCP surgical specimen from a 65-year-old patient. Our results show for the first time that the optical “clear cells” or foamy-cell variants of PDAC display ultrastructural hallmarks of altered mitochondria. Indeed, we discovered that the clear vacuoles within tumor cells correspond to swollen electron-lucent mitochondria, in the process of losing structure integrity and containing cristae remnants. These observations are unprecedented. It is known that several factors can lead to mitochondrial swelling, including oxidative and/or energetic stress such as free radical production, lipidic peroxidation, and hypoxia [16,17]. Mitochondrial swelling has been linked to the oxidation of the complexes of the oxidative chain and is correlated, together with mitochondrial crowding, with the accumulation of misfolded proteins inside mitochondria and the endoplasmic reticulum, causing the oxidation of mitochondrial proteases and chaperones [18].
Mitochondrial swelling and cytoplasmic vacuolization are also important signs of paraptosis, a non-apoptotic form of regulated cell death (RCD) first described by Sperandio et al. [19]. There is evidence that paraptosis is triggered when neoplastic cells are treated with specific compounds [20], but the real role of this alternative cell death in cancer has yet to be defined. The ultrastructural similarities found in our case suggest a link with paraptosis. This is not so unusual for cancer cells, considering that other forms of RCD, such as necroptosis, are emerging as strategies for tumor cell survival and growth, especially in PDAC [4].
Electron-lucent vacuoles were present inside the cells and in the lumen of neoplastic acini (Figure 6). The frequency of transitions across the plasma membrane suggests vacuole exchange between cells (Figure 6 and Figure 7). Under physiological conditions, mitochondrial transfer across cells is a common mechanism triggered by cells to maintain tissue homeostasis. It generally refers to the effective horizontal transport of functioning mtDNA copies or of entire mitochondria from donor cells, to restore the normal activity in the recipient hosts [21].
Both healthy and damaged mitochondria can be transferred among cells [22]. For example, within the bone marrow stem cell niche, the low oxygen environment induces mesenchymal stem cells (MSCs) to package partially depolarized mitochondria into larger vesicles (>100 nm), which are then accepted by surrounding macrophages [23]. This phenomenon is known as “trans-mitophagy” and allows the cell to “commission” the degradation of unfunctional organelles to recipient cells, after releasing them in the extracellular space. It is worth noting that the functionality of the mitochondria is irrelevant for their transfer, while intact mtDNA is indispensable for mitochondria to be transferred to another cell to recover the respiratory capacity of mitochondria-deficient cancer cells [24].
There are different modalities to “shuttle” mitochondria. Neighboring cells can connect each other by forming ultrafine cytoplasmic bridges called tunneling nanotubes (TNTs). Alternatively, they can establish a distance connection through the release of extracellular vesicles (EVs) containing the mitochondrial cargo, as was observed in our case. EVs are critical mediators of organized communities of cells, involved in several pathways such as differentiation, migration and immunoregulation [25]. In this case, we propose that CCCP could dispose of unfunctional mitochondria through a mechanism of trans-mitophagy, becoming a potential source of extracellular vesicles. The reason for this release is unknown, but given the main role of TME in PDAC, it could be an additional means of communication between the tumor and surrounding cells.
NDUFA4L2 mediates the function of oxidative phosphorylation, so it is critical for mitochondrial activity [26]. Loss of NDUFA4L2 through gene silencing leads to the inhibition of the autophagic machine and increased mitochondrial mass [15]. The hypothesis that clear cells could contain unfunctional mitochondria undergoing mitophagy is supported by the general absence of NDUFA4L2 in CCCP cells, except for sparsely positive groups. The opposite situation is seen within non-tumoral pancreatic structures that are significantly labeled by the anti-NDUFA4L2 antibody and might explain the different functional states of the mitochondria between normal and neoplastic cells.
Mitochondrial reprogramming is critical for tumorigenesis [27]. Hence, our findings suggest a link between mitochondrial disfunction and CCCP tumor progression.
5. Materials and Methods
5.1. Light Microscopy
Human samples of primary pancreatic ductal adenocarcinoma were collected immediately after surgical excision and fixed with 4% formaldehyde in 1× phosphate-buffered saline (PBS) for 24 h. After dehydration in graded alcohols (from 70% to 100% ethanol), samples were embedded in paraffin, and cut on a microtome (Leica, RM 2265, Nussloch, Germany). Sections were stained with Mayer’s hemallum (Bio-Optica, Milan, Italy) and 1% Eosin Y aqueous solution (Bio-Optica), Alcian Blue pH 2.5–PAS. (Bio-Optica), or Mucicarmine Mayer (Bio-Optica), according to the manufacturer’s instructions. MetaMorph 6.1 Software System (Universal imaging Corp, Molecular Device Corp, San Jose, CA, USA) with 2D reconstruction was employed to acquire digital images.
