Generation of single-cysteine E. coli ProQ variants to study RNA-protein interaction mechanisms
Helen S. Washington, Shiying Wang, Katherine E. Berry

TL;DR
Researchers created modified versions of the ProQ protein in E. coli to better understand how it interacts with RNA.
Contribution
A set of 13 single-cysteine ProQ variants was developed for studying RNA-protein interaction mechanisms.
Findings
The variants retain RNA-binding ability while having single-cysteine residues at different positions.
These variants will help map RNA orientation around ProQ for biochemical and biophysical studies.
Abstract
ProQ is a FinO-domain protein found in E. coli and other proteobacteria that has a global RNA-binding profile. In order to probe the detailed mechanism of RNA interactions, we have developed a collection of 13 E. coli ProQ variants that possess single-cysteine residues at varied positions on the surface of the N-terminal FinO domain and retain the ability to bind well to RNA. This set of variant ProQ proteins will support future biochemical and biophysical studies to map the orientation of bound RNAs to different sites around the ProQ protein, shedding light on the mechanism of ProQ-RNA interactions.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
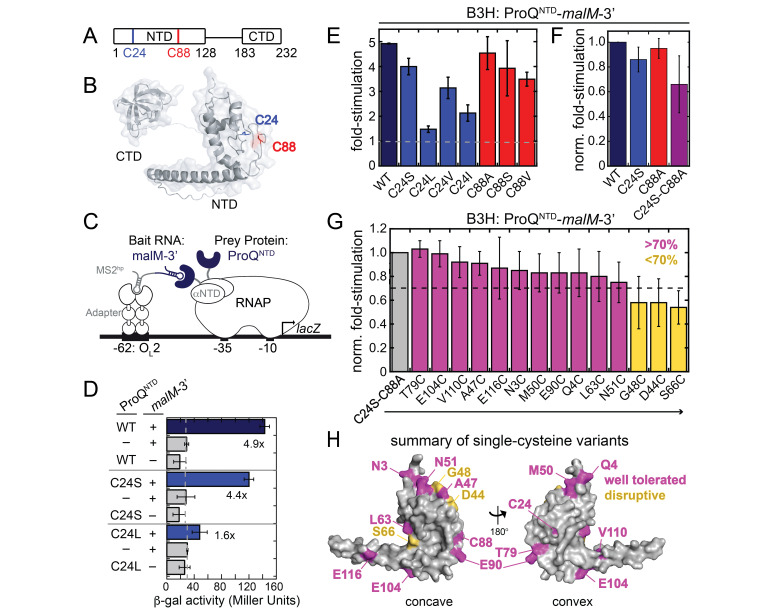 Figure 1
Figure 1|
|
|
|
|
|
pAClCI |
empty vector |
Encodes full-length lCI under the control of the
|
|
|
pBrα |
Empty vector |
Encodes full-length
|
|
|
pCH1 |
pCDF-1XMS2hp (empty vector) |
pCDF-pBAD-MS2hp-XmaI-HindIII; confers SpcR |
|
|
pCW17 |
pAC-constitutive-λCIMS2CP |
Encodes residues 1-248 of CI fused to an MS2 coat protein; transcription of this protein under the control of a constitutive promoter; confers CmR |
|
|
pKB951 |
pBrα-ProQ NTD |
Encodes residues 1-248 of alpha fused to residues 1-131 of wild type
|
|
|
pKB1210 |
pCDF-pBAD-1xMS2-
|
3'UTR of
|
|
|
pASW24 |
pBrα-ProQ NTD -C24S |
|
oASW55 + oASW56 |
|
pASW25 |
pBrα-ProQ NTD -C24L |
|
oKB1552 + oKB1553 |
|
pASW26 |
pBrα-ProQ NTD -C24V |
|
oKB1552 + oKB1553 |
|
pASW27 |
pBrα-ProQ NTD -C24I |
|
oKB1552 + oKB1553 |
|
pASW28 |
pBrα-ProQ NTD -C88A |
|
oASW53 + oASW54 |
|
pASW29 |
pBrα-ProQ NTD -C88S |
|
oASW31 + oASW32 |
|
pASW30 |
pBrα-ProQ NTD -C88V |
|
oASW33 + oASW34 |
|
pASW35 |
pBrα-ProQ NTD -C24S-C88A |
|
oASW55 + oASW56 |
|
pASW39 |
pBrα-ProQ NTD -C24S-C88A-T79C |
|
oASW65 + oASW66 |
|
pASW43 |
pBrα-ProQ NTD -C24S-C88A-M50C |
