A new mouse model highlights the need for better JAK inhibitors in myeloproliferative neoplasms
Charles E. de Bock

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
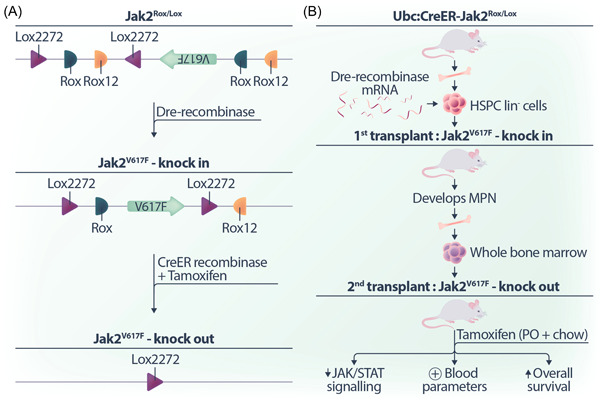 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMyeloproliferative Neoplasms: Diagnosis and Treatment · Kruppel-like factors research · Chronic Myeloid Leukemia Treatments
The discovery that the gain of function JAK2 ^ V617F ^ mutation is present in myeloproliferative neoplasms (MPNs) has led to numerous clinical trials assessing the efficacy of JAK inhibitors. Most notably, ruxolitinib, a combined JAK1/2 selective inhibitor, has gained approval in patients with myeolofibrosis (MF), and additional JAK2 inhibitors including fedratinib, pacritinib, and momelotinib also under evaluation for patients with MF. However, while these inhibitors demonstrate some clinical benefit, they do not adequately reduce the mutant clone fraction.1, 2 Consequently, a critical question for the field has been whether the lack of a durable response is attributed to either (i) the inability of current JAK inhibitors to completely block the pathway or (ii) the possibility that mutant clones are not entirely dependent on this activated pathway.
To address these two possibilities, a new study from the laboratory of Ross Levine, published in Cancer Discovery,3 developed an innovative mouse model of Jak2 ^ V617F ^ alone or in combination with Tet2 loss. The novel aspect of this mouse model lies in the ability to control the expression and genetic deletion of Jak2 ^ V617F ^ allele from mutant clones upon development of MPN. To do so, it utilizes two orthogonal site‐specific recombinases which exert precise control over the temporal expression and deletion of the Jak2 ^ V617F ^ allele.
The mouse model uses the well‐established Cre recombinase that recognises short nucleotide target sequences called Lox sites, in conjunction with the relatively new Dre recombinase which recognizes short nucleotide sequences called Rox sites. Importantly, the strategic arrangement and orientation of these sequences can lead to either flipping or deletion of the intervening DNA sequence. In this context, Dre recombinase is employed to initiate the expression of the Jak2 ^ V617F ^ allele. Subsequently, a modified CreER recombinase, translocated to the nucleus upon tamoxifen treatment, can delete the Jak2 ^ V617F ^ allele (Figure 1A). This intricate mouse model provided a powerful tool for comparing the durability of response between JAK inhibitors and the genetic loss of Jak2 ^ V617F ^ in the context of MPNs.
To understand the role of Jak2^V617F^ in the initiation and maintenance in MPNs in vivo, lineage‐negative hematopoietic stem and progenitor cells were harvested from donor Ubc:CreER‐Jak2 ^ Rox/Lox ^ mice. These cells were then electroporated with Dre recombinase‐encoding messenger RNA to induce Jak2^V617F^ expression and then transplanted into lethally irradiated recipient mice, that all went on to develop MPNs, confirming that Jak2^V617F^ is both sufficient and necessary for MPN development.
To investigate whether Jak2^V617F^ remained essential after the development of MPNs, the whole bone marrow of mice exhibiting MPN was injected into secondary recipients. At 12 weeks posttransplantation, the secondary recipient mice were treated with tamoxifen for Cre recombinase‐mediated deletion of the mutant Jak2 ^ V617F ^ allele. The authors found that the genetic deletion of Jak2 ^ V617F ^ not only reduced JAK/STAT signalling but also normalized key blood parameters and increased the overall survival of mice (Figure 1B).
