Relationship between 233 colorectal cancer risk loci and survival in 1926 patients with advanced disease
Christopher Wills, Amy Houseman, Katie Watts, Timothy S. Maughan, David Fisher, Richard S. Houlston, Hannah D. West, Valentina Escott-Price, Jeremy P. Cheadle

TL;DR
This study found two genetic variants linked to survival outcomes in patients with advanced colorectal cancer, suggesting potential new biomarkers for prognosis.
Contribution
Identified two SNPs (rs117079142 and rs9924886) significantly associated with survival in advanced colorectal cancer patients.
Findings
rs117079142 near UTP23 and EIF3H was strongly linked to worse survival in a recessive model.
rs9924886 near CDH1 and CDH3 was associated with increased risk of poor survival.
Lower CDH1 expression in tumors correlated with worse survival outcomes.
Abstract
Genome, transcriptome and methylome-wide association studies have identified single-nucleotide polymorphisms (SNPs) or genes at 258 loci associated with colorectal cancer (CRC) risk. We studied the relationship between these and patient outcome. We studied 1926 unrelated patients with advanced CRC from COIN and COIN-B. Of 205 CRC-risk SNPs, 19 were directly genotyped and 162 were imputed, and of 53 risk genes, 52 were tested. An additive Cox model for overall survival was adjusted for known prognostic factors. For nominally significant SNPs or genes, we considered a recessive model with a Bonferroni corrected threshold of P = 2.1 × 10−4. We examined SNPs as expression quantitative trait loci (eQTL) and the relationship between gene expression in colorectal tumours and survival in 597 unrelated patients. Eleven SNPs or genes were nominally associated with survival under an additive…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
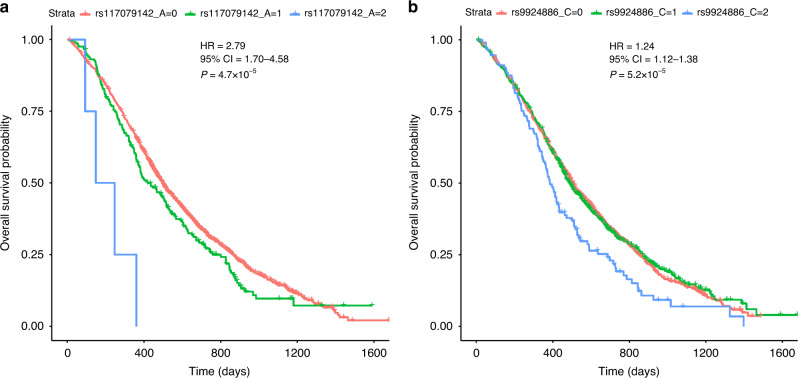 Figure 1
Figure 1- —Tenovus Cancer Care
- —http://dx.doi.org/10.13039/100009828Cancer Research Wales
- —Cardiff University School of Medicine
- —UK Medical Research Council
- —Cancer Research UK
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCancer-related molecular mechanisms research · Genetic Associations and Epidemiology · RNA modifications and cancer
Introduction
Genome-wide association studies (GWAS) have identified single-nucleotide polymorphisms (SNPs) associated with risk of developing colorectal cancer (CRC) [1]. Some studies have suggested that a subset of these may also influence patient survival [2–7] although other studies have not supported these observations [8–11]. We previously studied the relationship between SNP genotype and patient outcome for 83 CRC-risk SNPs [12] by analysing patients with advanced CRC from the COIN and COIN-B clinical trials [13, 14]. A recent meta-analysis of all available GWAS augmented by transcriptome and methylome-wide association studies (TWAS and MWAS, respectively) has identified further loci taking the total number of CRC-risk loci to 258 [15].
To gain a more comprehensive understanding of the relationship between inherited genetic variation and patient survival, we assessed 233 of these risk loci for their prognostic role in 1926 patients from COIN and COIN-B.