|
oASW73 + oASW74 |
|
pASW44 |
pBrα-ProQ NTD -C24S-C88A-E116C |
|
oASW75 + oASW76 |
|
pASW46 |
pBrα-ProQ NTD -C24S-C88A-E90C |
|
oASW80 + oASW89 |
|
pASW47 |
pBrα-ProQ NTD -C24S-C88A-L63C |
|
oASW81 + oASW82 |
|
pASW48 |
pBrα-ProQ NTD -C24S-C88A-A47C |
|
oASW83 + oASW84 |
|
pASW49 |
pBrα-ProQ NTD -C24S-C88A-N51C |
|
oASW85 + oASW86 |
|
pHW1 |
pBrα-ProQ NTD -C24S-C88A-N3C |
|
oHW1 + oHW2 |
|
pHW2 |
pBrα-ProQ NTD -C24S-C88A-Q4C |
|
oHW3 + oHW4 |
|
pHW3 |
pBrα-ProQ NTD -C24S-C88A-D44C |
|
oHW5 + oHW6 |
|
pHW4 |
pBrα-ProQ NTD -C24S-C88A-G48C |
|
oHW7 + oHW8 |
|
pHW5 |
pBrα-ProQ NTD -C24S-C88A-S66C |
|
oHW9 + oHW10 |
|
pHW7 |
pBrα-ProQ NTD -C24S-C88A-E104C |
|
oHW13 + oHW14 |
|
pHW8 |
pBrα-ProQ NTD -C24S-C88A-V110C |
|
oHW15 + oHW16 |
|
|
|
|
|
oASW31 |
Q5 F C88S |
CGGCAACCCAaGCGGTGAGCT |
|
oASW32 |
Q5 R C88S |
TCAAGATCGACACGCGTTGC |
|
oASW33 |
Q5 F C88V |
CGGCAACCCAgtCGGTGAGCTG |
|
oASW34 |
Q5 R C88V |
TCAAGATCGACACGCGTT |
|
oASW53 |
Q5 F C88A |
CGGCAACCCAgcCGGTGAGCTG |
|
oASW54 |
Q5 R C88A |
TCAAGATCGACACGCGTTG |
|
oASW55 |
Q5 F C24S |
TTTCCCCACTcTTTCAGTGCGGAAG |
|
oASW56 |
Q5 R C24S |
ACGTTCGGCCAGAAACGC |
|
oASW65 |
Q5 F T79C |
ACCCGGCGCAtgtCGTGTCGATC |
|
oASW66 |
Q5 R T79C |
TTAACACCGTAAAGATAACG |
|
oASW73 |
Q5 F M50C |
TGCTGGGGAAtgcAACCTGAGCAAAAC |
|
oASW74 |
Q5 R M50C |
ACACGATCGACCAAATCC |
|
oASW75 |
Q5 F E116C |
ACAGCGTGCTtgtCAGCAAGCGAAAAAAC |
|
oASW76 |
Q5 R E116C |
GCCTGAACACGCGCTTTC |
|
oASW89 |
Q5 F C88A-E90C |
CCCAgccGGTtgtCTGGACGAGC |
|
oASW80 |
Q5 R C88A-E90C |
TTGCCGTCAAGATCGACAC |
|
oASW81 |
Q5 F L63C |
CGCTTTACGTtgcTACACTTCGAG |
|
oASW82 |
Q5 R L63C |
GATCGCAATTGCGTTTTG |
|
oASW83 |
Q5 F A47C |
CGATCGTGTTtgtGGGGAAATGAAC |
|
oASW84 |
Q5 R A47C |
ACCAAATCCTGAAAAATACC |
|
oASW85 |
Q5 F N51C |
TGGGGAAATGtgcCTGAGCAAAAC |
|
oASW86 |
Q5 R N51C |
GCAACACGATCGACCAAA |
|
oHW1 |
Q5 F N3C |
GGCCGCAGAAtgTCAACCTAAGTTG |
|
oHW2 |
Q5 R N3C |
GCCTCTGGTTTCTCTTCTTTC |
|
oHW3 |
Q5 F Q4C |
CGCAGAAAATtgcCCTAAGTTGAATAGCAGTAAAG |
|
oHW4 |
Q5 R Q4C |
GCCGCCTCTGGTTTCTCT |
|
oHW5 |
Q5 F D44C |
GGATTTGGTCtgTCGTGTTGCTGG |
|
oHW6 |
Q5 R D44C |
TGAAAAATACCGATTTTCAG |
|
oHW7 |
Q5 F G48C |
TCGTGTTGCTtgcGAAATGAACCTG |
|
oHW8 |
Q5 R G48C |
TCGACCAAATCCTGAAAAATAC |
|
oHW9 |
Q5 F S66C |
CTCTACACTTgcAGCTGGCGTTATC |
|
oHW10 |
Q5 R S66C |
ACGTAAAGCGGATCGCAA |
|
oHW13 |
Q5 F E104C |
CAAGCAGCTTtgcGAAGCGAAAGC |
|
oHW14 |
Q5 R E104C |
CGAGCATGCTCTACATGTTG |
|
oHW15 |
Q5 F V110C |
GAAAGCGCGTtgTCAGGCACAG |
|
