Complete genome and characteristics of cluster BC bacteriophage SoJo, isolated using Streptomyces mirabilis NRRL B-2400 in Columbia, MD
Soven Verma Kumar, Nicholas Schaffer, Zainab Bharmal, Quinn Mood, Nameerah Abdullah, Ivan Erill, Steven M. Caruso

TL;DR
This paper describes a new Streptomyces-infecting bacteriophage, SoJo, with a genome similar to others in its cluster.
Contribution
The paper introduces a newly isolated and characterized bacteriophage from the BC cluster.
Findings
SoJo has a 39 kbp genome with 71.5% GC content.
The genome is circularly permuted and similar to other BC cluster phages.
SoJo was isolated from soil in Columbia, MD.
Abstract
Here, we present bacteriophage SoJo, a siphovirus infecting Streptomyces mirabilis, with a circularly permuted genome of 39 kbp and GC content of 71.5%. Its genome length and content are similar to that of other phages in the Actinobacteriophage Database BC cluster. SoJo was isolated from soil in Columbia, MD, USA.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
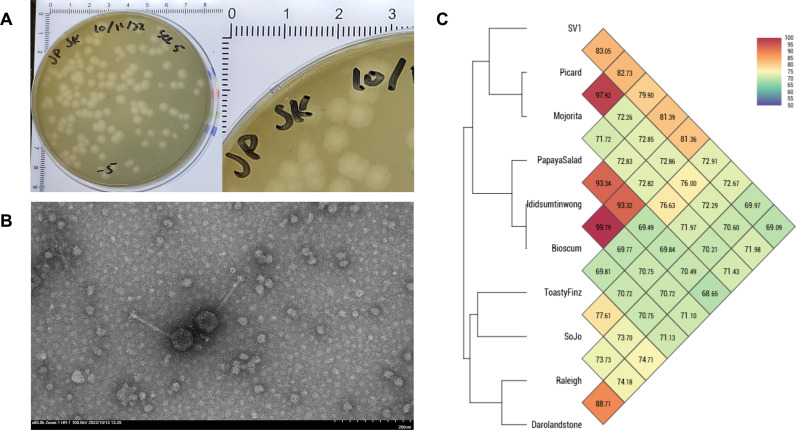 Fig 1
Fig 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBacteriophages and microbial interactions · Genomics and Phylogenetic Studies · Plant Disease Resistance and Genetics
ANNOUNCEMENT
Bacteriophages are a prospective treatment option for bacterial infections that are difficult to treat with traditional antibiotics. Typically, lytic phages are studied for their potential efficacy in these cases, but studying lysogenic phages lends greater insight into bacterial pathogenicity via phage gene transfer (1). Streptomyces bacteriophage SoJo was directly isolated from warm (25°C), dry, and coarse dirt collected from Lake Elkhorn in Columbia, MD, USA (39.182121 N, 76.843525 W) using the bacterial host Streptomyces mirabilis NRRL B-2400. The soil sample was mixed with a phage buffer (10 mM Tris, 10 mM MgSO_4_, 68 mM NaCl, and 1 mM CaCl_2_) and centrifuged for 5 minutes at 2,500 rcf at room temperature. The supernatant was filtered (0.22 µm), combined with a 48 h culture of S. mirabilis, incubated for 10 minutes mixed with tryptic soy soft agar (BD), and plated on nutrient agar (BD Difco) supplemented with 10 mM MgCl_2_, 8 mM Ca(NO_3_)2, and 0.5% glucose. All methods were adapted from the Phage Discovery Guide (2). Following plating and overnight incubation at 30°C, turbid, round plaques with a diameter of approximately 2–3 mm were observed (Fig. 1A). A plate lysate was isolated after three rounds of plaque purification. Analysis via transmission electron microscope revealed SoJo’s siphoviral morphotype with a 126 nm long, flexible, non-contractile tail (Fig. 1B).
Phage imaging and estimation of genomic homology within the BC cluster. (A) Visible plaques present after 24 hours of incubation at 30°C. Plaques shown above were typically 3 mm in diameter and turbid. (B) Transmission electron micrograph of SoJo. Fresh plate lysate was negatively stained with 2% uranyl acetate and imaged using a Hitachi HT7800 120 kV TEM with an AMT Nanosprint15 B digital camera. Measurements of three different images of SoJo show an average capsid width of 55 nm, capsid length of 57 nm, tail length of 126 nm, and a tail width of 10 nm. (C) Average nucleotide identity as determined by OAT: OrthoANI v0.93.1 between SoJo and the nine other phages in the BC cluster. Higher percentages and darker colors correspond to more closely related phage genomes.
