Canine Schistosomiasis in the West Coast: Heterobilharzia americana in Two Natural Intermediate Hosts Found in the Colorado River, California
Anil Baniya, Connor J. Goldy, Jiranun Ardpairin, Perla Achi, Yu Wei Chang, Rose C. Adrianza, Apichat Vitta, Adler R. Dillman

TL;DR
This study reports the first discovery of Heterobilharzia americana, a parasitic worm, in two snail species along California's Colorado River, highlighting its public health and ecological implications.
Contribution
The study identifies the first occurrence of H. americana in California and confirms its presence in two new intermediate host snail species.
Findings
Heterobilharzia americana was found in Galba cubensis and Galba humilis snails along the Colorado River.
DNA sequencing and phylogenetic analysis confirmed the presence of H. americana in California.
The findings suggest a broader geographic distribution of the parasite than previously known.
Abstract
The emergence of infectious diseases presents a significant global health, economic, and security risk. Climate change can unexpectedly lead to the spread of pathogens, vectors, or hosts into new areas, contributing to the rise of infectious diseases. Surveillance plays a crucial role in monitoring disease trends and implementing control strategies. In this study, we document the first discovery of Heterobilharzia americana, a parasitic schistosome of mammals and its intermediate hosts Galba cubensis and Galba humilis along the banks of the Colorado River in California. We conducted multiple samplings of snails from various locations in the region with a previous history of canine schistosomiasis. Nucleotide sequencing of the multiple regions of the snails’ and parasites’ DNA revealed the coexistence of G. cubensis and G. humilis, both infected with H. americana. Phylogenetic analyses…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
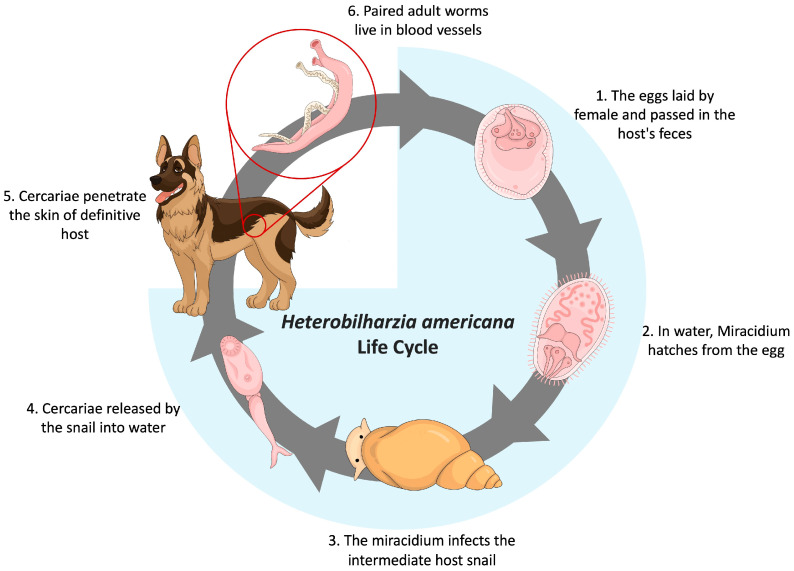 Figure 1
Figure 1 Figure 2
Figure 2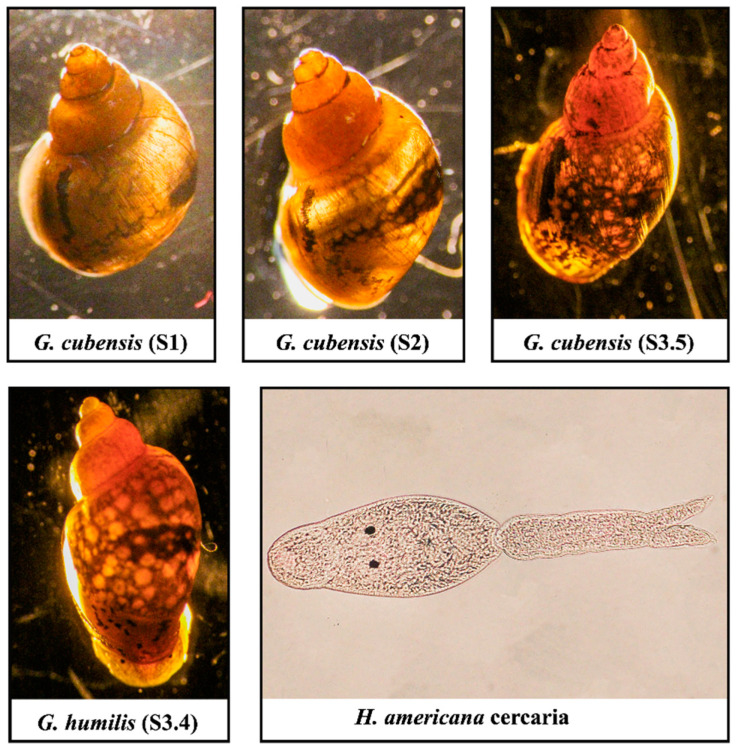 Figure 3
Figure 3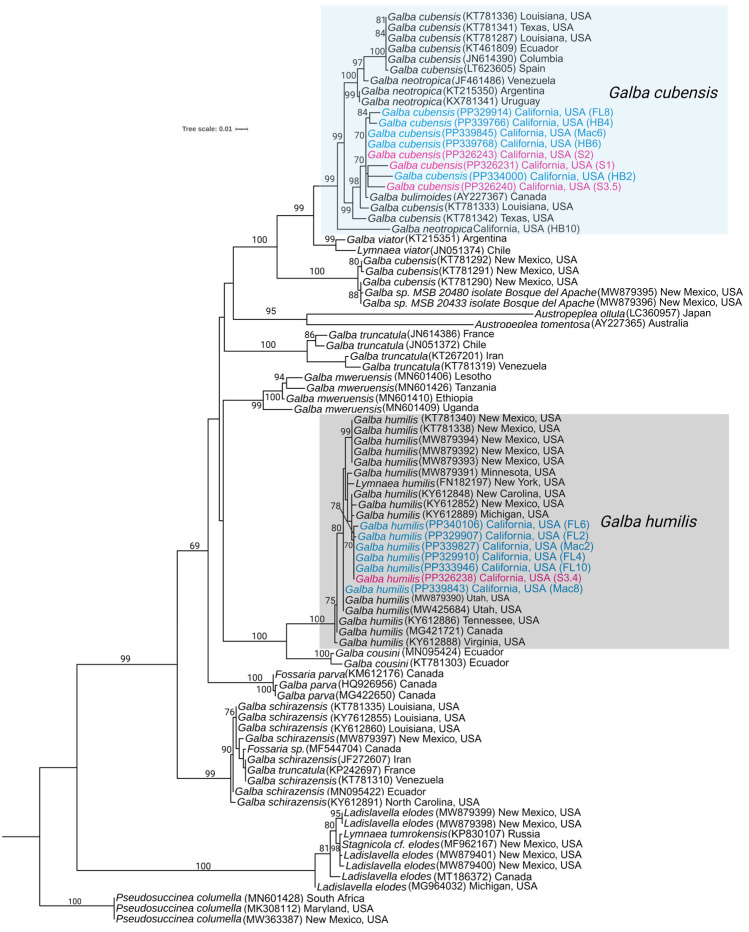 Figure 4
Figure 4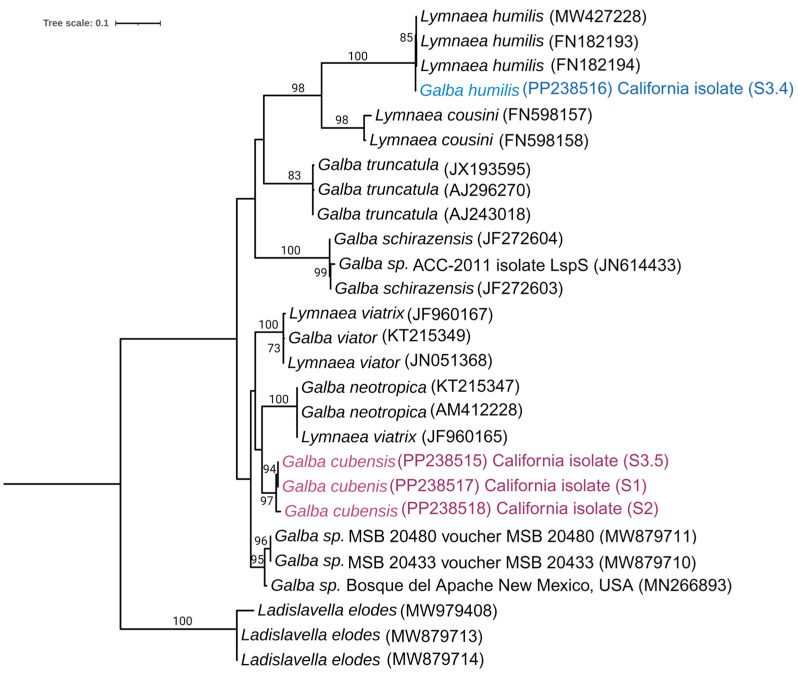 Figure 5
Figure 5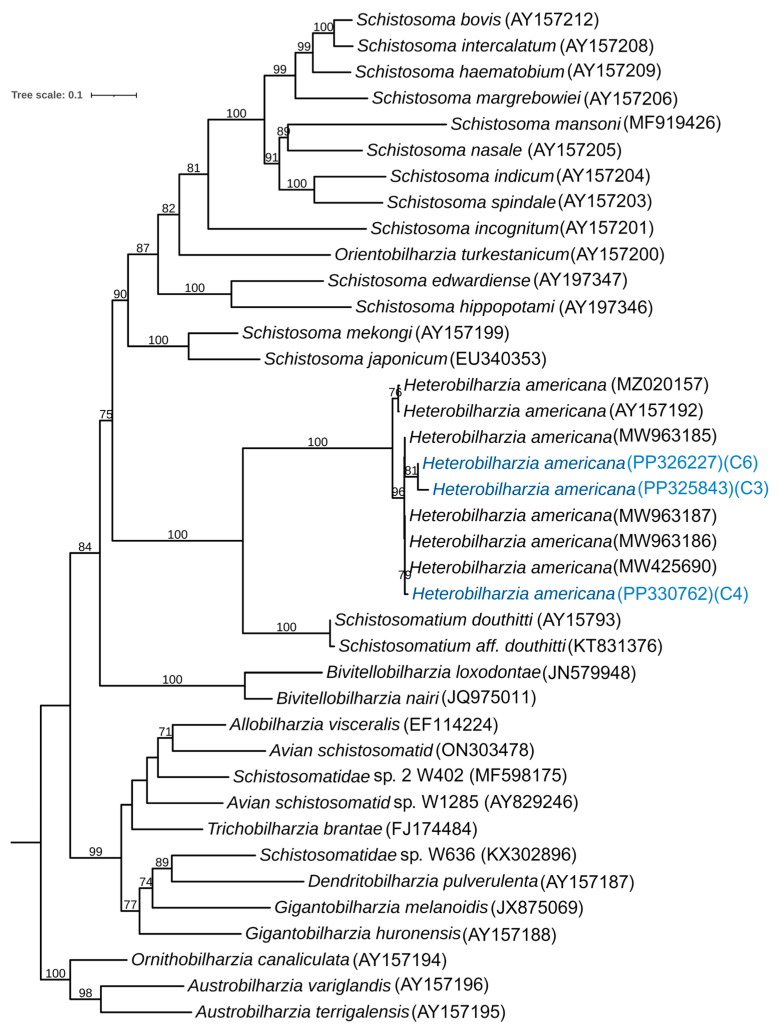 Figure 6
Figure 6 Figure 7
Figure 7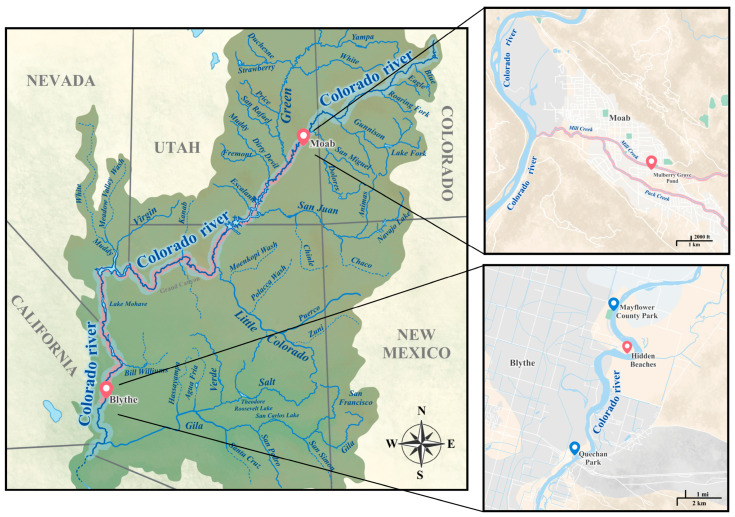 Figure 8
Figure 8- —National Institute of General Medical Sciences
- —Royal Golden Jubilee Ph.D. Program
- —National Research Council of Thailand (formerly the Thailand Research Fund)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsParasites and Host Interactions · Parasite Biology and Host Interactions · Zoonotic diseases and public health
1. Introduction
The rise of new and infectious diseases poses a substantial and increasing risk to global health, the worldwide economy, and international security [1]. Most emerging infectious diseases are zoonotic and originate from domesticated animals and wildlife [2,3]. Climate