Biochemical Fractionation of Human α-Synuclein in a Drosophila Model of Synucleinopathies
Khondamir Imomnazarov, Joshua Lopez-Scarim, Ila Bagheri, Valerie Joers, Malú Gámez Tansey, Alfonso Martín-Peña

TL;DR
This study develops a method to analyze human α-synuclein solubility in fruit fly brains, helping understand how it forms harmful clumps in diseases like Parkinson's.
Contribution
A reproducible protocol was developed to fractionate human α-synuclein in Drosophila brains based on solubility.
Findings
A 5% SDS buffer and three-step protocol effectively separates soluble and insoluble α-synuclein fractions.
Sonication increases solubility of α-synuclein by breaking insoluble complexes in fly brains.
Abstract
Synucleinopathies are a group of central nervous system pathologies that are characterized by the intracellular accumulation of misfolded and aggregated α-synuclein in proteinaceous depositions known as Lewy Bodies (LBs). The transition of α-synuclein from its physiological to pathological form has been associated with several post-translational modifications such as phosphorylation and an increasing degree of insolubility, which also correlate with disease progression in post-mortem specimens from human patients. Neuronal expression of α-synuclein in model organisms, including Drosophila melanogaster, has been a typical approach employed to study its physiological effects. Biochemical analysis of α-synuclein solubility via high-speed ultracentrifugation with buffers of increasing detergent strength offers a potent method for identification of α-synuclein biochemical properties and the…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
