Single‐cell transcriptomic landscape reveals distinct tumourigenesis and immune microenvironments in secondary radiation‐exposed rectal cancer
Xu Guan, Xiaoman Bi, Ran Wei, Zhixun Zhao, Zhao Lu, Zheng Jiang, Xishan Wang, Deng Wu

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
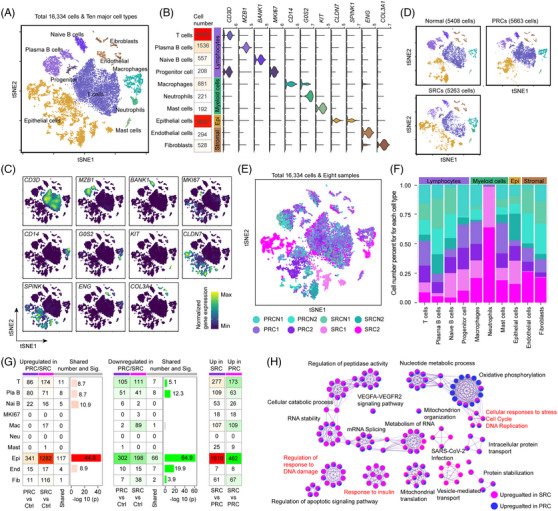 Figure 1
Figure 1 Figure 2
Figure 2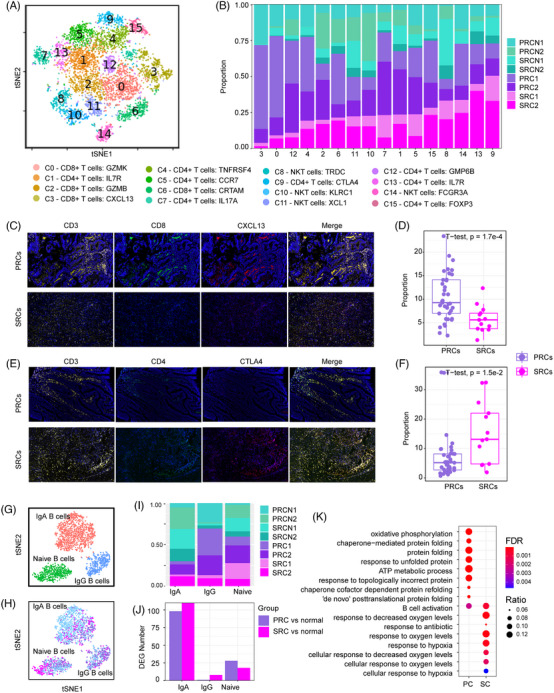 Figure 3
Figure 3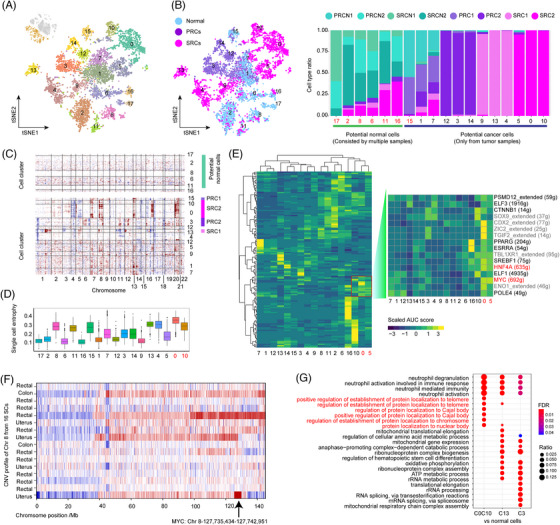 Figure 4
Figure 4- —The Sanming Project of Medicine in Shenzhen
- —National Natural Science Foundation of China 10.13039/501100001809
- —Hainan Provincial Natural Science Foundation of China
- —Youth Science and Technology Talent Innovation Program of Hainan Association for Science and Technology
- —Postdoctoral Science Foundation of Hainan Province
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCancer Immunotherapy and Biomarkers · Effects of Radiation Exposure · Cancer Cells and Metastasis
Dear Editor,
Radiation therapy is a common treatment option for pelvic cancer patients1 and can induce secondary radiation‐exposed rectal cancers (SRCs).2 However, compared with those of primary rectal cancers (PRCs), the molecular characteristics and the cause of SRCs are not fully understood.3 By sequencing the single‐cell transcriptome to reveal the cell type‐specific profile, this study revealed an augmented copy number variation (CNV) profile and a distinct immune microenvironment in SRCs.