Materials and methods
Patients and genotyping
Germline DNAs were extracted from EDTA venous blood samples from 2244 unrelated patients with metastatic or locally advanced colorectal adenocarcinoma participating in the MRC clinical trials COIN (NCT00182715) [13] and COIN-B (NCT00640081) [14]. All patients gave fully informed consent for bowel cancer research (approved by NHS Research Ethics Committee [04/MRE06/60]). COIN patients were randomised 1:1:1 to receive continuous oxaliplatin and fluoropyrimidine chemotherapy, continuous chemotherapy and cetuximab, or intermittent chemotherapy. COIN-B patients were randomised 1:1 to receive intermittent chemotherapy and cetuximab, or intermittent chemotherapy and continuous cetuximab. There was no heterogeneity in overall survival (OS; time from trial randomisation to death or end of trial) between patients when analysed by trial, trial arm, type of chemotherapy received, or cetuximab use [12], so we combined groups for survival analyses. Patient DNAs were genotyped using Affymetrix Axiom Genome-Wide CEU 1 Human Mapping Arrays [16].
Prediction of untyped SNPs was carried out using IMPUTE2 v2.3.0 [17] based on data from the 1000 Genomes Project as reference [18, 19]. After quality control (QC), SNP genotypes were available on 1950 patients. Two patients had no data on survival and a further 22 lacked clinicopathological data leaving 1926 for analysis (of which 1435 died at censorship).
SNPs and genes analysed
For the 205 CRC-risk SNPs, 19 were directly genotyped, 162 were imputed and 24 were not analysed (one because it was on the X-chromosome which was not genotyped, 19 had INFO scores <0.7 and 4 had minor allele frequencies [MAFs] <0.01). Therefore, in total, 181 CRC-risk SNPs were tested for an association with OS.
For the CRC-risk genes identified from TWAS and MWAS, we used data from a GWAS of COIN and COIN-B (2.9 million SNPs post-QC; [16]). SNPs were mapped to a region spanning 35 kilobases before and 10 kilobases after the transcription zone and analysed using MAGMA v1.07b [20]. Of the 53 genes, 52 were successfully analysed (one had insufficient SNPs in their annotation window).
Statistical analysis
The relationship between genotype and OS was determined using an additive Cox survival model adjusting for 11 prognostic covariates previously identified in COIN and COIN-B: WHO performance status (P = 3.1 × 10^−23^), resection status of the primary tumour (P = 1.8 × 10^−21^), WBC count (P = 1.2 × 10^−31^), platelet count (P = 1.7 × 10^−29^), alkaline phosphatase levels (P = 1.5 × 10^−27^), number of metastatic sites (P = 1.7 × 10^−13^), liver metastases (P = 1.3 × 10^−4^), site of primary tumour (P = 9.1 × 10^−9^), surface area of primary tumour (P = 1.1 × 10^−5^), time from diagnosis to metastases (P = 1.7 × 10^−7^), and metachronous versus synchronous metastases (P = 6.0 × 10^−8^) [21]. For gene level analysis in MAGMA, SNP P-values were assessed with the linkage disequilibrium (LD) between them using the multi=snp-wise option. This model takes advantage of the sum of the -log_10_(P) for all SNPs, as well as the top SNP associations within each gene, to assess the association of their constituent genes. For any SNPs or genes nominally associated with OS (P < 0.05), we also considered a recessive model to uncover associations potentially missed under additive analyses [22]. We used Bonferroni correction to address multiple testing with P < 2.1 × 10^−4^ being considered statistically significant (0.05/233 SNPs or genes tested). Based on the number of patients analysed, our analysis provided over 70% power to demonstrate a HR of 1.2 for SNPs with MAFs >0.30. Power was calculated using the ‘survSNP.power.table’ function from the ‘survSNP’ package in R [23].