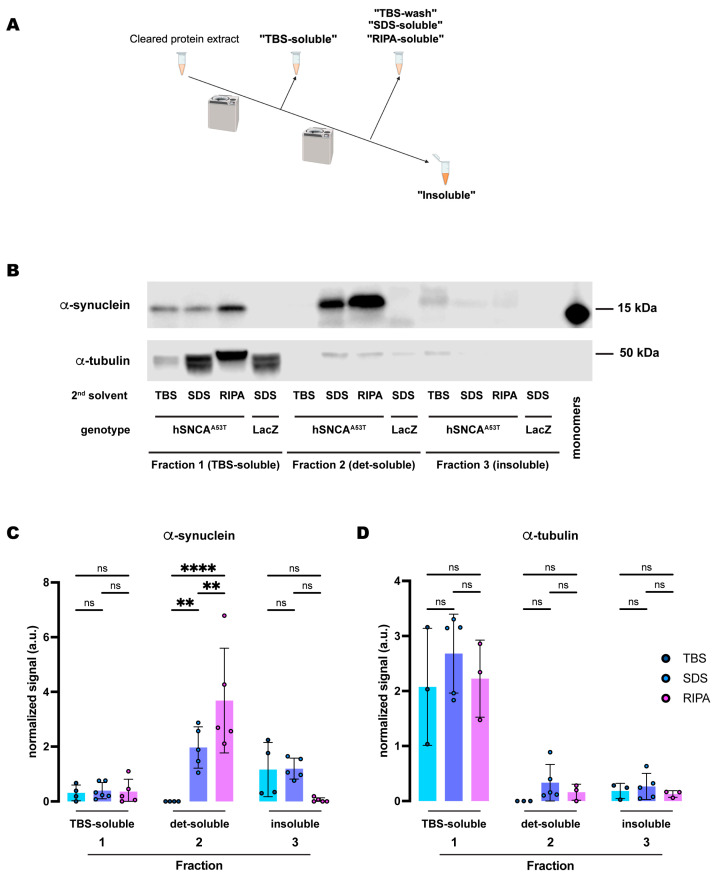 Figure 1
Figure 1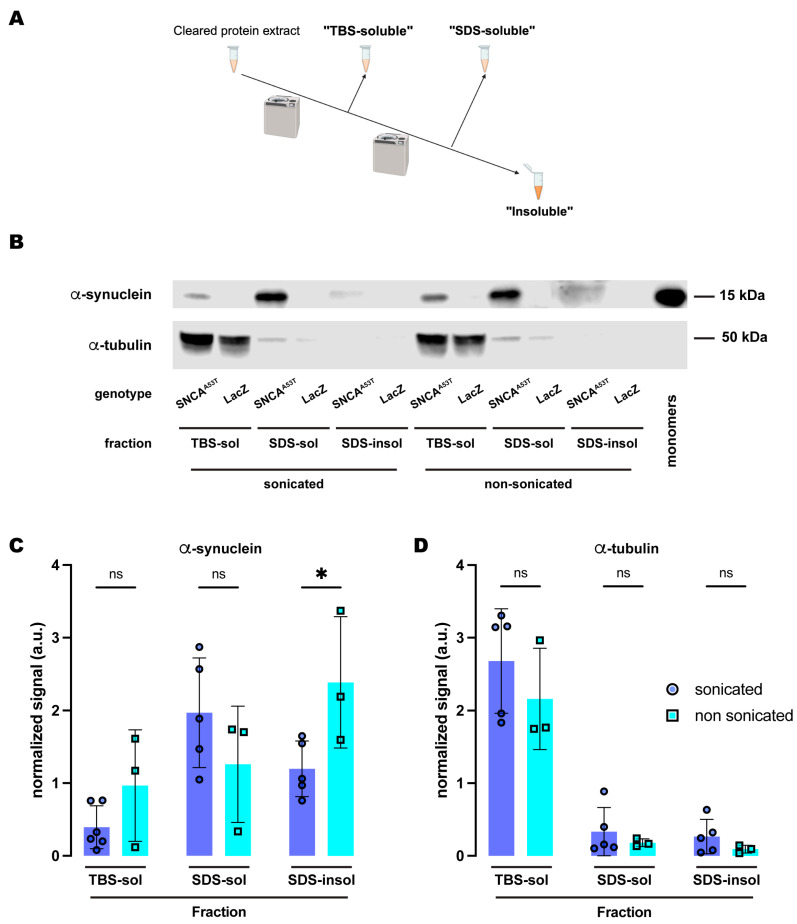 Figure 2
Figure 2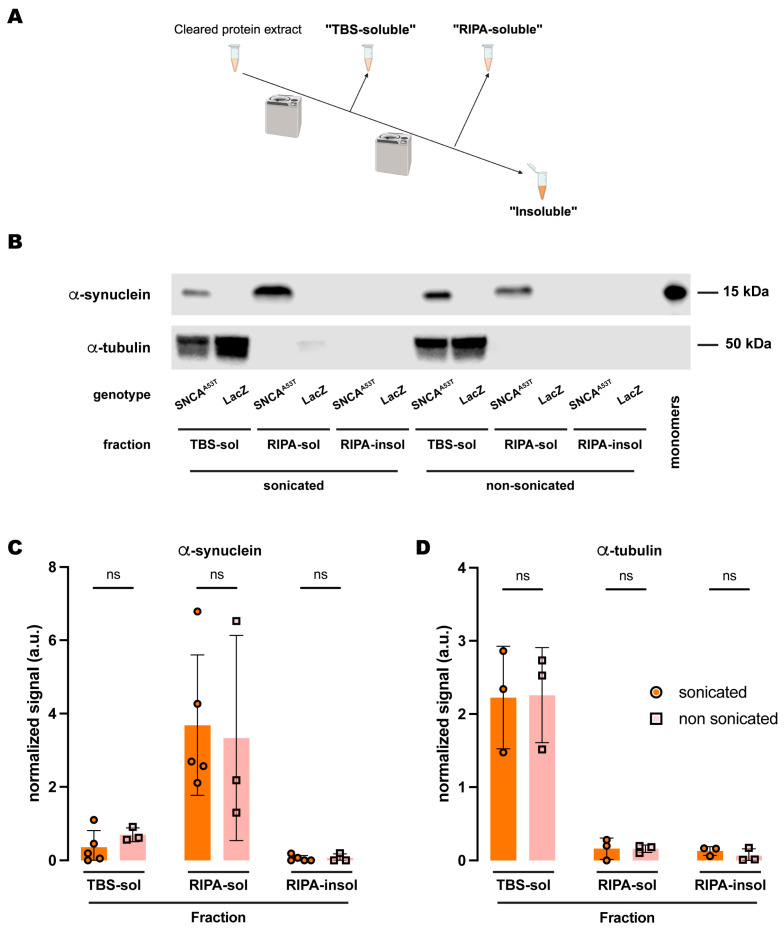 Figure 3
Figure 3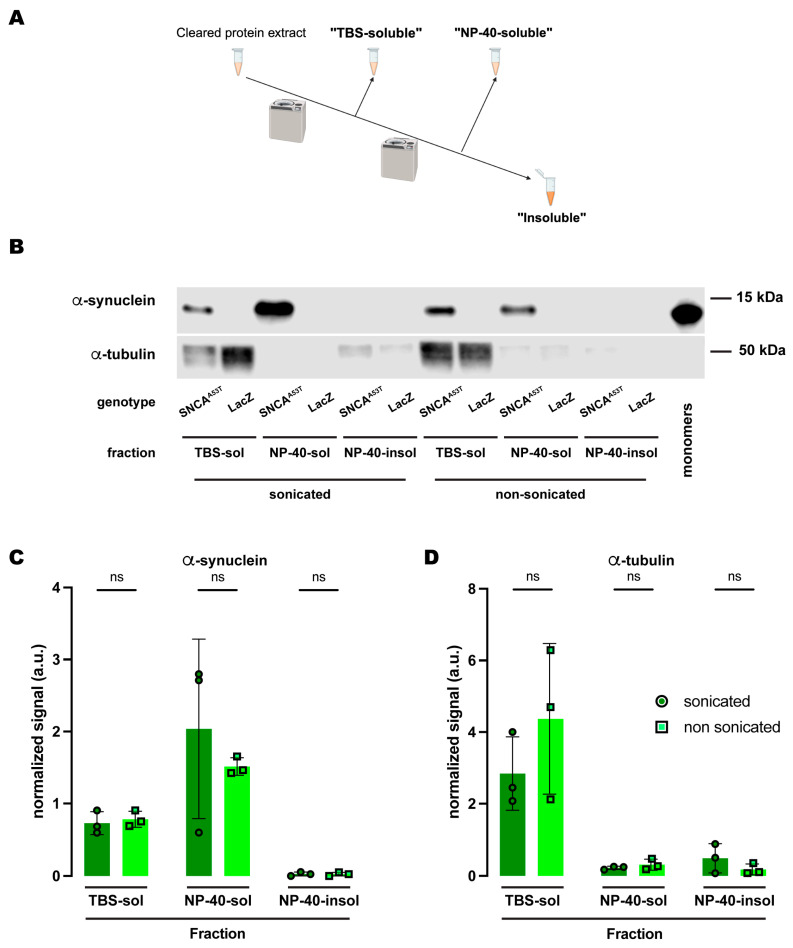 Figure 4
Figure 4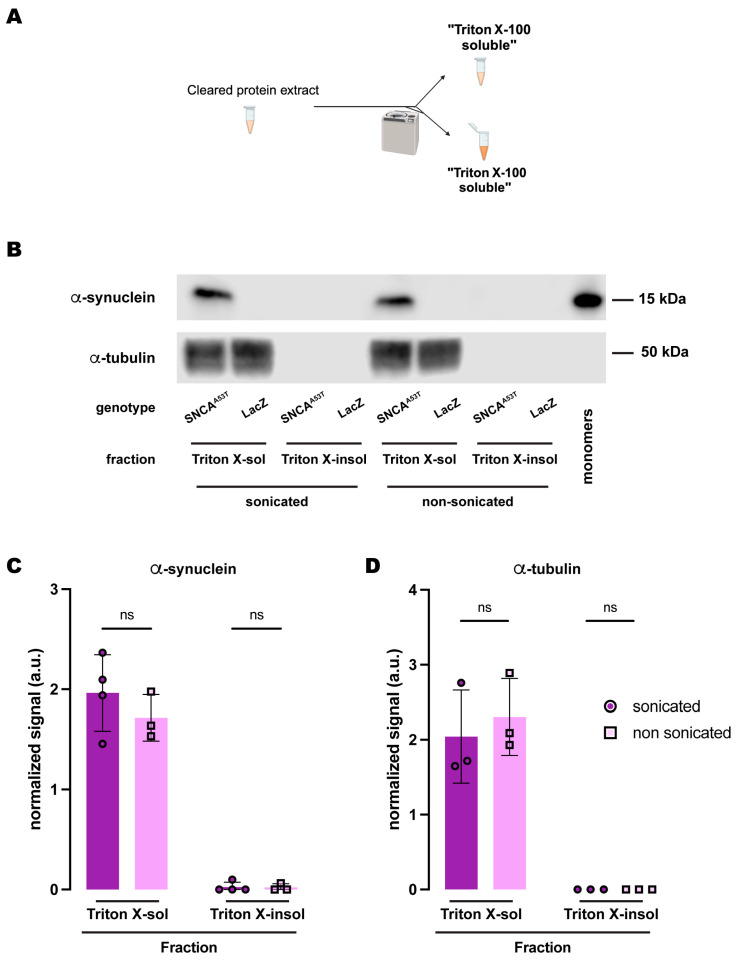 Figure 5
Figure 5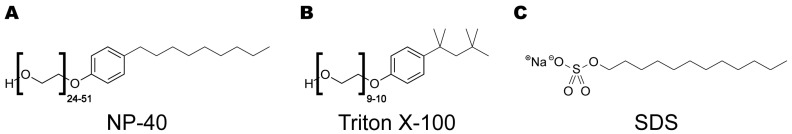 Figure 6
Figure 6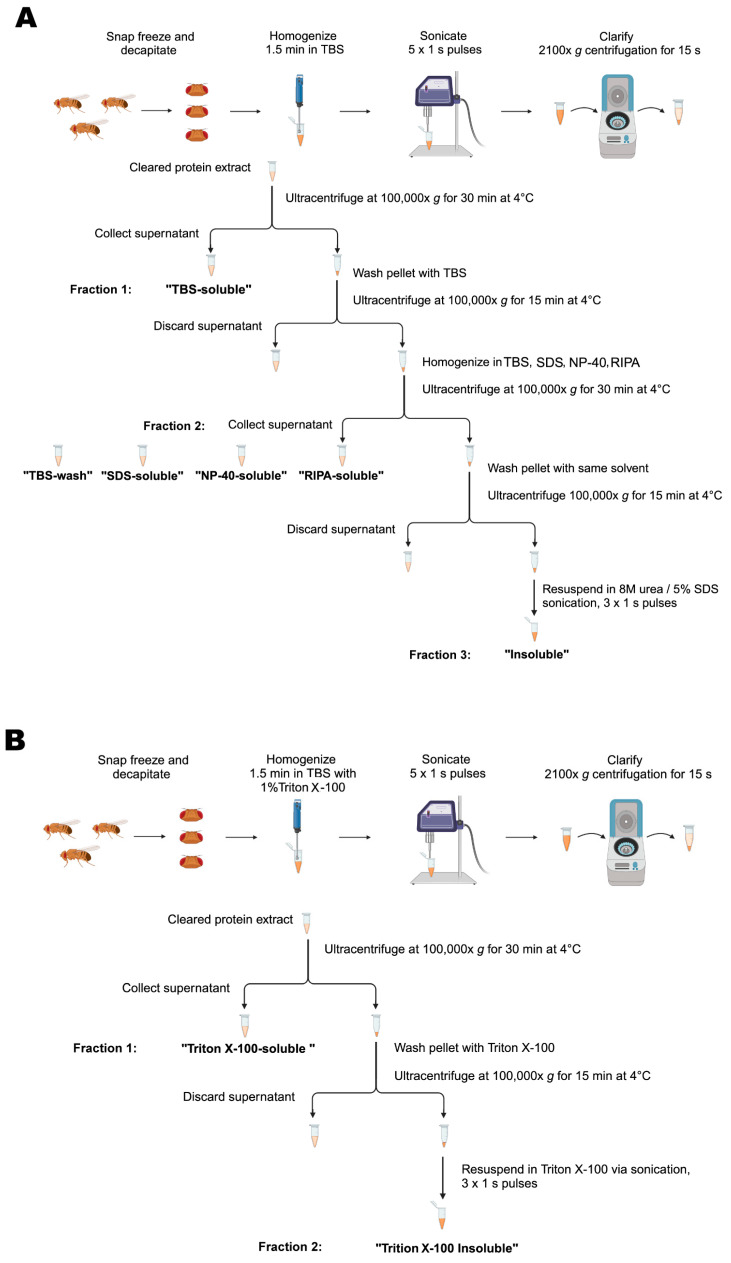 Figure 7
Figure 7- —Alzheimer’s Association
- —Center for Translational Research in Neurodegenerative Disease
- —Parkinson’s Foundation
- —NIH NINDS
- —Aligning Science Across Parkinson’s
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsParkinson's Disease Mechanisms and Treatments · Alzheimer's disease research and treatments · Genetic Neurodegenerative Diseases
1. Introduction
Misfolding and aggregation of the product encoded in the human α-synuclein gene (hSNCA) characterizes a group of nervous system disorders known as synucleinopathies [1,2]. These disorders include Parkinson’s disease (PD), the second most common neurodegenerative disease, PD dementia (PDD), multiple system atrophy (MSA), Lewy Body dementia (LBD) and dementia with Lewy Body (DLB), among others [1,2], many of which are classified under the umbrella term of parkinsonism due to their effects on motor control (slowed movement, rigidity, and/or tremor). A common pathological hallmark of synucleopathies is the deposition of α-synuclein in intracellular neuronal inclusions termed Lewy Bodies (LB).
Pathological α-synuclein is the major protein component of LBs, where it is mainly found in its insoluble fibrillar form. Conversely, in neurons, physiological α-synuclein is located in the cytoplasm and/or is associated with membranous structures in synaptic terminals in a soluble monomeric form [3]. Although monomeric α-synuclein can aggregate spontaneously to form amyloid fibrils, which are the core components of LBs, α-synuclein presents a stable conformation upon binding to lipid membranes [4]. The transition of α-synuclein from its benign soluble form into the amyloidogenic fibrillar conformation that constitutes the principal scaffold component of LBs is thought to be caused by extensive post-translational modifications [5,6,7,8,9,10], rather than a mere increase in its concentration [4,11]. Although, protein abundance positively correlates with a higher propensity for aggregation [12,13,14]. Furthermore, the presence of LB and, consequently, histopathologic deposition of α-synuclein occurs in 80–90% of PD cases.
In general, the increasing insolubility of α-synuclein and a decrease in soluble α-synuclein in the frontal cortex of postmortem PD brains were reported to positively correlate with both disease duration and disease stage [15]. Postmortem samples of the basal ganglia and the limbic cortex from patients with idiopathic PD also show a drastic increase in insoluble α-synuclein content compared to age-matched healthy controls [16].
