Endogenous fluorescent reporters for heat shock proteins are not detectable after stress induction
Priya Thirumaran, Rebecca Cornell, Roger Pocock

TL;DR
Researchers tried to create endogenous fluorescent reporters for heat shock proteins in C. elegans but found they couldn't detect the proteins after stress due to low expression.
Contribution
Generated endogenous reporters for HSP-4 and HSP-60 in C. elegans using CRISPR/Cas9, revealing detection limitations.
Findings
Endogenous reporters for HSP-4 and HSP-60 were successfully created using CRISPR/Cas9.
GFP induction was not detectable after stress induction, likely due to low expression levels.
Transgenic reporters may not fully represent endogenous HSP regulation.
Abstract
Mitochondria and the endoplasmic reticulum (ER) utilise unique unfolded protein response (UPR) mechanisms to maintain cellular proteostasis. Heat shock proteins (HSPs) are UPR chaperones induced by specific stressors to promote protein folding. Previous research has successfully employed transgenic reporters in Caenorhabditis elegans to report HSP induction. However, transgenic reporters are overexpressed and only show promoter regulation and not post-transcriptional regulation. To examine endogenous HSP regulation, we attempted to generate and validate endogenous reporters for mitochondrial ( HSP-60 ) and ER ( HSP-4 ) chaperones. Using CRISPR/Cas9 technology, F2A-GFP-H2B coding DNA was inserted downstream of each HSP gene and stress induction assays conducted to validate these tools. Endogenous reporters were successfully generated for hsp-4 and hsp-60 . However, GFP induction…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
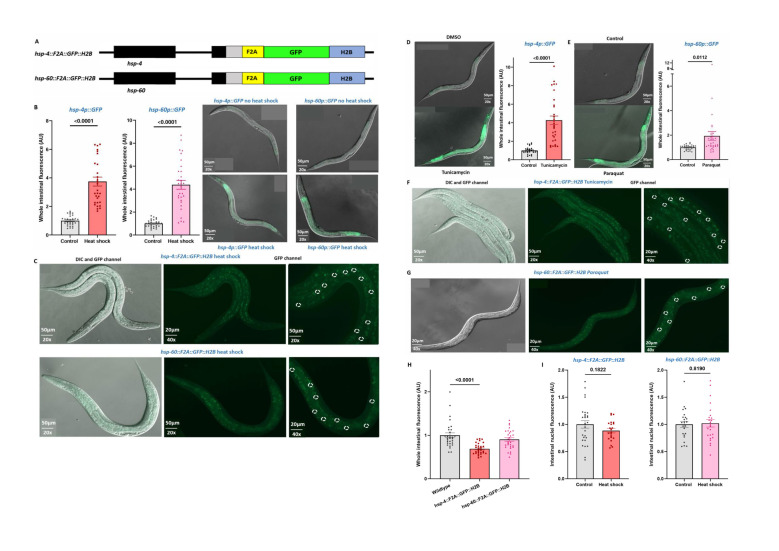 Figure 1
Figure 1|
|
|
|
|
Stress assessment |
Methyl viologen dichloride hydrate (Paraquat) |
Sigma Aldrich (856177) |
|
Tunicamycin |
Sigma (T7765-1MG) |
|
|
|
|
|
Wild-type |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Cas9 |
0.5 µl of 10 µg/µl |
|
tracrRNA |
5µl of 0.4 µg/µl |
|
crRNA |
2.8µl of 0.4 µg/µl |
|
dsDNA donor cocktail |
2.2 µl of 200 ng/µl |
|
|
1.6 µl of 4g/µl |
|
Nuclease-free water (optional) |
To bring total volume to 20µl |
|
|
|
|
|
|
Hsp-4 GFP F geno |
GAATCTGTTGTTCAACCAATCG |
|
Hsp-4/6/60 GFP R geno |
cgtcacgacttcttcaagtc | |
|
|
Hsp-60 GFP F geno |
CCAAGATGCTTCAGGAGTCG |
|
Hsp-4/6/60 GFP R geno |
cgtcacgacttcttcaagtc |
|
|
|
|
|
|
|
871 bp, 58°C |
RC
|
CAAACTTTACTCGGCGGGA |
|
RC
|
TCCGATTGGGGTGTTTTGTT | ||
|
813 bp, 58°C |
RC
|
GGAAACATCCTCGGACACAA | |
|
RC
|
GGGTTGGGTTGGGAAAGAAT | ||
|
|
977 bp, 57.5°C |
RC
|
CTCCAAGATGCTTCAGGAGT |
|
RC
|
TAGTGGTTGTCTGGGAGGAG | ||
|
731 bp, 58°C |
RC
|
TCAGATCCGTCACAACATCG | |
|
RC
|
AATGGCTCAGAGCACAAAAGA |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsEicosanoids and Hypertension Pharmacology
Description
Every organelle in eukaryotic cells performs distinct roles that are integral to normal cellular function. Proteostasis is crucial for maintaining organelle health and cellular homeostasis (Osellame et al., 2012; Moehle et al., 2019; Sengupta and Weljie, 2019) . Cellular stress generated by genetic variations, the environment, and dietary changes disrupt proteostasis and can lead to cell and tissue dysfunction (Fulda et al., 2010; Jiang et al., 2021) . To combat damage from toxic protein build-up, the mitochondria and ER utilise unique stress responses called the mitochondrial unfolded protein response (UPR ^mt^ ) and the ER unfolded protein response (UPR ^ER^ ), respectively (Fulda et al., 2010; Heldens et al., 2011) . Chaperone and protease genes are key players in mediating these stress responses. The primary roles of ER heat shock protein 4 ( HSP-4 ) and mitochondrial heat shock protein 60 ( HSP-60 ) chaperone proteins are to regulate protein quality control and folding to maintain homeostasis (Fulda et al., 2010) . Although the key chaperone proteins of cellular stress responses are widely known (Fulda et al., 2010; Heldens et al., 2011; Chovatiya and Medzhitov, 2014; Jiang et al., 2021; Inigo and Chandra, 2022) , there is currently no tool available that enables the *in vivo * visualization of endogenous UPR ^mt^ and UPR ^ER^ stress responses. To obtain further insight into the function and mechanics of cellular stress responses, we aimed to generate tools to analyse endogenous chaperone protein function and regulation. These tools would enable visualisation and quantification of endogenous mitochondrial and ER stress responses in vivo .