Bioinformatic analyses
We queried the GTEx [24] database to examine SNPs as potential expression quantitative trait loci (eQTLs) for neighbouring genes. Significance for tissue association was set at P < 1.0 × 10^-3^ (Bonferroni correction for 49 tissues [0.05/49]). We correlated gene expression with survival by analysing tumours from 597 patients with CRC from The Human Protein Atlas (THPA) [25]. RNA-seq data was reported as median number of fragments per kilobase of exon per million reads (FPKM) [26]. Samples were classified as high expression using the thresholds recommended by THPA (for CDH1 FPKM was >137; https://www.proteinatlas.org/ENSG00000039068-CDH1/pathology/colorectal+cancer). A log-rank P-value was obtained for a difference in survival between patients with CRCs with high and low expression levels. We also performed survival analysis using a linear Cox-proportional hazards model.
Results
In total, we had survival, clinicopathological and germline genotyping data on 1926 patients with advanced CRC (Table 1). We found that eight CRC-risk SNPs (rs13086367 at 3q13.2, rs280097 at 4q22.2, rs16892766 at 8q23.3, rs117079142 at 8q24.11, rs11255841 at 10p14, rs4444073 at 11p15.4, rs1497077 at 14q22.1 and rs9924886 at 16q22.1) and three CRC-risk genes (EPB41L2, ADAMTS15 and F2), were nominally associated with survival under an additive model (Table 2, Supplementary Table 1).Table 1. Clinicopathological features of patients with advanced colorectal cancer.Clinicopathological factorPatients with advanced CRC(n = 1926)n%SexMale126165.5Female66534.5AgeMedian (years)64-Overall survivalMedian494-(95% CI) (days)(469–514)WHO performance status090046.7188546.021417.3Site of primary tumourLeft colon49325.6Right colon51426.7Rectosigmoid junction28314.7Rectum60931.6Unknown colon60.3Multiple sites211.1Status of primary tumourResected102253.1Unresected90446.9Stage100.0200.0300.041926100.0Timing of metastasesMetachronous57529.9Synchronous135170.1Type of metastasesLiver only42622.1Liver + others101952.9Non-liver^a^47924.9None20.1Number of metastatic sites169035.8275839.4≥347824.8Data are n (%) or median.^a^Non-liver metatases included those in the lungs, peritoneum and lymph nodes.Table 2CRC-risk SNPs or genes associated with survival.SNP/GeneLocusMinor alleleGenesAdditive modelRecessive modelHR95% CIPHR95% CIPrs1125584110p14ARNA5SP2990.880.81–0.951.7 × 10^−3^0.850.77–0.942.0 × 10^−3^rs168927668q23.3CEIF3H, LOC1053757131.21.06–1.364.0 × 10^−3^1.280.95–1.730.1rs1170791428q24.11AEIF3H, UTP231.261.07–1.486.0 × 10^−3^2.791.70–4.584.7 × 10^−5^rs444407311p15.4CCAND1.11, ADM, LOC653503, SBF21.111.03–1.207.0 × 10^−3^1.010.94–1.070.87rs992488616q22.1CCDH3, CDH1, HSPE1P5, RNA5SP4291.121.03–1.231.1 × 10^−2^1.241.12–1.385.2 × 10^−5^rs2800974q22.2C1.11.02–1.191.4 × 10^−2^1.081.01–1.153.5 × 10^−2^rs149707714q22.1TNID2, RTRAF1.11.02–1.191.7 × 10^−2^1.121.03–1.217.2 × 10^−3^rs130863673q13.2GBOC, LINC020440.920.86–1.004.4 × 10^−2^0.950.89–1.010.12EPB41L26q23.2----2.6 × 10^−3^---ADAMTS1511q24.3----1.7 × 10^−2^---F211p11.2----3.2 × 10^−2^---Risk SNPs or genes nominally associated with survival (P < 0.05) under an additive model. Genes annotated within a region spanning 50 kb up or downstream of the SNP. Only rs117079142 and rs9924886 passed the threshold for multiple testing (P < 2.1 × 10^−4^) when considered under a recessive model (in bold).