Given the major role that α-synuclein plays in the pathophysiology of these neurodegenerative diseases, the ability of the fruit fly, Drosophila melanogaster, to express human α-synuclein represents an invaluable system for investigating its effects on brain physiology, behavior and disease progression, in PD in particular. The genetic amenability and tractability of the fly have provided considerable insights into PD pathogenesis [17,18,19,20,21,22,23,24,25,26,27,28,29,30,31,32,33,34], demonstrating, for instance, the selective vulnerability of dopaminergic neurons to α-synuclein toxicity [22], the protective role of glucocerebrosidase against α-synuclein aggregation [19,27], the importance of the vacuolar protein sorting 35 (VPS35) for lysosomal α-synuclein degradation [26] or the protective activity of the protein deglycase DJ-1 against oxidative stress [25].
The original work on the expression of hSNCA in the fly brain provided histological evidence of α-synuclein aggregation, revealing the punctate immunostaining characteristic of proteinaceous aggregates, and electron micrographs illustrating α-synuclein inclusions with radiating protein filaments in neurons [22]. However, a robust biochemical characterization of these α-synuclein inclusions in Drosophila models has been lacking; therefore, the extent to which these α-synuclein aggregates resemble those found in LBs from human brain specimens remains unclear. Given that the progression in the pathophysiology of these diseases strongly depends on the transition from monomeric soluble α-synuclein to increasingly insoluble species [15], it becomes critical to develop a robust protocol to identify the specific α-synuclein species and their solubility status at each stage of the model organism’s lifespan. Developing the tools and techniques for this identification is instrumental to unravelling the molecular mechanisms that underlie the transition from physiological soluble monomers to pathological insoluble aggregates or detecting their therapeutic effects on α-synuclein assemblies for future physiological, behavioral or treatment studies using Drosophila models of synucleopathies.
Here we implemented a multi-step fractionation protocol in which the second fraction employs a buffer containing 5% SDS that solubilizes membrane-associated α-synuclein but not insoluble α-synuclein. The combination of this buffer with a three-step serial fractionation procedure exhibits a resolution that we could use to detect changes in α-synuclein segregation upon sonication of the samples prior to the fractionation protocol, also demonstrating that the aggregates formed in DA neurons of Drosophila can be disaggregated upon this mechanical application.
2. Results
2.1. Differential Solubility of α-Synuclein in Selective Detergents
Disease progression largely correlates with the species-specific (from monomers to oligomers and fibrils) accumulation of α-synuclein and its deposition into insoluble neuronal inclusions (LBs) [15]. Chemical fractionation of human specimens or tissues from vertebrate models has provided benchmark methods for the quantification of α-synuclein solubility and the identification of pathology stage. To translate this methodology to Drosophila models of synucleinopathies, we first tested different buffers containing selective detergents for their ability to fractionate human α-synuclein (hSNCA) expressed in dopaminergic (DA) neurons of the fly brain. Flies were aged up to 20 days post-eclosion at 25C to allow time for α-synuclein aggregation; by this time point, the flies had developed significant behavioral and cellular phenotypes [22]. At this point, we snap-froze the flies in liquid nitrogen and collected the heads of the controls (TH-Gal4/UAS-LacZ) and α-synuclein expressing flies (TH-Gal4/UAS-hSNCA^A53T^). We then homogenized and sonicated the samples prior to the procedure for sequential protein extractions. The multi-step fractionation protocol resulted in three fractions where different detergent solvents were tested in the second fraction: the first fraction (fraction 1: TBS-soluble) extracted TBS-soluble proteins; the second fraction (fraction 2: detergent-soluble) extracted either TBS-soluble (TBS-wash), SDS-soluble, or RIPA-soluble proteins; and, finally, we sonicated and resuspended the TBS-insoluble, SDS-insoluble or RIPA-insoluble pellet in urea and SDS (fraction 3: insoluble) (Figure 1A).
From the three protocols described above, we detected similar amounts of α-synuclein in the TBS-soluble fractions (fraction 1) (Figure 1B,C). However, in the second fraction, we only detected α-synuclein when solubilizing protein extracts in the SDS (SDS-soluble) or RIPA (RIPA-soluble) buffers, with a significantly higher yield in the RIPA-soluble fraction (Figure 1B,C). The second TBS fraction (fraction 2: TBS-wash) was unable to extract any α-synuclein, indicating that the TBS-soluble α-synuclein was fully extracted in the first fraction. The insoluble fractions (fraction 3) were resuspended in urea/SDS and we observed low but detectable amounts of α-synuclein in the protocols that used the SDS buffer (Table 1) for the second fraction (Figure 1B,C). However, we were not able to consistently detect α-synuclein in the insoluble fraction that followed the RIPA extraction (fraction 3: RIPA-insoluble; Figure 1B,C), with it being detectable in two of the five experiments. Together, these data indicate that expression of hSNCA in DA neurons of the fruit fly, Drosophila melanogaster, led to the accumulation of TBS-insoluble α-synuclein, which was most abundant in the detergent-containing fraction (SDS- and RIPA-soluble fraction 2) of the protocol (Table S1). In addition, the brain-accumulated α-synuclein exhibited a sufficiently high solubility in the RIPA buffer so that no additional protein was detectable in the insoluble (urea) fraction.
Next, we assessed the amount of α-tubulin, a soluble control protein, in each of these fractions. We detected most of the protein in the TBS-soluble fraction (fraction 1), with no significant differences between protocols (Figure 1B,D). Although some residual α-tubulin was detected in the other two fractions, the amounts were never significant (Figure 1D and Table S2). Therefore, α-tubulin remained in the soluble fraction irrespective of the fractionation protocol employed, in contrast to the segregation observed for α-synuclein throughout the different solubility fractions and protocols.
Furthermore, we employed the same exact protocols for control samples expressing LacZ instead of hSNCA. In this case, the specific 15kDa band that the α-synuclein antibody (4B12) detected did not appear in any fractions of head lysates (Figure 1B and Figure S1), confirming the specificity of the signal and the lack of α-synuclein in flies that do not carry the UAS-hSNCA transgene. On the other hand, the soluble control protein, α-tubulin, was detected at a similar level in all the TBS-soluble fractions but remained undetectable or at a very low level in the detergent-soluble or insoluble fractions, regardless of the protocol employed (Figure S1).