To investigate endogenous HSP expression, endogenous reporters for HSP-4 and HSP-60 were generated using CRISPR/Cas9 technology. The CRISPR design for *hsp-4::F2A::GFP::H2B * ( Fig. 1A, top ) and *hsp-60::F2A::GFP::H2B * ( Fig. 1A, bottom ) contained linker, F2A, GFP and H2B sequences. Both the linker sequence and F2A ribosome skipping sequence aimed to ensure that the endogenous protein function was maintained by separating the endogenous protein from the GFP fluorophore. As both HSP-4 and HSP-60 proteins are expressed ubiquitously, the histone H2B nuclear localization signal was added to direct GFP to the nuclei and thereby aid visualization. Successful in-frame insertion of the linker::F2A::GFP::H2B sequence into each gene was confirmed by Sanger sequencing prior to analysis of induction by relevant stressors.
To examine stress induction of endogenous *hsp-4::F2A::GFP::H2B * and hsp-60::F2A::GFP::H2B reporters we used treatments previously shown to induce the *hsp-4p::GFP * and *hsp-60p::GFP * transgenic reporters (Wallace et al., 2015) . First, we used a general heat shock treatment that induces expression of the transgenic *hsp-4p::GFP * and hsp-60p::GFP reporters ( Fig. 1B ). However, GFP was not visibly induced in the *hsp-4::F2A::GFP::H2B * ( Fig. 1C, top ) or *hsp-60::F2A::GFP::H2B * ( Fig. 1C, bottom ) endogenous tools following heat shock.
Next, we treated *hsp-4p::GFP * and *hsp-4::F2A::GFP::H2B * animals with the ER stress inducer tunicamycin (Guha et al., 2017) . We found that as previously described (Kim et al., 2010) , the *hsp-4p::GFP * transgenic reporter is robustly induced by tunicamycin ( Fig. 1D ). However, no induction was detected in the hsp-4::F2A::GFP::H2B endogenous reporter ( Fig. 1F ). Likewise, expression of hsp-60p::GFP is induced by the mitochondrial stressor paraquat (12) ( Fig. 1E ) but no induction was detected in the hsp-60::F2A::GFP::H2B endogenous reporter ( Fig. 1G ).
To confirm that any fluorescence observed in the hsp-4::F2A::GFP::H2B and *hsp-60::F2A::GFP::H2B * strains is due to background, we compared their fluorescence to wild-type animals when unstressed. We found that intestinal fluorescence of hsp-4::F2A::GFP::H2B and *hsp-60::F2A::GFP::H2B * animals was not greater than wild-type and thus the fluorescence detected was likely background ( Fig. 1H ). Similarly, there was no increase in intestinal nuclear fluorescence resulting from exposure to heat stress in the hsp-4::F2A::GFP::H2B and *hsp-60::F2A::GFP::H2B * strains ( Fig. 1I ).
Together, these data suggest that endogenous expression of * hsp-4 * and * hsp-60 * may be very low even when induced by a stressor. As the previously generated transgenic tools likely contain many promoter-GFP copies, protein expression is strongly intensified and can be visualised by fluorescence microscopy. The endogenous tools could be further validated by qPCR to detect *gfp * transcripts or western blotting to detect and quantify low-abundant proteins, however, the ability to visualize these proteins in vivo is at present beyond detection. Owing to the vast amount of time and resources poured into the project, we would like to share our results with the scientific community to prevent the further investment of other academics potentially attempting to generate such tools.
Methods
Genome editing of HSPs using CRISPR/Cas9
The * Alt-R ^TM^ * CRISPR HDR design tool by Integrated DNA Technologies, Inc. was used to design CRISPR sequences generated in this study. Injection mixtures for both tools were prepared according to a previous description by (Paix et al., 2015) . The components of the injection mixture, PCR primers and reaction conditions used for screening and sequencing are listed in Tables 2-4.