oHW16 |
Q5 R V110C |
GCTTCTTCAAGCTGCTTG |
|
oKB1552 |
Q5 F C24L/V/I |
TTTTCCCCACntaTTCAGTGCGGAAGGTGAAGCG (degenerate codon encodes each of 3 amino acids; individual colonies were sequenced to clone each individual mutant) |
|
oKB1553 |
Q5 R C24L/V/I |
CGTTCGGCCAGAAACGCG |
|
|
|
|
|
|
NEB 5α F’Iq |
|
TetR |
New England Biolabs |
|
KB473 |
FW102
|
KanR, StrR |
|
- —National Institute of General Medical Sciences (United States)https://ror.org/04q48ey07
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsRNA modifications and cancer · RNA and protein synthesis mechanisms · RNA Research and Splicing
Description
RNA-binding proteins (RBPs) play essential roles in gene regulation in all organisms, including bacteria. Hfq is the most well characterized bacterial RNA-chaperone protein that stabilizes small RNAs (sRNAs) and facilitates sRNA base pairing with 5’ untranslated regions (UTRs) of mRNAs. One class of bacterial RBPs that has garnered recent attention is FinO-domain proteins [ (Liao and Smirnov 2023) ](https://www.zotero.org/google-docs/?vp7EmL) ; the *E. coli * ProQ protein is a global RBP with many sRNA and mRNA targets [ (Olejniczak and Storz 2017; Holmqvist et al. 2020) ](https://www.zotero.org/google-docs/?8tbTTP) (Fig 1a,b). The core RNA-binding site of ProQ is found in a conserved concave pocket in the FinO-like N-terminal domain (NTD), which recognizes the 3’ base and poly(U) tail of intrinsic terminators [ (Kim et al. 2022; Stein et al. 2023; Pandey et al. 2020) ](https://www.zotero.org/google-docs/?L7Hnll) . Still, several important questions remain unanswered about how ProQ interacts with RNA, such as: what role the C-terminal domain (CTD) and/or unstructured linker of *E. coli * ProQ play in RNA interactions; the degree of conformational flexibility and heterogeneity of ProQ-RNA interactions; whether RNA upstream of the intrinsic terminator participates in transient protein interactions; which surfaces ProQ uses to interact with RNA ligands that do not possess a 3’ intrinsic terminator, such as fragments of 5’ UTRs or open-reading frames (ORFs) [ (Holmqvist et al. 2018; Melamed et al. 2020) ](https://www.zotero.org/google-docs/?fSY4f7) ; and whether ProQ can interact with two RNAs simultaneously or otherwise promote restructuring of RNAs. These mechanistic questions are important to fully understanding the role ProQ plays in regulating RNA stability and gene expression.