2.2. Sonication Increases α-Synuclein Solubility in SDS Buffer
Our results indicated that most of the α-synuclein from 20-day-old fly brains was detected in the detergent-soluble fraction (fraction 2), with minimal amounts being found in the insoluble (urea) fractions. This contrasts with the previous literature that reported histological evidence of highly aggregated α-synuclein in a similar Drosophila model [22], strongly suggesting the presence of insoluble α-synuclein. We then questioned whether the protocol that we adapted from the vertebrate literature had been fully optimized for Drosophila. In this protocol, we sonicated samples immediately after homogenization. Sonic cavitation produces pressure fluctuations of gigapascal magnitude, generating high-energy mechanical shockwaves that have been reported to degrade some proteins [35]. We hypothesized that this high-energy process could be disaggregating physiologically insoluble α-synuclein in the protein extracts. To investigate this possibility, we compared the distribution of α-synuclein from fly heads that were either sonicated or non-sonicated before biochemical fractionation across the three previous fractions.
We first compared the effects of sonication on α-synuclein fractionation under the protocol that employed SDS as the detergent solvent in fraction 2. Under this protocol (Figure 2A), we did not observe significant differences between sonicated and non-sonicated samples for the TBS-soluble and SDS-soluble fractions (Figure 2B,C). However, we found a significant increase of about two-fold in the amount of insoluble α-synuclein detected in the SDS-insoluble fraction (fraction 3) without sonication when compared to the sonicated samples (Figure 2B,C). We then evaluated the distribution of α-tubulin across these three fractions upon either sonication or non-sonication. Regardless of sonication, α-tubulin was still predominantly present in the TBS-soluble fraction with almost undetectable traces of the protein in the SDS-soluble and SDS-insoluble fractions (Figure 2B,D). These data indicated that sonication did not influence cell lysis or protein extraction efficiency as the levels of α-tubulin were equivalent for both protocols, sonicated and non-sonicated. In contrast, these results support the conclusion that sonication affects α-synuclein solubility, perhaps by breaking down the aggregates. This could explain why the sonicated samples herein were found mostly in the SDS-soluble fraction, while the non-sonicated samples exhibited higher levels of α-synuclein in the SDS-insoluble fraction.
2.3. Sonication Does Not Affect α-Synuclein Solubility in RIPA Buffer
We next tested whether sonication elicited a similar effect on the fractionation of α-synuclein when the buffer employed was RIPA. Interestingly, the amount of α-synuclein in the three fractions (Figure 3A) was statistically equivalent when comparing sonicated versus non-sonicated samples (Figure 3B,C). The results revealed that α-synuclein was still undetectable in the RIPA-insoluble fraction, while the RIPA-soluble fraction retained most of the protein (Figure 3B,C). Therefore, we did not observe any changes due to sonication when using RIPA buffer as the buffer in fraction 2 (Figure 3B,C). We then assessed α-tubulin and observed similar results to those obtained upon sonication. There were no significant changes between sonicated and non-sonicated samples for any of the fractions, while α-tubulin was predominantly present in the TBS-soluble fraction and almost undetectable in the RIPA-soluble and RIPA-insoluble fractions (Figure 3B,D). Together, these data indicated that sonication has no effect on α-synuclein fractionation when using RIPA buffer and suggested a superior extraction efficiency of the detergent solvents contained in this buffer when evaluating the solubility of α-synuclein, irrespective of its aggregation state due to sonication.
2.4. α-Synuclein Is Fully Soluble in Polyethoxylate Detergents Irrespective of Sonication
Given the discrepancy between the SDS and RIPA buffers regarding α-synuclein solubility upon sonication, we decided to further investigate the chemical properties underlying this behavior. Since the RIPA buffer is composed of sodium deoxycholate, NP-40, a polyethoxylated detergent, and sodium dodecyl sulfate (SDS) at a concentration 50X lower than the SDS buffer, we posed the question of whether polyethoxylated detergents could be conferring the differential properties of RIPA versus SDS-alone solubilization. To address this, we performed an experiment using a similar three-fraction protocol (Figure 4A) with NP-40 as the detergent solvent in the buffer employed in the second fraction and compared the effects of sonication. Similar to what we observed for the protocol with the RIPA buffer, α-synuclein was undetectable in the NP-40-insoluble fraction (sonicated and non-sonicated), while the fraction with the most α-synuclein was the NP-40-soluble fraction (Figure 4B,C). The amount of α-synuclein in each fraction was statistically equivalent regardless of whether or not the samples were previously sonicated (Figure 4B,C). We also evaluated α-tubulin and obtained the exact same results as for RIPA or SDS as the detergent solvents (Figure 2 and Figure 3): mostly present in the TBS-soluble fraction but undetectable or at a very low level for NP-40-soluble and NP-40-insoluble fractions (Figure 4B,D). Together, these results led us to conclude that NP-40, a polyethoxylated detergent, is responsible for the high efficiency with which α-synuclein is solubilized in RIPA buffer irrespective of sonication before fractionation.
Lastly, we tested whether another polyethoxylated detergent commonly used for fractionation of α-synuclein in vertebrate animal models, Triton X-100, conferred the same chemical properties as NP-40 regarding α-synuclein solubility. In this case, we employed a two-step protocol (Figure 5A), containing Triton X-100-soluble and Triton X-100-insoluble fractions. Similarly, α-synuclein was undetectable in the Triton X-100-insoluble fraction regardless of whether the samples were previously sonicated, remaining entirely in the X-Triton-soluble fraction for the sonicated and non-sonicated samples (Figure 5B,C). We also evaluated α-tubulin, which was only detected in the Triton X-100-soluble fraction (Figure 5B,D). In summary, the solubility of α-synuclein in polyethoxylated detergents is such that these detergents sequester all α-synuclein present in the protein extracts from Drosophila heads, regardless of whether they have been previously sonicated or not. In our opinion, fractionation buffers containing SDS alone uniquely provide sufficient dynamic range for evaluation of the solubility state of human α-synuclein expressed in Drosophila brains with high resolution.
3. Discussion
Chemical fractionation of misfolded and aggregated proteins is a critical method to assess the solubility and the species composition of the proteinaceous deposits formed by these proteins and their relationship to disease stage. We tested a variety of protocols to evaluate the solubility of hSNCA expressed in the DA neurons of the Drosophila brain and found that buffers containing polyethoxylated detergents such as NP-40 or Triton X-100 exert such a strong extraction efficiency for TBS-insoluble α-synuclein that no α-synuclein remains in the insoluble fraction. However, SDS-containing solvents only solubilize a fraction of the remaining α-synuclein, leaving the most insoluble molecules in the precipitate. We also observed that sonication prior to chemical fractionation breaks down insoluble aggregated α-synuclein species, making them soluble in the SDS-containing fraction. Below, we consider several possible explanations for this biochemical behavior of human α-synuclein.
