Microbial diversity in shallow-sea sediment from Tsukumo Bay, Japan, determined by 16S rRNA gene amplicon sequencing
Moe Kato, Reina Sugitani, Sota Niiyama, Takahiro Kamiya, Norihisa Matsuura

TL;DR
This paper explores the microbial diversity in shallow-sea sediment from Tsukumo Bay, Japan, using 16S rRNA gene sequencing.
Contribution
The study provides new insights into the microbial community structure in Tsukumo Bay's marine sediments.
Findings
16S rRNA gene amplicon sequencing was used to assess microbial diversity across 19 sediment stations.
The study identified a diverse range of microbial taxa in the marine sediments of Tsukumo Bay.
Abstract
Information about the microbiota in marine sediments is important because the microbiota and their activities in sediments affect the surrounding marine environment. To evaluate the microbial diversity, we performed 16S rRNA gene amplicon sequencing on sediment samples from 19 stations in Tsukumo Bay, the northern area of Noto Peninsula, Japan.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
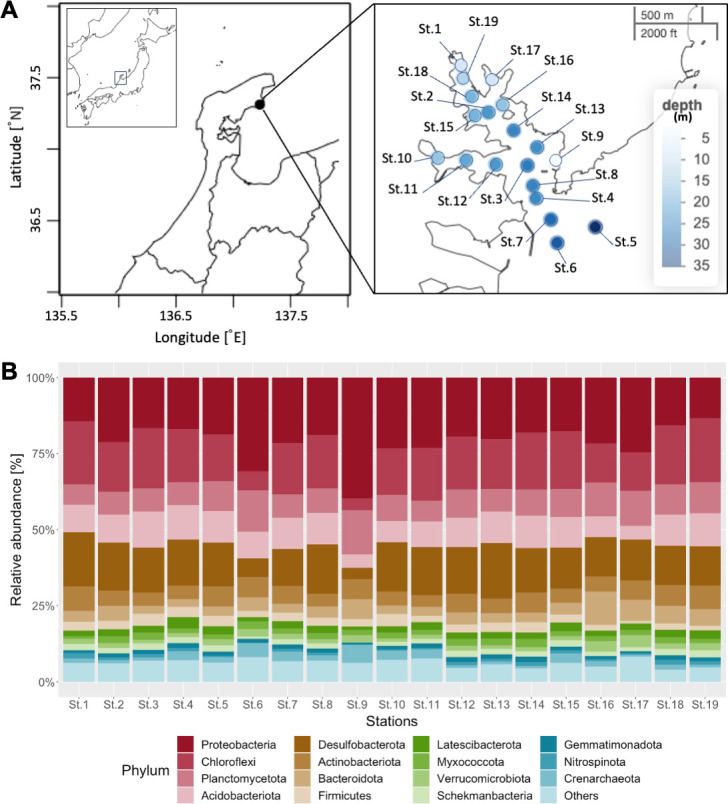 Fig 1
Fig 1| Sample ID | Latitude | Longitude | Depth (m) | Sediment type | No. of raw sequencing reads | DRA accession no. |
|---|---|---|---|---|---|---|
| Station 1 | 37.31104999°N | 137.2311828°E | 7.3 | Mud | 8,817 |
|
| Station 2 | 37.30804999°N | 137.2334169°E | 22.1 | Mud | 7,009 |
|
| Station 3 | 37.30466694°N | 137.2365669°E | 25.5 | Mud | 9,569 |
|
| Station 4 | 37.30254972°N | 137.2372497°E | 23.8 | Sandy mud | 4,964 |
|
| Station 5 | 37.30068277°N | 137.2420828°E | 35.1 | Sandy mud | 10,429 |
|
| Station 6 | 37.29966694°N | 137.2389497°E | 29.8 | Sand | 11,099 |
|
| Station 7 | 37.30116694°N | 137.23845°E | 27.6 | Sandy mud | 15,430 |
|
| Station 8 | 37.30336694°N | 137.2370169°E | 24.5 | Sandy mud | 8,463 |
|
| Station 9 | 37.30491694°N | 137.239°E | 2.2 | Sand | 5,848 |
|
| Station 10 | 37.30511694°N | 137.2293169°E | 15.3 | Mud | 4,886 |
|
| Station 11 | 37.30498277°N | 137.2315828°E | 19.6 | Sandy mud | 9,820 |
|
| Station 12 | 37.30469972°N | 137.2339669°E | 22 | Mud | 14,489 |
|
| Station 13 | 37.30583277°N | 137.2373169°E | 23.2 | Mud | 19,671 |
|
| Station 14 | 37.30686694°N | 137.2355°E | 24.3 | Sandy mud | 38,071 |
|
| Station 15 | 37.30786694°N | 137.2323°E | 17.7 | Mud | 12,064 |
|
| Station 16 | 37.30856694°N | 137.2344997°E | 15.6 | Sandy mud | 8,246 |
|
| Station 17 | 37.31018277°N | 137.2336169°E | 7.7 | Sandy mud | 8,390 |
|
| Station 18 | 37.30908277°N | 137.2320669°E | 17.7 | Mud | 5,884 |
|
| Station 19 | 37.31024999°N | 137.2313°E | 12.1 | Sandy mud | 5,721 |
|