By comprehensively evaluating the tumour types that occurred with the therapeutic ionising radiation and the short time, compared to the evolution of PRCs, a total of 18 SRCs were diagnosed (Table S1). Two classical SRCs and two PRCs as well as their normal tissues were sequenced at single‐cell resolution (Figure S1A, Table S1), resulting in 16 334 cells (Figure S1B) from 10 broad cell types, including epithelial cells, endothelial cells, fibroblasts, T cells, plasma B cells, naïve B cells, mast cells, macrophages, neutrophils and MKI67 progenitor cells (Figure 1A), according to their validated markers4, 5 (Figures 1B,C and S1B,C). t‐Distributed stochastic neighbour embedding (t‐SNE) analysis suggested that the major difference between normal cells, PRCs and SRCs was derived from epithelial cells (Figure 1D,E) and neutrophils (Figure 1D–F). A total of 2297 unique differentially expressed genes (DEGs) was identified (Figure 1G, Table S2) between tumour and normal tissues. A direct comparison of epithelial cells between PRCs and SRCs revealed that tumour‐activated pathways, such as DNA repair, apoptosis inhibition and mitosis, were significantly enhanced in SRCs, while oxidative phosphorylation was upregulated in PRCs (Figures 1H and S1D).
A total of 18 subclusters was obtained from epithelial cells (Figure 2A), six subclusters were initially treated as candidate normal cells (Figures 2B and S2A), and another 12 subclusters were defined as potential tumour cells because they originated from one tumour sample (Figures 2B and S2A). DNA CNV analysis confirmed that Cluster 0 and 10 displayed large‐scale genomic rearrangements, including gains of chr 8q, chr 16q and chr 20p (Figures S2B and 2C). Cluster 13, which originated from another SRC patient, shared a gain of chr 8q with Cluster 0 and 10. The PRC‐related clusters exhibiting a substantial number of CNV were Clusters 3 and 15. Overall, we defined Cluster 0, 10 and 13 as the SRC tumour cells, which displayed the highest CNV score (Figure S2C) and the highest differentiation capacity (Figures 2D and S2C) regardless of the dimensionality reduction method (Figure S2D,E). The specific highly expressed genes from the SRC tumour cells were validated in additional SRC samples (Figure S2G–I) further confirming the common features of the SRC malignant cells.
The amplification of chr 8q led to increased gene expression of the tumour driver gene MYC (Figure 2E), an activated transcription factor across tumour cells (Figure 2E), which has been observed in secondary angiosarcoma6 and radiogenic adenocarcinoma.7 The CNV profile of the 18 additional SRCs determined by whole‐genome sequencing further validated the gain of chr 8q among the SRCs (Figure 2F). These additional CNVs might contribute to telomere maintenance among these tumour cells (Figure 2G).
We next reclustered the immune cells and identified 16 T‐cell subclusters (Figure 3A) from CD4^+^, CD8^+^ and natural killer T cells (Figure S3A–C). CD4^+^ clusters contained T helper 1 cells (IL7R/C1, C13), T helper 17 cells (IL17A/C7), activated T cells (TNFRSF4/C4), naïve/center memory T cells (CCR7/C5), two types of regulatory T cells (CTLA4/C9, FOXP3 /C15) and another unsigned subcluster marked by GPM6B. Among the eight subclusters of CD8^+^ T cells, two were CD8^+^ effector T cells (GZMK/C0, GZMB/C2), one highly expressed CXCL13 (C3), and the other was activated CD8^+^ T cells (CRTAM/C6). The remaining three clusters belong to natural killer cells (KLRC1/C10, XCL1/C11 and FCGR3A/C14).
We found that the number of CXCL13^+^ CD8^+^ T cells (C3) increased in the PRCs (Figure 3B) and CTLA4^+^ CD4^+^ T cells (C9) accumulated in the SRCs (Figure 3B). Analysis of tissue sections stained for CXCL13 (Figure 3C) and CTLA4 (Figure 3E) in an additional 39 PRCs and 13 SRCs further validated a significantly lower percentage of CXCL13^+^ CD8^+^ T cells in the PRCs (Figure 3D) and a greater percentage of CTLA4^+^ CD4^+^ T cells in SRCs (Figure 3F) than in the normal tissues.
In addition to the above divergent T‐cell subtype shift between PRCs and SRCs, cell proliferation‐associated genes FOS and JUN are differentially regulated between PRC and SRCs (Figure S3D,E), and the expression of cell migration‐associated genes, including TMSB10 and TMSB4X, decreased in SRCs but did not change in PRCs (Figure S3D,E). The expression of heat shock response‐associated genes8 increased in both PRCs and SRCs (Figure S3D,E).
Naïve B cells, IgA B cells and IgG B cells were recognised among the B cells (Figure 3G,H). IgA B cells were the most significantly affected in both the PRCs and SRCs (Figure 3I) and had the most DEGs in both PRCs and SRCs (Figure 3J). Oxidative phosphorylation and the ATP metabolic process were enriched in PRCs, and hypoxia response was overrepresented in SRCs (Figure 3K).