First, we looked at the chemical properties of these detergent solvents seeking to find the basis for the difference in extraction efficiency. SDS is an anionic detergent that consists of a short acyl chain attached to a sulfate head group. On the other hand, RIPA is a composite detergent buffer that contains 1% NP-40 (a nonionic polyethoxylate detergent with 24–51 ethyl ether groups per molecule), 1% sodium deoxycholate (an anionic detergent that consists of a steroid core attached to a carboxylate head group), and 0.1% SDS. At these concentrations, all these molecules form micelles (critical micelle concentrations: SDS—0.079% w/v [36], NP-40—0.035% w/v [37], sodium deoxycholate—0.099% w/v [38]) that encapsulate the hydrophobic portions of proteins to various degrees, extracting them into a solution. We suspect that the presence of NP-40 is responsible for the higher content of α-synuclein in the RIPA-soluble fractions, given that the use of NP-40 alone mimicked the results obtained with RIPA. With 24–51 ethyl ether groups [37], NP-40 molecules can be 87–168 Å long, which is 7–14 times longer than an SDS molecule (Figure 6). Moreover, the average number of molecules per micelle (as measured by the aggregation number, N_A_) for NP-40 is around 150 [39], more than twice as many as SDS [N_A_ = 67; [40]]. As a result of these biophysical properties, NP-40 forms larger and more flexible micelles, which we hypothesize are more capable of encapsulating and extracting larger α-synuclein complexes than the smaller SDS micelles. Therefore, RIPA buffer as a solubilization solvent may lead to an underestimation of the insoluble human α-synuclein content due to its higher capacity to sequester larger insoluble α-synuclein aggregates. We tested this hypothesis by extracting α-synuclein using a buffer with 1% NP-40 as the detergent and found that NP-40-soluble fractions were highly enriched in α-synuclein, while α-synuclein was almost undetectable in NP-40-insoluble fraction. These data recapitulate the results we observed when using RIPA, consistent with our hypothesis.
Another detergent that is commonly used in the field to assess insoluble α-synuclein levels is Triton X-100, a nonionic polyethoxylate detergent very similar to NP-40. Earls et al. (2020) used suspensions of Triton X-100-insoluble material to measure the uptake of aggregated α-synuclein in human NK cell cultures from media containing pre-formed fibrils (PFFs) [41]. In an independent study, Quintin et al. used the same approach to assess α-synuclein aggregation in HEK cells incubated with PFFs [6]. To the best of our knowledge, Triton X-100 is the only detergent used for α-synuclein biochemical fractionation in the Drosophila literature. Miura et al., Suzuki et al., Davis et al. and Khair et al. [17,19,26,27] used Triton X-100-insoluble α-synuclein as a measure of protein aggregation. Suzuki et al. obtained a very weak α-synuclein monomer signal in the insoluble fraction, many times weaker than soluble α-synuclein signal, with no higher-molecular-weight insoluble α-synuclein species detected [27]. Miura et al. did not detect any insoluble α-synuclein monomers but reported detecting a weak α-synuclein signal at 140 kDa [26]. Similarly, Davis et al. also reported their ability to detect soluble but not insoluble α-synuclein monomers, while they observe higher-molecular-weight species in the insoluble fraction [19]. We were unable to detect Triton X-100-insoluble α-synuclein. The undetectability of Triton X-100-insoluble α-synuclein in our expression system is consistent with the rest of our experimental findings herein. As mentioned earlier, Triton X-100 is a polymer detergent with repeating ethyl ether groups. It differs from NP-40 only in the alkyl group attached to the polyethoxylated phenyl ring (tetramethylbutyl in TritonX and nonyl in NP-40) and the fact that is has a fewer number of ethyl ether repeats. Therefore, we would expect Triton X-100 to form micelles that are similar in their physical and chemical properties to those formed by NP-40, thus extracting α-synuclein with a similar efficiency. The undetectability of detergent-insoluble α-synuclein in RIPA buffer, NP-40- and Triton X-100-containing solvents suggests that the repeating ethyl ether groups are responsible for the higher solubilization of α-synuclein in buffers containing these detergents.
One potential reason for not detecting detergent-insoluble α-synuclein, which differs from previously published data, may be the lower overall expression of human α-synuclein in our model; the aforementioned Drosophila studies expressed human α-synuclein to higher levels, either pan-neuronally with elav or ubiquitously with actin promoters, while we intentionally restricted the expression to DA neurons (~130 per protocerebrum hemisphere), which constitutes only a small percentage of all the neurons in the fly brain (<200,000 neurons). It is likely that α-synuclein is not processed in the same way in every neuronal type and, therefore, we focused our study on a more physiologically relevant neuronal population, DA neurons, which are widely known to develop α-synuclein pathology and degenerate in PD. In addition, it is possible that our driver is not only more selective, but it may also drive weaker expression than other ubiquitous promoters such as actin. The relative abundance of α-synuclein could increase the aggregation rate, as mutations that lead to higher expression levels of α-synuclein trigger aggregation and LB pathology [12,13,14], with an overall higher total accumulation of aggregated α-synuclein leading to more robust and resistant detection of aggregated α-synuclein via standard biochemical fractionation and detection methods.
An additional critical factor to keep in mind when performing these fractionation experiments is whether to perform sample sonication before the serial fractionation. In our experiments using SDS as the detergent solvent in the fractionation buffer, sonication of the fly head homogenates before biochemical fractionation induced an enrichment in α-synuclein content in the SDS-soluble fraction while reducing the amount of total α-synuclein in the SDS-insoluble fraction. These findings demonstrate that sonication is sufficient to solubilize physiologically insoluble α-synuclein accumulated in Drosophila DA neurons and suggest that, to a certain degree, sonication triggers the disaggregation of α-synuclein complexes. For this reason, we would recommend that, when using Drosophila to model synucleinopathies, sonication before chemical fractionation should be avoided to prevent confounding the interpretation of its solubility state in vivo. Contrary to the need for sonication in samples from vertebrate animals that are rich in myelin and other fatty components, fly samples do not require sonication to break down those additional components present in vertebrates. Therefore, the use of sonication prior to serial fractionation is unnecessary and likely undesirable as it will compromise the interpretation of the biophysical and biochemical properties of the α-synuclein species that are being extracted and studied.