Significantly, when extending the mouse model to include coincident loss of *Tet2^−/−^ *, the genetic loss of Jak2 ^ V617F ^ resolved hematological parameters to the same extent as observed in the single Jak2 ^ V617F ^ mutant setting, thereby providing evidence that the dependence on Jak2^V617F^ persists even in the presence of other high‐risk mutations.
Having established that Jak2^V617F^ remains essential, the authors proceeded to compare the transcriptional and phenotypic effects of Jak2^V617F^ genetic deletion with the impact of ruxolitinib treatment. Their findings showed that ruxolitinib was unable to reduce JAK/STAT or inflammatory signalling to the same degree as genetic deletion. Additionally, with ruxolitinib treatment, hematological parameters did not show comparable improvement, nor was there any reduction in mutant lineage‐Sca1+cKit+ cell fraction. Taken together, this mouse model supports the hypothesis that the elimination of mutant clones requires JAK inhibitors with enhanced potency.
One approach to enhancing the potency of JAK inhibitors has been the clinical development of type II JAK2 inhibitors that block JAK2 in the “inactive form.” In the current study, the authors show that the type II JAK2 inhibitor CHZ868 exhibits significantly greater efficacy in improving hematological parameters compared to ruxolitinb. In an independent study, another strategy has been to repurpose ruxolitinib and a related JAK inhibitor baricitinib as cereblon‐directed JAK proteolysis‐targeting chimeras (PROTACs). Notably, these PROTACs demonstrated efficacy within a pre‐B‐ALL mouse model.4
Nevertheless, even with improved potency, several questions remain. These include whether (i) the level of tyrosine kinase inhibition can ever fully phenocopy complete genetic deletion; (ii) resistance mechanisms, such as acquired point mutations, are inevitable; and (iii) potent inhibitors might drive the outgrowth of clones with other high‐risk mutations excluding JAK2^V617F^? Interestingly, in relation to the third question, a recent study found that patients with clones containing high‐risk mutations in genes such as TET2, DNMT3A, and ASXL1 but lacking JAK2^V617F^ did not impact prognosis.5
Overall, this new study provides a strong rationale for the ongoing development of more potent JAK2 inhibitors. It also nicely underscores the relevance of faithful animal models that accurately recapitulate hematological malignancies. Historically these mouse models have focussed on the expression of a single transgene or lesion like Jak2^V617F^. Nevertheless, as techniques like CRISPR/Cas9 become more efficient in introducing specific genetic mutations, the development of new models will continue to help address important clinical questions.
AUTHOR CONTRIBUTIONS
Charles E. de Bock is the sole author of this article.
CONFLICT OF INTEREST STATEMENT
The author declares no conflict of interest.
FUNDING
This research received no funding.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1How J , Garcia JS , Mullally A . Biology and therapeutic targeting of molecular mechanisms in MP Ns. Blood. 2023;141(16):1922‐1933.36534936 10.1182/blood.2022017416 PMC 10163317 · doi ↗ · pubmed ↗
- 2Kiladjian JJ , Vannucchi AM , Gerds AT , et al. Momelotinib in myelofibrosis patients with thrombocytopenia: post hoc analysis from three randomized phase 3 trials. Hema Sphere. 2023;7(11):e 963.37908862 10.1097/HS 9.0000000000000963 PMC 10615557 · doi ↗ · pubmed ↗
- 3Dunbar AJ , Bowman RL , Park YC , et al. Jak 2V 617F reversible activation shows its essential requirement in myeloproliferative neoplasms. Cancer Discovery. Published online March 4, 2024. 2024:OF 1‐OF 15. 10.1158/2159-8290.CD-22-0952 PMC 1106160638230747 · doi ↗ · pubmed ↗
- 4Chang Y , Min J , Jarusiewicz JA , et al. Degradation of Janus kinases in CRLF 2‐rearranged acute lymphoblastic leukemia. Blood. 2021;138(23):2313‐2326.34110416 10.1182/blood.2020006846 PMC 8662068 · doi ↗ · pubmed ↗
- 5Luque Paz D , Bader MS , Nienhold R , et al. Impact of clonal architecture on clinical course and prognosis in patients with myeloproliferative neoplasms. Hema Sphere. 2023;7(5):e 885.37153874 10.1097/HS 9.0000000000000885 PMC 10158927 · doi ↗ · pubmed ↗