Draft genome sequences of two β-glucuronidase positive strains of Salmonella enterica subspecies salamae isolated from reptile feces in KwaZulu-Natal, South Africa
Maike Claussen, Stefan Schmidt

TL;DR
This study reports the genome sequences of two Salmonella strains from reptile feces in South Africa that show β-glucuronidase activity.
Contribution
The novelty lies in the genome sequencing and identification of two β-glucuronidase positive Salmonella salamae strains from reptiles.
Findings
Two Salmonella isolates from reptile feces showed β-glucuronidase activity.
Genome sequencing identified them as Salmonella enterica subspecies salamae with sequence type 1208 and serotype 42:r:-.
Abstract
Two Salmonella enterica isolates obtained from reptile feces displayed β-glucuronidase activity. Nearly complete genome sequences were obtained after shotgun sequencing and de novo genome assembly. By comparison to reference genomes, both isolates were identified as Salmonella enterica subspecies salamae with the sequence type identified as 1208 and the serotype as 42:r:-.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
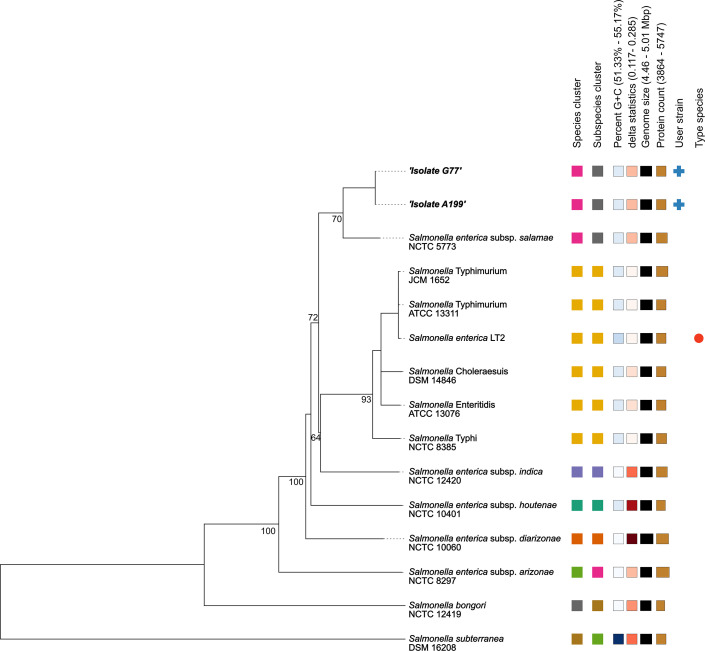 Fig 1
Fig 1| Strain | A199 | G77 |
|---|---|---|
| Origin, year | Agama feces, 2019 | Gecko feces, 2021 |
| Location | Pietermaritzburg, South Africa (29°36'S 30°23'E) | Pietermaritzburg, South Africa (29°36'S 30°23'E) |
| Identity | ||
| Digital DNA:DNA hybridization (dDDH, d4) best type strain genome match | ||
| Average nucleotide identity (ANIb) best type strain genome match | ||
| Predicted serotype | 42:r:- | 42:r:- |
| MLST sequence type | 1208 | 1208 |
| Total genome size | 4,733,636 bp | 4,733,809 bp |
| Completeness (BUSCO) | 97.97% | 97.97% |
| Number of reads (input) | 10,679,960 | 7,179,091 |
| Average read length | 140 | 137 |
| Number of contigs | 59 | 59 |
| N50 | 226,585 | 226,585 |
| Largest contig | 417,545 | 417,720 |
| GC content | 51.96% | 51.96% |
| Protein coding sequences (CDS) | 4,715 | 4,716 |
| tRNA | 73 | 73 |
| rRNA | 3 | 3 |
| Predicted resistance genes | aac(6′)-Iaa | aac(6′)-Iaa |
| Salmonella pathogenicity islands | SESS-LEE, SPI-1, SPI-2, SPI-3, SPI-9 | SESS-LEE, SPI-1, SPI-2, SPI-3, SPI-9 |
| Human pathogen probability | 0.926 | 0.926 |
| Number of predicted intact prophages | 1 | 1 |
| Plasmids | Not detected | Not detected |
- —National Research Foundation (NRF)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsSalmonella and Campylobacter epidemiology · Genomics and Phylogenetic Studies · Bacteriophages and microbial interactions
ANNOUNCEMENT
β-Glucuronidase activity is typically present in Salmonella enterica subspecies diarizonae but can also occur in the subspecies enterica, salamae, and indica (1). Although human salmonellosis’ is mostly caused by Salmonella enterica subspecies enterica, infections by other non-enterica subspecies do occur but are usually rare (2–6). Salmonella enterica subspecies salamae, like all non-enterica subspecies, are typically associated with cold-blooded animals such as reptiles but can also be found in warm-blooded animals (3, 6–9).
The two Salmonella isolates, designated A199 and G77, were originally obtained from agama (2019) and gecko feces (2021). Fecal reptile pellets collected in a suburban garden in Pietermaritzburg, South Africa, were decimally diluted in sterile saline (0.85%). Subsamples of agama fecal dilutions (100 µL) were directly spread-plated onto XLD-Agar (24 h, 37°C). A portion of the pooled gecko feces dilution was pre-enriched in buffered peptone water (24 h, 37°C), followed by selective enrichment in Rappaport-Vassiliadis broth (41.5°C, 24 h) and spread-plating on XLD-Agar (37°C, 24 h). A purified (Nutrient agar) single colony was sub-cultivated in Nutrient broth to prepare glycerol stock cultures for storage at −20°C. Both isolates were confirmed using TSI- and Urea-Agar and by PCR-based detection of the invA gene (10). Sanger sequencing (CAF, Stellenbosch) of the 16S rRNA gene amplicon (primers fD1 + rP2) (11) further confirmed that both isolates belonged to the species Salmonella enterica (accession no. ON630378.1, ON630388.1). Notably, both isolates showed untypical colony morphology on improved Salmonella-Chromo-Select-Agar due to their β-glucuronidase activity, which was confirmed using TBX agar.