change resulting in vector and intermediate host range expansion may have widespread effects on the emergence of infectious diseases [4,5]. Examples of this include the outbreak of canine schistosomiasis in Utah [6] and the range expansions of various diseases, such as ticks and the meningeal worm Parelaphostrongylus tenuis [7,8,9]. Another potential source for the emergence of new diseases is the encounter of a particular pathogen or parasite with immune-compromised hosts. Examples include the recent infection of the nematode Ophidascaris robertsi in a woman’s brain and the horizontal transmission of cancerous tapeworm cells from Hymenolepus nana to a man’s lungs [10,11]. Disease and pathogen surveillance is a critical component of the health system because it provides timely information to monitor disease trends, evaluate health outcomes, and set parasite control and elimination strategies before they cause widespread disease. Here, we outline the discovery of a pathogenic schistosome infecting both wild and domestic mammals, along with its intermediate hosts, on the banks of the Colorado River in California.
Heterobilharzia americana is a schistosome trematode endemic to the Gulf Coast and South Atlantic region of North America, infecting raccoons, marsh rabbits, dogs, horses, nutria, bobcats, mountain lions, opossums, and other mammals [12,13]. Nevertheless, there is a growing number of reports documenting the incidence of parasite infection across various new states, including Indiana [14], Tennessee [15], Oklahoma [13,16], Arkansas [13], Kansas [17], and, most recently, Utah [6]. The life cycle of H. americana is that of a typical digenic trematode within the family Schistosomatidae (Figure 1). Mammals that wade or swim in freshwater areas, such as marshes, mudflats, ponds, and canals, are exposed to H. americana [18]. The intermediate host snails release the free-swimming cercariae into the water, which then penetrate the definitive host’s skin. Once inside the skin, the parasite transforms into a juvenile form called a schistosomula. The young schistosomes first migrate to the lungs, where they can cause hemorrhaging [19]. Afterward, they reach the liver and undergo development into both male and female. The male and female migrate to the mesenteric veins, mate, and lay spine-free eggs. Eggs make their way into venules, penetrating the intestinal wall, and are then released into the intestinal lumen. They are eventually excreted in feces. Upon reaching freshwater, the eggs hatch rapidly, releasing ciliated miracidia that swim to locate a suitable snail host. In infected snails, asexual development leads to the formation of both mother and daughter sporocysts and cercariae which then migrate to the hepatopancreas/digestive gland. It takes approximately 68 days post-infection to observe eggs in the feces of the definitive host [15,19].