Several biochemical and biophysical techniques make use of site-directed bioconjugation of macromolecules for functionalization with small-molecule ligands [ (Hermanson 2013) ](https://www.zotero.org/google-docs/?hJxA2p) . For instance, site-specific hydroxyl radical probing with an FeBABE reagent can reveal details of RNA-protein interaction mechanisms [ (Duval et al. 2017) ](https://www.zotero.org/google-docs/?hhpl4W) . This approach makes use of the uniquely nucleophilic thiol group of cysteine side-chains [ (Gunnoo and Madder 2016) ](https://www.zotero.org/google-docs/?JYDffr) : single-cysteine variants of a protein of interest are achieved by the removal of surface-exposed native cysteines and the installation of new cysteine residues that can be site-specifically labeled with a thiol-reactive FeBABE molecule [ (Duval et al. 2017) ](https://www.zotero.org/google-docs/?cI84xI) . An important step in this process is identifying locations of cysteine variants that do not interfere with the native protein’s function. In order to identify single-cysteine variants of *E. coli * ProQ that retain the ability to bind to RNA targets, we utilized a bacterial three-hybrid assay (B3H; [ (Berry and Hochschild 2018; Stockert et al. 2022) ](https://www.zotero.org/google-docs/?mL0mUs) to test the RNA-binding activity of a panel of cysteine-free and single-cysteine variants of this protein. Briefly, this genetic assay makes use of a *lacZ * reporter gene where transcription is increased by interaction between a ‘bait’ RNA moiety and a ‘prey’ protein of interest (Fig 1c; see methods).
*E. coli * ProQ naturally possesses two cysteine residues—both in the N-terminal FinO domain (Fig 1a,b). We began by mutating each of these cysteine codons to either hydrophilic or hydrophobic amino acids ( e.g. serine, leucine, valine or alanine) and testing their interactions in a B3H assay with the *malM * 3’ UTR (Fig 1d,e) *. * This RNA has been previously identified as a ProQ interactor in CLIP-seq and RIL-seq experiments [ (Holmqvist et al. 2018; Melamed et al. 2020) ](https://www.zotero.org/google-docs/?Lud0v4) and its interaction with ProQ has been previously characterized in the B3H assay [ (Stein et al. 2020, 2023) ](https://www.zotero.org/google-docs/?BTMxB0) . In the B3H analysis of ProQ binding to malM-3’ , Cys24 was most robustly replaced by a serine residue (C24S), while Cys88 was best replaced by an alanine (C88A). A cysteine-free ProQ containing both of these substitutions together (C24S-C88A) retained ~70% of WT ProQ’s interaction with *malM-3’ * (Fig 1f) *. * Using this cysteine-free C24S-C88A variant, we selected 14 additional residues that were predicted to be surface-exposed and not highly conserved to mutate to cysteine. These single-cysteine variants were tested for malM-3’ interaction (Fig 1g). While some cysteine mutants were strongly disruptive to RNA interaction ( e.g. G48C, D44C, S66C), 11 mutants retained at least 70% of the cysteine-free variant’s interaction and were deemed suitable for future purification and site-directed conjugation, along with the C88A and C24S single-cysteine proteins. Importantly, this collection of 13 single-cysteine mutants encircles both the concave and convex faces of ProQ (Fig 1h).