Among the myeloid cell subtypes (Figures 4A and S4A), neutrophils were observed in SRCs (Figure 4B), and all subtypes of macrophages increased in both PRCs and SRCs (Figure S4B). A greater number of DEGs were found in M2‐like (tumour‐like) macrophages between SRCs and normal tissue (Figure 4C).
Next, we investigated cell–cell communication. Both outgoing and incoming signalling revealed the strongest signal from SRCs (Figure 4D, Table S3). The LAMB3 and LAMC2‐mediated (Figure 4E) laminin crosstalk was observed only in SRCs (Figure 4F). Tight communication between neutrophils and other cells was also observed, through the high expression of ligands including S100A9, S100A8 and CXCL8 and receptors, including CXCR2 and CXCR4.
In conclusion, we found that oncogene amplification during radiotherapy might promote the progression of SRCs, and the activation of Treg cells highly expressed CTLA4 promote tumour cell survival. The emergence of neutrophils and the activation of macrophages provided an additional immune microenvironment for SRCs. Our findings provide a comprehensive understanding of SRC tumourigenesis, and the activation of CTLA4‐marked Tregs also provides theoretical support for the combination of radiotherapy and anti‐CTLA‐4 therapy.9
AUTHOR CONTRIBUTIONS
Deng Wu, Xishan Wang and Xu Guan conceptualised the study. Deng Wu and Xiaoman Bi performed bioinformatics analysis. Xu Guan and Ran Wei performed histology‐based experiments. Deng Wu, Xu Guan and Xiaoman Bi wrote and discussed the manuscript. Zhixun Zhao, Zhao Lu and Zheng Jiang collected the samples. Deng Wu and Xu Guan supervised the project. All authors have read and approved the final manuscript.
FUNDING INFORMATION
The work was supported, in part, by grants from the following: The Sanming Project of Medicine in Shenzhen (Ref: SZSM201911012); the National Natural Science Foundation of China (Ref: 82072750); Hainan Provincial Natural Science Foundation of China (Ref: 822QN462 and 823RC581); Youth Science and Technology Talent Innovation Program of Hainan Association for Science and Technology (Ref: QCQTXM202212) and Postdoctoral Science Foundation of Hainan Province.
CONFLICT OF INTEREST STATEMENT
All authors report there are no potential conflicts of interest.
ETHICS STATEMENT
This study had been approved by the Ethics Committee of Cancer Hospital, Chinese Academy of Medical Sciences (Approval ID:23/139‐3881)
Supporting information
Supporting Information
Supporting Information
Supporting Information
Supporting Information
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Yang J , Cai H , Xiao ZX , Wang H , Yang P . Effect of radiotherapy on the survival of cervical cancer patients: an analysis based on SEER database. Medicine (Baltimore). 2019;98:e 16421.31348242 10.1097/MD.0000000000016421 PMC 6708958 · doi ↗ · pubmed ↗
- 2Travis LB , Ng AK , Allan JM , et al. Second malignant neoplasms and cardiovascular disease following radiotherapy. J Natl Cancer Inst. 2012;104:357‐370.22312134 10.1093/jnci/djr 533PMC 3295744 · doi ↗ · pubmed ↗
- 3Dracham CB , Shankar A , Madan R . Radiation induced secondary malignancies: a review article. Radiat Oncol J. 2018;36:85‐94.29983028 10.3857/roj.2018.00290 PMC 6074073 · doi ↗ · pubmed ↗
- 4Khaliq AM , Erdogan C , Kurt Z , et al. Refining colorectal cancer classification and clinical stratification through a single‐cell atlas. Genome Biol. 2022;23:113.35538548 10.1186/s 13059-022-02677-z PMC 9092724 · doi ↗ · pubmed ↗
- 5Lee HO , Hong Y , Etlioglu HE , et al. Lineage‐dependent gene expression programs influence the immune landscape of colorectal cancer. Nat Genet. 2020;52:594‐603.32451460 10.1038/s 41588-020-0636-z · doi ↗ · pubmed ↗
- 6Guo T , Zhang L , Chang NE , Singer S , Maki RG , Antonescu CR . Consistent MYC and FLT 4 gene amplification in radiation‐induced angiosarcoma but not in other radiation‐associated atypical vascular lesions. Genes Chromosomes Cancer. 2011;50:25‐33.20949568 10.1002/gcc.20827 PMC 3150534 · doi ↗ · pubmed ↗
- 7Wade MA , May FE , Onel K , Allan JM . Does radiation‐induced c‐MYC amplification initiate breast oncogenesis? Mol Cell Oncol. 2016;3:e 1010950.27308527 10.1080/23723556.2015.1010950 PMC 4845163 · doi ↗ · pubmed ↗
- 8Chu Y , Dai E , Li Y , et al. Pan‐cancer T cell atlas links a cellular stress response state to immunotherapy resistance. Nat Med. 2023;29:1550‐1562.37248301 10.1038/s 41591-023-02371-y PMC 11421770 · doi ↗ · pubmed ↗