- —MEXT | Japan Society for the Promotion of Science (JSPS)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMicrobial Community Ecology and Physiology · Methane Hydrates and Related Phenomena · Gut microbiota and health
ANNOUNCEMENT
Marine sediments contain abundant microorganisms that play important roles in biogeochemical element cycling (1–3). The diversity and distribution patterns of marine microbiota in marine sediments are significantly affected by multiple environmental factors (4). Therefore, studying the microbiota is important to understand the interrelationship between microorganisms and marine environments. Tsukumo Bay (37.305°N, 137.236°E) is an inlet on the Noto Peninsula, Japan, 1,300 m long, 250 m wide, and 25 m deep in the central region. Tsukumo Bay is maintained in a stable environment inside the bay generally because the entrance to Tsukumo Bay is relatively narrow and shallow (5), and some types of bottom sediments are distributed there. In this study, 16S rRNA gene amplicon sequencing was used to explore the diversity of microbial communities in sediment samples collected from various locations in Tsukumo Bay, Japan.
Sediment samples were collected as core samples using plastic cylinders (φ28 mm) from 19 stations (Stations 1–19) in Tsukumo Bay, the northern area of Noto Peninsula, Japan (Fig. 1; Table 1), in May 2022. The samples were stored in a cooler with ice packs for about 3 hours and transferred to the lab, where they were stored at −20°C. Surface sediments (0–2 cm in depth) were used for 16S rRNA gene amplicon sequencing. After homogenizing each sample by using a Vortex-Genie 2 (Scientific Industries, USA), total DNA was extracted using the DNeasy PowerSoil Pro kit and automatic DNA extractor QIAcube Connect (Qiagen, Germany), following the manufacturer’s protocol. Amplicon libraries were prepared using a two-step tailed PCR method (6) with 515F and 806R primers targeting the V4 region of the prokaryotic 16S rRNA gene (7). Illumina adapter sequences were attached to the 5′ end of the forward and reverse sequencing primers. The prepared libraries were sequenced on an Illumina MiSeq platform at Kanazawa University using the MiSeq Reagent Nano Kit v2 (Illumina, USA). PhiX Control v3 (Illumina) was added to the libraries at a concentration of approximately 30% (vol/vol). This yielded 208,870 reads for the 19 data sets of the 16S rRNA gene. The 2 × 250-bp paired-end reads were trimmed (options: TRUNCLENF = 200, TRUNCLENR = 170), quality-filtered, denoised, and merged. Amplicon sequence variant (ASV) inference was performed, chimeras were removed, and ASVs were generated using DADA2 v1.16.0 (8). Taxonomic assignments were performed using the SILVA 138 SSU Ref NR 99 database (9). The default settings were used for all software unless otherwise indicated.