Because the interaction of 3’ UTRs with ProQ does not depend strongly on the CTD or unstructured linker [ (Pandey et al. 2020; Stein et al. 2020) ](https://www.zotero.org/google-docs/?KQZomr) , we were unable to test single-cysteine mutants in these regions. However, it should be possible to introduce cysteine residues into positions of limited conservation that are predicted to be surface-exposed. One caveat to keep in mind is that ProQ variants may be less stable *in vitro * than they are when expressed inside of the cell as fusion proteins with α-NTD. It is also possible that a cysteine would be tolerated at a given position but that a larger moiety such as FeBABE or a fluorophore would sterically disrupt interaction. The RNA binding of site-specifically labeled proteins should therefore be confirmed under the conditions used for downstream biochemical or biophysical experiments.
Methods
**Bacterial Strains and Plasmids. ** Plasmids, oligonucleotides and bacterial strains used in this study are provided in the Reagents section below. Site-directed mutants were constructed using Q5 site-directed mutagenesis (New England Biolabs) as previously described [ (Stein et al. 2023) ](https://www.zotero.org/google-docs/?DrqgVW) with the specified mutagenic end-to-end primers, designed with NEBaseChanger.
**β-galactosidase assays. ** RNA binding of ProQ mutants were assessed using a previously established bacterial three-hybrid (B3H) assay (Fig 1c) [ (Stockert et al. 2022; Berry and Hochschild 2018) ](https://www.zotero.org/google-docs/?9BOLBi) . Bait RNA is anchored to the B3H system by a DNA-RNA ‘adapter’ protein that consists of a fusion between the CI protein from bacteriophage 𝜆 and the bacteriophage MS2 coat protein. Bait RNA is fused to an MS2 RNA hairpin (MS2 ^hp^ ). The prey protein is fused to the NTD of the RNA polymerase (RNAP) alpha subunit (α). Interaction between the bait RNA and prey protein is measured by the fold-stimulation of lacZ activity when both bait and prey are present in reporter cells over basal levels, defined by the highest value of negative controls when either bait or prey protein is missing.
For B3H assays, Δ hfq p lac -O L 2–62 reporter strain cells (KB473) were transformed with three compatible plasmids: first by pAdapter (pCW17), and subsequently with a combination pPrey and pBait plasmids. pPrey plasmids encoded either the WT α-ProQ ^NTD ^ fusion protein (resi=2-131; pKB951 or a site-specific mutant derivative) or α alone as a negative control. pBait plasmids encoded a hybrid RNA with the 3’ UTR of *malM * following the MS2 ^hp^ (pKB1210) or an RNA that contained only the MS2 ^hp^ moiety (pCH1) as a negative control. Transformations were conducted in 96-well plates as previously described [ (Stockert et al. 2022) ](https://www.zotero.org/google-docs/?SPR4Wg) . Overnight cultures were directly inoculated with transformants and grown in 1 mL of LB supplemented with carbenicillin (100 μg/mL), chloramphenicol (25 μg/mL), kanamycin (50 μg/mL), spectinomycin (100 μg/mL) and 0.2% arabinose. Plates were sealed with breathable film and grown ON at 37℃, shaking at 900 rpm. In the morning, day cultures (200 μL) were inoculated with 10 μL of ON culture, covered with a plastic lid and grown at 37℃, shaking at 900 rpm until OD 600 values reached mid-log (0.3-0.6). Cells were lysed with the addition of 10 μL of PopCulture Reagent (Novagen) and of 4 units of rLysozyme (Novagen). β-gal activity was measured as previously described [ (Stockert et al. 2022) ](https://www.zotero.org/google-docs/?5F7TLh) .