Only rs117079142 (MAF = 0.06, HR = 2.79, 95% CI = 1.70–4.58, P = 4.7 × 10^−5^) and rs9924886 (MAF = 0.25, HR = 1.24, 95% CI = 1.12–1.38, P = 5.2 × 10^−5^) passed the threshold for multiple testing when considered under a recessive model (Table 2). Patients homozygous for the rs117079142 minor allele (n = 4) had a median survival of 198 days compared to 420 days for heterozygotes (n = 204) and 497 days for patients homozygous for the major allele (n = 1724) (Fig. 1). Patients homozygous for the rs9924886 minor allele (n = 113) had a median survival of 385 days compared to 487 days for heterozygotes (n = 715) and 507 days for patients homozygous for the major allele (n = 1026) (Fig. 1).Fig. 1. Relationship between rs117079142 and rs9924886 genotype and overall survival.Kaplan–Meier Plots for a rs117079142 and b rs9924886. P-values are for multivariate recessive Cox-regression models and patients are grouped by number of copies of the minor allele. The relationship between genotype and overall survival was adjusted for eleven prognostic covariates: WHO performance status, resection status of the primary tumour, white blood cell count, platelet count, alkaline phosphatase levels, number of metastatic sites, liver metastases, site of primary tumour, surface area of primary tumour, time from diagnosis to metastases and metachronous versus synchronous metastases.
rs117079142 was an eQTL for UTP23 (Supplementary Table 2) and rs9924886 was an eQTL for CDH1, CDH3 and ZFP90 (Supplementary Table 2) in multiple tissues, but neither were significant in the sigmoid or transverse colon. Low CDH1 expression in CRCs was associated with worse survival in patients from THPA (5-year survival: low CDH1 expression = 58%, high CDH1 expression = 71%, HR = 2.18, 95% CI = 1.3–3.5, P = 1.8 × 10^−3^; linear Cox-proportional hazards model P = 2.8 × 10^−2^). UTP23, EIF3H and CDH3 expression levels were not associated with survival.
Discussion
In this study, we investigated the relationship between CRC-risk variants and patient outcome. We identified two SNPs associated with survival under a recessive model that were significant beyond the threshold for multiple testing. Interestingly, both SNPs were only nominally significant under additive analyses and others have previously reported on the value of considering recessive models to uncover associations potentially missed [22]. rs117079142 had a modest effect size (HR = 2.79), but relatively low frequency in our cohort; furthermore, in the 1000 genomes dataset the MAF ranges from 0.0076 in the African population to 0.073 in South Asians. In contrast, rs9924886 was more commonly observed in our cohort (and was 0.178 in the African population and 0.3095 in East Asians), but the effect size was lower. These data suggest that neither SNPs are likely to have a direct clinical impact although their identification helps inform potential therapeutic targets.
rs117079142 lies 4 kb downstream of UTP23. UTP23 codes for part of the 90S pre-ribosome and is required for 18S rRNA early processing. Reduced UTP23 expression has been associated with poor prognosis in patients with ovarian cancer possibly by affecting sensitivity to paclitaxel-based chemotherapy [27]. rs117079142 also lies 23 kb downstream of EIF3H, which regulates translation through its interaction with the 40S ribosome and other initiation factors. EIF3 subunits are thought to have oncogenic potential [28] through increased protein synthesis of oncoproteins such as cyclinD1, c-Myc, FGF2 and ornithine decarboxylase [29].