Regardless of the protocol, we could not detect solubilized α-synuclein of a higher molecular weight than that of recombinant α-synuclein monomeric protein. It is important to note that western blots were not conducted under completely native conditions. However, protein lysate samples were not reduced with BME or boiled, but they were diluted with SDS-containing Laemmli buffer prior to loading in the native gel, which likely had an effect on the protein’s natural structure and may account for the monomeric weight. Nevertheless, the limitations of the model in detecting higher molecular weight species are likely due to the low amount of α-synuclein that we find in the brain of these flies, as discussed earlier. However, it is also possible that Drosophila DA neurons expressing hSNCA do not produce high-molecular-weight species like those found in LBs, which typically require tough solvents such as urea or acid to solubilize the large α-synuclein aggregates [42,43].
Importantly, the choice of detergent for biochemical fractionation depends on the research question and the conditions suitable for the model. In cases when the detergent-soluble protein content is used as a measure of membrane-associated protein, our data demonstrate that SDS is a better detergent for these purposes. SDS, being a fatty acid similar in length to a typical phospholipid, resembles the plasma membrane more than RIPA (NP-40) or Triton X-100 because they contain considerably longer polymer detergents. In addition, extraction with SDS allows for a more direct comparison to the existing literature in human postmortem brains when assessing α-synuclein solubility, which has mainly employed SDS as a detergent in the serial biochemical fractionation of α-synuclein [15,16,43,44,45]. In conclusion, when selecting an appropriate fractionation protocol for Drosophila studies, our findings support the use of a three-step fractionation protocol where the buffer for the second fraction contains only SDS at a 5% concentration. The combination of a buffer with these biochemical properties and the multi-fraction protocol provides a dynamic range capable of resolving several degrees of insolubility in human α-synuclein processed in DA neurons of the Drosophila brain.
4. Materials and Methods
4.1. Fly Strains
Fly stocks were raised on standard corn syrup–soy (W1) food (Bloomington stock center recipe) from Archon Scientific (Durham, NC, USA). All crosses were maintained at 25 °C and 70% relative humidity on a 12 h light–dark cycle. Flies carrying the TH-Gal4 driver were provided by Dr. Matthew Kayser from the University of Pennsylvania. Expression lines UAS-LacZ (BDSC, catalog #8530; RRID:BDSC_8530) and UAS-hSNCA^A53T^ (BDSC, catalog #8148; RRID:BDSC_8148) were obtained from the Bloomington Stock center (Bloomington, IN, USA). The line for the expression of the human α-synuclein bearing the pathology-associated mutation A53T (hSNCA^A53T^) was previously described [22]. Flies were collected immediately upon eclosion and housed in individual vials in groups of 20 males. All experiments were performed on flies aged to 20 days post-eclosion.
4.2. Tissue Collection and Fractionation
4.2.1. Three-Step Fractionation Protocol
Groups of 20 male flies were transferred to Eppendorf cryovials, snap-frozen in liquid nitrogen and then vortexed. Then, 20–40 fly heads were collected into BioMasher^®^ II Tissue Grinder (DWK Life Sciences; Wertheim, Germany; 749625–0020) tubes and homogenized for 1.5 min in 50 μL of Tris-buffered saline (TBS; Table 1) with 1X protease and 1X phosphatase inhibitor cocktails (Roche; Basel, Switzerland; 04693124001 and 04906837001). The homogenates were then probe-sonicated with a Branson Sonifier^®^ SFX150 (Emerson; Emerson, MO, USA) at 35% amplitude (~52.5 watts) for five 1 s pulses, unless otherwise specified. The suspension was clarified by 15 s centrifugation at 2100× g and the supernatant was ultracentrifuged at 100,000× g for 30 min at 4 °C using the Sorvall WX80 Plua Ultracentrifuge (ThermoScientific; Waltham, MA, USA). The supernatant was labelled as the TBS-soluble fraction (fraction1). To ensure complete removal of soluble material, the pellet was washed in TBS and ultracentrifuged again at 100,000× g for 15 min at 4 °C, after which the supernatant was discarded. The pellet was then re-homogenized in 30 μL of either SDS, NP-40, RIPA, or TBS buffer (Table 1) with protease and phosphatase inhibitors and ultracentrifuged at 100,000× g for 30 min at 4 °C. The supernatant (fraction 2) was collected and labelled as SDS-soluble, NP-40-soluble, RIPA-soluble or TBS-wash, respectively. The remaining pellet was washed in the previous solvent and centrifuged again at 100,000× g for 15 min at 4 °C, after which the supernatant was discarded. The final pellet was resuspended in 30 μL urea/SDS (Table 1) with protease and phosphatase inhibitors by probe-sonication with three 1 s pulses at 35% amplitude and the suspension was labelled as the insoluble fraction (fraction 3; Figure 7A).
4.2.2. Two-Step Fractionation Protocol
Groups of 20 male flies were transferred to Eppendorf cryovials, snap-frozen in liquid nitrogen and then vortexed. Then, 20–40 fly heads were collected into BioMasher^®^ II Tissue Grinder (DWK Life Sciences; Wertheim, Germany; 749625–0020) tubes and homogenized for 1.5 min in 50 μL of TBS containing 1% Triton X-100 (Table 1) and 1× protease and 1× phosphatase inhibitor cocktails (Roche; Basel, Switzerland; 04693124001 and 04906837001). The homogenates were then probe-sonicated with a Branson Sonifier^®^ SFX150 (Emerson; Emerson, MO, USA) at 35% amplitude (~52.5 watts) for five 1 s pulses, unless otherwise specified. The suspension was clarified by 15 s centrifugation at 2100× g and the supernatant was ultracentrifuged at 100,000× g for 30 min at 4 °C using the Sorvall WX80 Plua Ultracentrifuge (ThermoScientific; Waltham, MA, USA). The supernatant was labelled as the Triton X-100-soluble fraction (fraction1). To ensure complete removal of soluble material, the pellet was washed in the Triton X-100 buffer (Table 1) and centrifuged again at 100,000× g for 15 min at 4 °C, after which the supernatant was discarded. The final pellet was resuspended in 50 μL of the Triton X-100 buffer (Table 1) with protease and phosphatase inhibitors by probe-sonication with three 1 s pulses at 35% amplitude and the suspension was labelled as the Triton-X-100-insoluble fraction (fraction2; Figure 7B).