The β-gal activity produced by reporter cells transformed with each combination of bait RNA and prey protein was divided by the highest value of the two corresponding negative controls to yield fold-stimulation over basal lacZ levels. These fold-stimulation values were then normalized using the equation: (fold-stimulation ^mutant^ -1)/(fold-stimulation ^comparison^ -1), where the point of comparison was either the fold-stimulation produced by ProQ ^WT ^ or ProQ ^C24S-C88A^ . This normalization sets the B3H value for the highest value to 1.0 and the lack of a detectable interaction (1-fold-stimulation of lacZ) equal to 0.
**Protein Structures. ** The positions of native and introduced cysteine residues were mapped onto the AlphaFold predicted structure of *E. coli * ProQ [ (Jumper et al. 2021; Varadi et al. 2022) ](https://www.zotero.org/google-docs/?TnrVwH) , which has been found to be consistent with genetic mutagenesis data [ (Stein et al. 2023) ](https://www.zotero.org/google-docs/?Zxlp5N) . Structures were rendered in the PyMOL Molecular Graphics System (Schrödinger, LLC).
Reagents
Plasmids used in this study:
**: **
Oligonucleotides used in this study:
**: **
Bacterial strains used in this study:
**: **
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Berry KE Hochschild A 2018125 A bacterial three-hybrid assay detects Escherichia coli Hfq-s RNA interactions in vivo.Nucleic Acids Res 4620305-1048 e 12e 1210.1093/nar/gkx 108629140461 PMC 5778611 · doi ↗ · pubmed ↗
- 2Dove SL Joung JK Hochschild A 1997410 Activation of prokaryotic transcription through arbitrary protein-protein contacts.Nature 38666250028-083662763010.1038/386627 a 09121589 · doi ↗ · pubmed ↗
- 3Duval M Marenna A Chevalier C Marzi S 20161224 Site-Directed Chemical Probing to map transient RNA/protein interactions.Methods 1171046-2023485810.1016/j.ymeth.2016.12.01128027957 · doi ↗ · pubmed ↗
- 4Gunnoo SB Madder A 201639 Chemical Protein Modification through Cysteine.Chembiochem 1771439-422752955310.1002/cbic.20150066726789551 · doi ↗ · pubmed ↗
- 5Hermanson GT. 2013. Bioconjugate Techniques . Elsevier Science & Technology https://linkinghub.elsevier.com/retrieve/pii/B 9780123822390000017
- 6Holmqvist E Berggren S Rizvanovic A 2020619 RNA-binding activity and regulatory functions of the emerging s RNA-binding protein Pro Q.Biochim Biophys Acta Gene Regul Mech 186391874-939919459619459610.1016/j.bbagrm.2020.19459632565402 · doi ↗ · pubmed ↗
- 7Holmqvist E Li L Bischler T Barquist L Vogel J 2018524 Global Maps of Pro Q Binding In Vivo Reveal Target Recognition via RNA Structure and Stability Control at m RNA 3' Ends.Mol Cell 7051097-2765971982.e 610.1016/j.molcel.2018.04.01729804828 · doi ↗ · pubmed ↗
- 8Jumper J Evans R Pritzel A Green T Figurnov M Ronneberger O Tunyasuvunakool K Bates R Žídek A Potapenko A Bridgland A Meyer C Kohl SAA Ballard AJ Cowie A Romera-Paredes B Nikolov S Jain R Adler J Back T Petersen S Reiman D Clancy E Zielinski M Steinegger M Pacholska M Berghammer T Bodenstein S Silver D Vinyals O Senior AW Kavukcuoglu K Kohli P Hassabis D 2021715 Highly accurate protein structure prediction with Alpha Fold.Nature 59678730028-083658358910.1038/s 41586-021-03819-234265844 PMC 8371605 · doi ↗ · pubmed ↗