Washed cells of pure overnight cultures of A199 and G77 were transferred into DNA/RNA Shield Lysis Tubes (ZYMO). Extracted DNA (ZymoBIOMICS-96 MagBead kit) was used for shotgun genome sequencing (paired-end 2 × 150 bp, Illumina Nova Seq) by Zymo Research (Irvine, CA, USA) after creating a library (Nextera DNA Flex Library Prep Kit). Using the BV-BRC genome analysis pipeline (12), raw reads were quality filtered, trimmed (Trim Galore v0.6.5, Cutadapt v2.2), assembled (Unicycler v0.4.8, minimum contig size 500 bp), and annotated (RASTtk) (13–16). Default parameters were used for all bioinformatic tools unless otherwise stated. Resistance genes were detected using Resfinder (v.4.1) (17), prophages using Phaster (18), plasmids using Plasmidfinder (v2.1, https://cge.cbs.dtu.dk), and human pathogenicity was predicted using Pathogenfinder (v1.1) (19).
The results for the main genomic features of both isolates are summarized in Table 1. The genome assembly was estimated to be 97.97% complete using the universal single-copy ortholog benchmarking tool available at http://cab.cc.spbu.ru (20). Using ANIb (21) and the phylogenetic analysis tools available in TYGS (22) demonstrated that the two isolates (G77, A199) belonged to Salmonella enterica subspecies salamae (Fig. 1). Multilocus sequence typing (MLST v2.0) (23) identified both isolates as sequence type 1208, and the serotype was predicted as 42:r:- (SeqSero2) (24).
Midpoint-rooted phylogenetic tree inferred with FastME (2.1.6.1) from GBDP distances calculated from type strain genome sequences using the TYGS server. Branch lengths are scaled based on the GBDP distance formula d5; numbers shown are GBDP pseudo-bootstrap support values > 60 % (100 replications, average branch support = 63%).
Our results show that Salmonella enterica subspecies salamae strains of the same serotype (42:r:-) are prevalent in the feces of geckos and agama lizards in South Africa. As both Salmonella isolates showed a high probability of human pathogenicity, feces shed by these reptiles might be a source of Salmonella contamination and a potential health risk, especially for the most vulnerable YOPI (young-old-pregnant-immunocompromised) citizens.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Grimont PAD, Weill F-X. 2007. Antigenic formulae of the Salmonella serovars, p 1–166. In WHO collaborating centre for reference and research on Salmonella. Vol. 9.
- 2Schrire L, Crisp S, Bear N, Mc Stay G, Koornhof HJ, Le Minor L. 1987. The prevalence of human isolates of Salmonella subspecies II in southern Africa. Epidemiol Infect 98:25–31. doi:10.1017/s 09502688000616893549338 PMC 2235276 · doi ↗ · pubmed ↗
- 3Aleksic S, Heinzerling F, Bockemühl J. 1996. Human infection caused by Salmonellae of subspecies II to VI in Germany, 1977-1992. Zentralbl Bakteriol 283:391–398. doi:10.1016/s 0934-8840(96)80074-08861877 · doi ↗ · pubmed ↗
- 4Editorial team, Bertrand S, Rimhanen-Finne R, Weill FX, Rabsch W, Thornton L, Perevoščikovs J, van Pelt W, Heck M. 2008. Salmonella infections associated with reptiles: the current situation in Europe. Euro Surveill 13:18902. doi:10.2807/ese.13.24.18902-en 18761944 · doi ↗ · pubmed ↗
- 5Nair S, Wain J, Connell S, de Pinna E, Peters T. 2014. Salmonella enterica subspecies II infections in England and Wales – the use of multilocus sequence typing to assist serovar identification. J Med Microbiol 63:831–834. doi:10.1099/jmm.0.072702-024696514 · doi ↗ · pubmed ↗
- 6Lamas A, Miranda JM, Regal P, Vázquez B, Franco CM, Cepeda A. 2018. A comprehensive review of non-enterica subspecies of Salmonella enterica. Microbiol Res 206:60–73. doi:10.1016/j.micres.2017.09.01029146261 · doi ↗ · pubmed ↗
- 7Lamas A, Regal P, Cepeda A, Franco CM. 2021. Complete genome sequences of six Salmonella enterica strains (S. enterica subsp. arizonae, S. enterica subsp. diarizonae, and S. enterica subsp. salamae) isolated from poultry houses. Microbiol Resour Announc 10:e 00768-21. doi:10.1128/MRA.00768-2134647803 PMC 8515881 · doi ↗ · pubmed ↗
- 8Crump JA, Thomas KM, Benschop J, Knox MA, Wilkinson DA, Midwinter AC, Munyua P, Ochieng JB, Bigogo GM, Verani JR, Widdowson M-A, Prinsen G, Cleaveland S, Karimuribo ED, Kazwala RR, Mmbaga BT, Swai ES, French NP, Zadoks RN. 2021. Investigating the meat pathway as a source of human nontyphoidal Salmonella bloodstream infections and diarrhea in East Africa. Clin Infect Dis 73:e 1570–e 1578. doi:10.1093/cid/ciaa 115332777036 PMC 8492120 · doi ↗ · pubmed ↗