5.2. Immunohistochemistry (IHC)
The expression of NDUFA4L2 was evaluated with IHC on formalin-fixed, paraffin-embedded tissue (FFPE) sections. Tissue sections were incubated with a 1:80 dilution of anti-NDUFA4L2 rabbit polyclonal antibody (Proteintech, Chicago, IL, USA) for 90 min, followed by incubation with HRP-conjugated goat anti-rabbit secondary antibody (Thermo Fisher Scientific, Waltham, MA, USA) according to the manufacturer’s instructions. A validated skin tissue microarray was used as a staining control. Whole-slide images were acquired with NanoZoomerXR, Hamamatsu.
5.3. Transmission Electron Microscopy (TEM)
Following the protocol of Basile at al. [28], pancreatic ductal adenocarcinoma samples from surgical excision were fixed with 2.5% glutaraldehyde (Electron Microscopy Sciences, Hatfield, PA, USA) in 0.1 M cacodylate buffer (Electron Microscopy Sciences) (pH 7.2–7.4) for 2 h at 4 °C and post-fixed in 1% osmium tetroxide (Electron Microscopy Sciences) for 1 h at 4 °C. After a washing cycle in cacodylate buffer, samples were dehydrated in graded alcohols (from 50% to 100% ethanol) and embedded in Spurr resin (Electron Microscopy Sciences). Ultrathin sections (70 nm) were obtained by using a Reichert ultramicrotome (Reichert, Inc, Teramo, Italy) mounted on 200-mesh copper grids (Electron Microscopy Sciences) and counterstained with UranyLess and lead citrate staining solutions (Electron Microscopy Sciences) for 10–15 min/each. Samples were observed under a ZEISS EM109 electron microscope equipped with a Gatan 830Z00W44 camera (Gatan GmbH, Ingol-stadterstr. 12 D-80807 München, Germany). Digital Micrograph software version 2.02.800.0 was used for acquiring and processing digital image data [28].
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Siegel R.L. Miller K.D. Wagle N.S. Jemal A. Cancer statistics, 2023 CA Cancer J. Clin.202373174810.3322/caac.2176336633525 · doi ↗ · pubmed ↗
- 2Park W. Chawla A. O’Reilly E.M. Pancreatic Cancer: A Review JAMA 202132685186210.1001/jama.2021.1302734547082 PMC 9363152 · doi ↗ · pubmed ↗
- 3Krebs A.M. Mitschke J. Lasierra Losada M. Schmalhofer O. Boerries M. Busch H. Boettcher M. Mougiakakos D. Reichardt W. Bronsert P. The EMT-activator Zeb 1 is a key factor for cell plasticity and promotes metastasis in pancreatic cancer Nat. Cell Biol.20171951852910.1038/ncb 351328414315 · doi ↗ · pubmed ↗
- 4Giansante V. Stati G. Sancilio S. Guerra E. Alberti S. Di Pietro R. The Dual Role of Necroptosis in Pancreatic Ductal Adenocarcinoma Int. J. Mol. Sci.2023241263310.3390/ijms 24161263337628814 PMC 10454309 · doi ↗ · pubmed ↗
- 5Truong L.H. Pauklin S. Pancreatic Cancer Microenvironment and Cellular Composition: Current Understandings and Therapeutic Approaches Cancers 202113502810.3390/cancers 1319502834638513 PMC 8507722 · doi ↗ · pubmed ↗
- 6Bazzichetto C. Luchini C. Conciatori F. Vaccaro V. Di Cello I. Mattiolo P. Falcone I. Ferretti G. Scarpa A. Cognetti F. Morphologic and Molecular Landscape of Pancreatic Cancer Variants as the Basis of New Therapeutic Strategies for Precision Oncology Int. J. Mol. Sci.202021884110.3390/ijms 2122884133266496 PMC 7700259 · doi ↗ · pubmed ↗
- 7Hahn S.A. Greenhalf B. Ellis I. Sina-Frey M. Rieder H. Korte B. Gerdes B. Kress R. Ziegler A. Raeburn J.A. BRCA 2 germline mutations in familial pancreatic carcinoma J. Natl. Cancer Inst.20039521422110.1093/jnci/95.3.21412569143 · doi ↗ · pubmed ↗
- 8Conroy T. Pfeiffer P. Vilgrain V. Lamarca A. Seufferlein T. O’Reilly E.M. Hackert T. Golan T. Prager G. Haustermans K. Pancreatic cancer: ESMO Clinical Practice Guideline for diagnosis, treatment and follow-up Ann. Oncol.202334987100210.1016/j.annonc.2023.08.00937678671 · doi ↗ · pubmed ↗