rs9924886 in CDH3 is a strong proxy for rs9929218 (D’ = 0.95 and r^2^ = 0.80) and rs9939049 (D’ = 0.96 and r^2^ = 0.80) in CDH1 (encoding E-cadherin) that we previously identified as a prognostic biomarker in CRC [12, 30]. Others have also demonstrated a relationship between rs9929218 and survival in CRC patients from Korea [31] and Spain [5]. rs9924886, rs9929218 and rs9939049 are in strong LD with rs16260 [32] in the CDH1 promoter, which down-regulates CDH1 expression [33]. Patients homozygous for the minor alleles of these variants would be expected to have reduced E-cadherin expression. Mechanistically, our data are consistent with the downregulation of CDH1 affecting survival. First, we found that patients homozygous for the rs9924886 minor allele had worse survival and second, we observed that patients with low CDH1 expression in their colorectal tumours had worse outcome. E-cadherin functions as a transmembrane glycoprotein involved in intercellular adhesion, cell polarity and tissue morphology and regeneration [34], and its loss is a key feature of epithelial to mesenchymal transition during metastasis. Together, these data support a prognostic role for CDH1 in colorectal tumourigenesis.
rs10161980 has been previously associated with survival from CRC under a recessive model [22]. However, we failed to replicate this SNP in COIN and COIN-B despite having over 98% power. rs10161980 may therefore represent a false-positive or a prognostic biomarker that is specific to patients with earlier stages of disease (we only considered patients with advanced disease in our analyses).
In conclusion, our work provides support for the importance of germline variation as a determinant of patient outcome. Understanding the biological basis of these relationships provides a focus for future work with the goal of identifying novel therapeutic targets for the treatment of CRC.
Supplementary information
Supplementary Information
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Law PJ Timofeeva M Fernandez-Rozadilla C Broderick P Studd J Fernandez-Tajes J Association analyses identify 31 new risk loci for colorectal cancer susceptibility Nat Commun 201910215410.1038/s 41467-019-09775-w 31089142 PMC 6517433 · doi ↗ · pubmed ↗
- 2Phipps AI Newcomb PA Garcia-Albeniz X Hutter CM White E Fuchs CS Association between colorectal cancer susceptibility loci and survival time after diagnosis with colorectal cancer Gastroenterology 201214351 U 55510.1053/j.gastro.2012.04.05222580541 PMC 3579620 · doi ↗ · pubmed ↗
- 3Dai JY Gu J Huang MS Eng C Kopetz ES Ellis LMGWAS-identified colorectal cancer susceptibility loci associated with clinical outcomes Carcinogenesis 20123313273110.1093/carcin/bgs 14722505654 PMC 4072910 · doi ↗ · pubmed ↗
- 4Garcia-Albeniz X Nan H Valeri L Morikawa T Kuchiba A Phipps AI Phenotypic and tumor molecular characterization of colorectal cancer in relation to a susceptibility SMAD 7 variant associated with survival Carcinogenesis 201334292810.1093/carcin/bgs 33523104301 PMC 3564438 · doi ↗ · pubmed ↗
- 5Abuli A Lozano JJ Rodriguez-Soler M Jover R Bessa X Munoz J Genetic susceptibility variants associated with colorectal cancer prognosis Carcinogenesis 20133422869110.1093/carcin/bgt 17923712746 · doi ↗ · pubmed ↗
- 6Takatsuno Y Mimori K Yamamoto K Sato T Niida A Inoue H The rs 6983267 SNP is associated with MYC transcription efficiency, which promotes progression and worsens prognosis of colorectal cancer Ann Surg Oncol 201320139540210.1245/s 10434-012-2657-z 22976378 · doi ↗ · pubmed ↗
- 7Morris EJA Penegar S Whiffin N Broderick P Bishop DT Northwood EA retrospective observational study of the relationship between single nucleotide polymorphisms associated with the risk of developing colorectal cancer and survival P Lo S ONE 2015101110.1371/journal.pone.0117816 PMC 433973125710502 · doi ↗ · pubmed ↗
- 8Tenesa A Theodoratou E Din FV Farrington SM Cetnarskyj R Barnetson RA Ten common genetic variants associated with colorectal cancer risk are not associated with survival after diagnosis Clin Cancer Res 2010163754910.1158/1078-0432.Ccr-10-043920628028 PMC 2906703 · doi ↗ · pubmed ↗