(A) Map of the sampling stations in Tsukumo Bay (Noto Peninsula, Ishikawa, Japan). Each circle color represents the water depth at each sampling station. The maps were obtained from GSI Maps published by the Geospatial Information Authority of Japan. (B) Column chart showing the relative abundance of the top 15 microbial phyla at each sampling station.
Proteobacteria, Chloroflexi, and Planctomycetota were the most commonly identified phyla in all the sediment samples, and their total relative abundance accounted for approximately 50% (Fig. 1). Phylum Desulfobacterota, known as sulfate-reducing bacteria, also showed a high abundance (12.2%–18.3%) comparable to the above three phyla in all localities; the average was 14.8%, except for the two stations whose sediment type was sand (Stations 6 and 9). Therefore, it is suggested that the bottom-surface sediments of Tsukumo Bay are relatively reducing environment. The data in this study provide important information about the microbial communities in shallow-sea-reducing sediments.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Fuhrman JA. 2009. Microbial community structure and its functional implications. Nature 459:193–199. doi:10.1038/nature 0805819444205 · doi ↗ · pubmed ↗
- 2Liu L, Sun F, Zhao H, Mi H, He S, Chen Y, Liu Y, Lan H, Zhang M, Wang Z. 2021. Compositional changes of sedimentary microbes in the Yangtze River Estuary and their roles in the biochemical cycle. Sci Total Environ 760:143383. doi:10.1016/j.scitotenv.2020.14338333189382 · doi ↗ · pubmed ↗
- 3Lee J-H, Patil MP, Kim J-O, Woo H-E, Kim K. 2022. Diversity of microbial communities in sediment from Yeosu Bay, Republic of Korea, as determined by 16S r RNA gene amplicon sequencing. Microbiol Resour Announc 11:e 0036322. doi:10.1128/mra.00363-2235731194 PMC 9302059 · doi ↗ · pubmed ↗
- 4Currie AR, Tait K, Parry H, de Francisco-Mora B, Hicks N, Osborn AM, Widdicombe S, Stahl H. 2017. Marine microbial gene abundance and community composition in response to ocean acidification and elevated temperature in two contrasting coastal marine sediments. Front Microbiol 8:1599. doi:10.3389/fmicb.2017.0159928878754 PMC 5572232 · doi ↗ · pubmed ↗
- 5Aida M, Kanemori M, Kubota N, Matada M, Sasayama Y, Fukumori Y. 2008. Distribution and population of free-living cells related to endsymbiont a harbored in Oligobrachia mashikoi (a siboglinid polychaete) inhabiting Tsukumo Bay. Microbes Environ 23:81–88. doi:10.1264/jsme 2.23.8121558692 · doi ↗ · pubmed ↗
- 6Matsuura N, Masakke Y, Karthikeyan S, Kanazawa S, Honda R, Yamamoto-Ikemoto R, Konstantinidis KT. 2021. Metagenomic insights into the effect of sulfate on enhanced biological phosphorus removal. Appl Microbiol Biotechnol 105:2181–2193. doi:10.1007/s 00253-021-11113-433555362 · doi ↗ · pubmed ↗
- 7Caporaso JG, Lauber CL, Walters WA, Berg-Lyons D, Lozupone CA, Turnbaugh PJ, Fierer N, Knight R. 2011. Global patterns of 16S r RNA diversity at a depth of millions of sequences per sample. Proc Natl Acad Sci U S A 108:4516–4522. doi:10.1073/pnas.100008010720534432 PMC 3063599 · doi ↗ · pubmed ↗
- 8Callahan BJ, Mc Murdie PJ, Rosen MJ, Han AW, Johnson AJA, Holmes SP. 2016. DADA 2: high-resolution sample inference from Illumina amplicon data. Nat Methods 13:581–583. doi:10.1038/nmeth.386927214047 PMC 4927377 · doi ↗ · pubmed ↗