Phage DNA was isolated from fresh viral lysates using the Promega Wizard DNA preparation kit as described previously (3). Extracted DNA was sent to the Pittsburgh Bacteriophage Institute, where samples were prepared using the NEBNext Ultra II library preparation kit (NEB #E7645) and sequenced using Illumina MiSeq with v3 reagents and 150 bp single-end reads, yielding 2,380× coverage. Sequencing reads totaling 654,326 were assembled using Newbler v2.9 with default settings and checked for accuracy using Consed v29 as described previously (4–6). Genomic termini were determined as previously described (6). SoJo was determined to have a circularly permuted genome of 39,033 bp and a genomic GC content of 71.5%.
Ab initio gene prediction was accomplished using DNAMaster v5.23.6 (7) integrated predictive algorithms Glimmer v3.02b (8) and GeneMark v4.28 (9). Automatic gene calls were refined as per SEA-PHAGES annotation guidelines (2, 7). GeneMark.hmm version 2.5p (9) with Streptomyces_scabiei_87_22 as the selected species was used to further support start site calls. Functional annotations were generated manually using BLASTP (maximum *e-*value: 10^−7^) (10), Phamerator v3.0 (11), and HHPred v57v87 (minimum probability 90%) (12, 13). Default settings were used unless noted otherwise.
Annotation revealed that SoJo’s genome contains 53 genes including loci coding for predicted integrases, endonucleases, and recombinases, suggesting that SoJo is a temperate phage. Analysis by sequence similarity using OAT: OrthoANI v0.93.1 (14) revealed that SoJo is clustered within the BC1 subcluster of phages (15), which are also identified as temperate, and has an average nucleotide identity (ANI) of 72.26% with BC1 phages (Fig. 1C). Analysis of ANI values between BC clustered phages showed that SoJo is most similar to phage ToastyFinz (accession No. KY676784).
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Gummalla VS, Zhang Y, Liao Y-T, Wu VCH. 2023. The role of temperate phages in bacterial pathogenicity. Microorganisms 11:541. doi:10.3390/microorganisms 1103054136985115 PMC 10052878 · doi ↗ · pubmed ↗
- 2Poxleitner M, Pope W, Jacobs-Sera D, Sivanathan V, Hatfull G. 2018. SEA-PHAGES Phage Discovery Guide. HHMI SEA-PHAGES Phage Discovery Guide. Howard Hughes Medical Institute, Chevy Chase.
- 3Caruso SM, De Carvalho TN, Huynh A, Morcos G, Kuo N, Parsa S, Erill I. 2019. A novel genus of actinobacterial Tectiviridae. Viruses 11:1134. doi:10.3390/v 1112113431817897 PMC 6950372 · doi ↗ · pubmed ↗
- 4Miller JR, Koren S, Sutton G. 2010. Assembly algorithms for next-generation sequencing data. Genomics 95:315–327. doi:10.1016/j.ygeno.2010.03.00120211242 PMC 2874646 · doi ↗ · pubmed ↗
- 5Gordon D, Abajian C, Green P. 1998. Consed: a graphical tool for sequence finishing. Genome Res 8:195–202. doi:10.1101/gr.8.3.1959521923 · doi ↗ · pubmed ↗
- 6Russell DA. 2018. Sequencing, assembling, and finishing complete bacteriophage genomes. Methods Mol Biol 1681:109–125. doi:10.1007/978-1-4939-7343-9_929134591 · doi ↗ · pubmed ↗
- 7Pope WH, Jacobs-Sera D. 2018. Annotation of bacteriophage genome sequences using DNA master: an overview. Methods Mol Biol 1681:217–229. doi:10.1007/978-1-4939-7343-9_1629134598 · doi ↗ · pubmed ↗
- 8Delcher AL, Bratke KA, Powers EC, Salzberg SL. 2007. Identifying bacterial genes and endosymbiont DNA with glimmer. Bioinformatics 23:673–679. doi:10.1093/bioinformatics/btm 00917237039 PMC 2387122 · doi ↗ · pubmed ↗