4.3. Quantitative Western Immunoblotting
The total protein concentration in each fraction was determined by bicinchoninic acid assay (BCA) (Thermo Fisher Scientific; Waltham, MA, USA; 23225). Then, 20 μg of total protein per fraction was collected and diluted with its respective solvents and 4X Laemmli buffer (Biorad; Hercules, CA, USA; 1610747) to obtain aliquots of equal volume. These samples were loaded into the 10% Bis-Tris polyacrylamide gel (BioRad; Hercules, CA, USA; 3450112) and subjected to electrophoresis at 80 V for 15 min and 100 V for 90 min. After separation, protein was transferred via a semi-wet electrophoretic transfer (Turbo-blot turbo transfer system, Bio-Rad) onto 0.2 μm polyvinylidene fluoride (PVDF) membranes (BioRad; Hercules, CA, USA; 1704273). The PVDF membranes were fixed with 0.8% paraformaldehyde for 30 min, washed with H_2_O, and then stained with Revert Total Protein Stain (LI-COR, 926–11016) and the signal was used for signal normalization. Membranes were then destained and blocked with 5% powdered milk in TBST buffer, following an overnight incubation at 4 °C with the primary antibody against residues 103–108 of human α-synuclein (BioLegend; San Diego, CA, USA; 807801 [4B12] 1:1000; RRID: AB_2564730). The next day, HRP-conjugated secondary antibody (Jackson Immuno Research; West Grove, PA, USA; 115–036-072; RRID: AB_2338525) was incubated for 1 h at room temperature, after which the protein bands were visualized using enhanced chemiluminescence (Thermo Fisher Scientific; Waltham, MA, USA; 34096) on a LI-COR Odyssey^®^ Fc. The membranes were then stripped from the secondary antibody with 0.2 M NaOH, re-blocked, and re-probed with the anti-tubulin primary antibody (Sigma-Aldrich, T6074 1:200,000; RRID: AB_477582, St. Louis, MO, USA) and the HRP-conjugated secondary antibody, after which they were re-imaged.
4.4. Experimental Design and Statistical Analyses
For all experiments, controls and genetically matched experimental genotypes were performed in parallel. The experimental design ensured that all groups were balanced throughout the experiments conducted. Statistical significance was assessed using GraphPad Prism v10.1 (La Jolla, CA, USA) (RRID:SCR_002798). Control flies were balanced for the presence of pUAS constructs by expressing the UAS-LacZ in place of the UAS-hSNCA^A53T^ of the experimental groups. Across all gels, a previously tested sample of known antibody reactivity was run as the internal control sample that enabled comparison across blots. The raw protein band signal was first normalized to total protein staining (Figures S1–S6) to control for loading differences and then reported as the fold difference from the internal control sample signal. Ordinary two-way ANOVA tests followed by Tukey’s multiple post hoc comparisons were employed to analyze statistical significance across fractions and solvents. Student’s t-test was used to assess significance from zero and determine whether there were significant levels of α-synuclein in the fraction. All data presented represent mean ± the SD.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Goedert M. Spillantini M.G. Lewy Body Diseases and Multiple System Atrophy as Alpha-Synucleinopathies Mol. Psychiatry 1998346246510.1038/sj.mp.40004589857966 · doi ↗ · pubmed ↗
- 2Tong J. Wong H. Guttman M. Ang L.C. Forno L.S. Shimadzu M. Rajput A.H. Muenter M.D. Kish S.J. Hornykiewicz O. Brain Alpha-Synuclein Accumulation in Multiple System Atrophy, Parkinson’s Disease and Progressive Supranuclear Palsy: A Comparative Investigation Brain 201013317218810.1093/brain/awp 28219903734 · doi ↗ · pubmed ↗
- 3Parra-Rivas L.A. Madhivanan K. Aulston B.D. Wang L. Prakashchand D.D. Boyer N.P. Saia-Cereda V.M. Branes-Guerrero K. Pizzo D.P. Bagchi P. Serine-129 Phosphorylation of α-Synuclein Is an Activity-Dependent Trigger for Physiologic Protein-Protein Interactions and Synaptic Function Neuron 202311140064023.e 1010.1016/j.neuron.2023.11.02038128479 PMC 10766085 · doi ↗ · pubmed ↗
- 4Marotta N.P. Ara J. Uemura N. Lougee M.G. Meymand E.S. Zhang B. Petersson E.J. Trojanowski J.Q. Lee V.M.-Y. Alpha-Synuclein from Patient Lewy Bodies Exhibits Distinct Pathological Activity That Can Be Propagated In Vitro Acta Neuropathol. Commun.2021918810.1186/s 40478-021-01288-234819159 PMC 8611971 · doi ↗ · pubmed ↗
- 5Ma L. Yang C. Zhang X. Li Y. Wang S. Zheng L. Huang K. C-Terminal Truncation Exacerbates the Aggregation and Cytotoxicity of α-Synuclein: A Vicious Cycle in Parkinson’s Disease Biochim. Biophys. Acta Mol. Basis Dis.201818643714372510.1016/j.bbadis.2018.10.00330290273 · doi ↗ · pubmed ↗
- 6Quintin S. Lloyd G.M. Paterno G. Xia Y. Sorrentino Z. Bell B.M. Gorion K.-M. Lee E.B. Prokop S. Giasson B.I. Cellular Processing of α-Synuclein Fibrils Results in Distinct Physiological C-Terminal Truncations with a Major Cleavage Site at Residue Glu 114J. Biol. Chem.202329910491210.1016/j.jbc.2023.10491237307916 PMC 10404685 · doi ↗ · pubmed ↗
- 7Moon S.P. Balana A.T. Galesic A. Rakshit A. Pratt M.R. Ubiquitination Can Change the Structure of the α-Synuclein Amyloid Fiber in a Site Selective Fashion J. Org. Chem.2020851548155510.1021/acs.joc.9b 0264131809571 PMC 7097865 · doi ↗ · pubmed ↗
- 8Burai R. Ait-Bouziad N. Chiki A. Lashuel H.A. Elucidating the Role of Site-Specific Nitration of α-Synuclein in the Pathogenesis of Parkinson’s Disease via Protein Semisynthesis and Mutagenesis J. Am. Chem. Soc.20151375041505210.1021/ja 513172625768729 · doi ↗ · pubmed ↗