Microinjection, Genotyping and Sequencing
CRISPR-Cas9 injection mixes were injected into the germline of 25 individual adult wild-type worms (P0). Injected worms were transferred onto individual plates and incubated at 20°C for 3 days to produce F1 progeny. Each P0 plate was assessed for mCherry fluorescence. The plate with the greatest number of worms displaying the red injection marker was deemed a good candidate injection plate for screening. 200 F1 progeny were picked from candidate plates and F2 progeny were screened by PCR for the insertion. For each plate, a worm lysate was obtained, and genomic DNA used as a PCR template for detection of the insertion. Homozygosity of a candidate insertion strain was confirmed on 12 single animals. Following this, purified DNA from the sequencing PCR was collected and the in-frame inserted sequence was confirmed by Sanger sequencing.
Stress induction assays
All animals were maintained for three generations at 20°C prior to each stress induction assay and late L4 or early adult stage worms were primarily investigated. In each type of stress induction, transgenic *hsp-4p::GFP * and *hsp-60p::GFP * animals acted as the positive control for the assay. Animals were imaged immediately after exposure to respective stress inducers and intestinal GFP intensity was analysed. Heat shock was conducted by exposing worms to 37°C for two hours (Wallace et al., 2015) . To induce ER stress, animals were transferred onto NGM plates containing 10μg/mL tunicamycin or DMSO (control) and were incubated at 20°C for five hours prior to imaging (Kim et al., 2010) . NGM plates with tunicamycin or DMSO were prepared according to the recipe as described previously (Kim et al., 2010) . To induce mitochondrial stress, animals were transferred onto NGM plates containing 3.33M paraquat or water (control) and incubated at 20°C for five hours prior to imaging (Senchuk et al., 2017) . NGM plates with paraquat or water were prepared according to the recipe as described previously (Castello et al., 2007) .
Microscopy
Worms were anaesthetized with levamisole (0.1 ng/ml) for imaging expression of fluorescent reporters. Worms were mounted on 5% agarose pads on glass slides. Images were acquired using a Zeiss Axio Imager M2 and Zen software tools. Figures were prepared using ImageJ and Adobe Illustrator.
Reagents
**: **
** Table 1 C. elegans strains used in this study **
**: **
*Generated in this study. Abbreviations: HSP: Heat shock proteins, F2A: ribosomal skipping sequence, GFP: Green fluorescent protein; H2B: histone 2B sequence.
** Table 2 Components of CRISPR microinjection mixture ^51^ **
**: **
Table 3 PCR Primers used for screening
**: **
Abbreviations: HSP: Heat shock proteins, GFP: Green fluorescent protein, F: Forward, R: Reverse, geno: genotype, PCR: Polymerase chain reaction.
Table 4 PCR primers for sequencing
**: **
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Castello PR Drechsel DA Patel M 2007327 Mitochondria are a major source of paraquat-induced reactive oxygen species production in the brain.J Biol Chem 282190021-9258141861419310.1074/jbc.M 70082720017389593 PMC 3088512 · doi ↗ · pubmed ↗
- 2Chovatiya R Medzhitov R 2014424 Stress, inflammation, and defense of homeostasis.Mol Cell 5421097-276528128810.1016/j.molcel.2014.03.03024766892 PMC 4048989 · doi ↗ · pubmed ↗
- 3Fulda S Gorman AM Hori O Samali A 2010221 Cellular stress responses: cell survival and cell death.Int J Cell Biol 20101687-887621407421407410.1155/2010/21407420182529 PMC 2825543 · doi ↗ · pubmed ↗
- 4Guha P Kaptan E Gade P Kalvakolanu DV Ahmed H 2017715 Tunicamycin induced endoplasmic reticulum stress promotes apoptosis of prostate cancer cells by activating m TORC 1.Oncotarget 840681916820710.18632/oncotarget.1927728978108 PMC 5620248 · doi ↗ · pubmed ↗
- 5Heldens L Hensen SM Onnekink C van Genesen ST Dirks RP Lubsen NH 2011810 An atypical unfolded protein response in heat shocked cells.P Lo S One 68e 23512 e 2351210.1371/journal.pone.002351221853144 PMC 3154502 · doi ↗ · pubmed ↗
- 6Inigo JR Chandra D 2022721 The mitochondrial unfolded protein response (UPR(mt)): shielding against toxicity to mitochondria in cancer.J Hematol Oncol 151989810.1186/s 13045-022-01317-035864539 PMC 9306209 · doi ↗ · pubmed ↗
- 7Jiang S Liu H Li C 2021811 Dietary Regulation of Oxidative Stress in Chronic Metabolic Diseases.Foods 1082304-815810.3390/foods 1008185434441631 PMC 8391153 · doi ↗ · pubmed ↗
- 8Kim HM Do CH Lee DH 2010824 Taurine reduces ER stress in C. elegans.J Biomed Sci 17 Suppl 1Suppl 11021-7770 S 26S 2610.1186/1423-0127-17-S 1-S 2620804601 PMC 2994371 · doi ↗ · pubmed ↗