The schistosome H. americana has been reported to infect a wide range of mammalian species in various regions of the United States. However, for a long time, the only snail species believed to be a natural intermediate host for H. americana was the small amphibious lymnaeid, Galba cubensis [20]. In 2021, a new species of snail host, Galba humilis, was recorded as testing positive for natural infection with this pathogen [6], and in 2023, another snail species, Galba schirazensis, was reported to be a compatible intermediate host of H. americana [21]. Similarly, Pseudosuccinea columella [22], a widely distributed aquatic lymnaeid, is experimentally susceptible [23], but it has not been observed with natural infection. In the United States, Galba cubensis has been recorded in Alabama, California, Florida, Georgia, Louisiana, Mississippi, North Carolina, New Mexico, South Carolina, and Texas [24]. The historical range of H. americana appears to match the occurrence of this snail in most of these states. Populations of G. cubensis have also been recorded from South America, Mexico, the West Indies, and Europe [25,26]. Another intermediate host, Galba humilis, also has a wide geographical distribution within the USA from Alaska to Florida and worldwide including Canada, Japan, and South America [6,20,27,28,29,30].
Dogs, when exposed to H. americana, manifest symptoms such as dermatitis (caused by penetrating cercariae), vomiting, coughing, fever, bloody diarrhea, anorexia, weight loss, lethargy, polyuria, and polydipsia, ultimately leading to collapse. Granulomatous reactions provoked by H. americana eggs are notable in organs including the lungs, liver, pancreas, and intestine. Common clinicopathologic observations in infected dogs include anemia, dehydration, hyperglobulinemia, hypoalbuminemia, hypercalcemia linked to elevated parathyroid hormone-related protein levels, and eosinophilia [15,31,32,33,34]. Similar symptoms are observed in H. americana-infected raccoons and horses [35,36]. In dogs, H. americana infections are treated with high-dose praziquantel or fenbendazole, with variable outcomes ranging from complete cure to treatment ineffectiveness [32]. The prolonged nature of the disease, poor treatment response, and diagnostic challenges often lead to euthanasia, and the clinical signs associated with schistosomiasis can mimic those of tumors and other diseases, requiring expensive and invasive procedures that may delay definitive diagnosis and appropriate therapy [31,34]. Veterinarians consistently emphasize that H. americana is frequently overlooked in diagnoses and is becoming a growing concern, particularly in dogs, horses, and other mammals [13,14,15,16,37,38].
The discovery of H. americana and its natural intermediate hosts in the Colorado River is noteworthy. Understanding the pathogen’s dissemination should prompt preemptive control strategies. This becomes particularly crucial in averting substantial damage to dog populations and other natural mammalian communities. In the subsequent section, we explore the discovery of H. americana within its natural hosts, Galba cubensis and Galba humilis, coexisting along the banks of the Colorado River in California. This discovery serves as compelling evidence of westward expansion of H. americana in the United States.
2. Materials and Methods
2.1. Collection and Screening of Snails
Snails were collected from the banks of the Colorado River multiple times. The first sampling of the snail population was conducted on 1 March 2023, among 2 sites (Mayflower County Park (33.67119, −114.53319) and Quechan Park (33.60725, −114.53104)). The second sampling of the snails took place on 4 April 2023, with samples collected from four different sites: First Leavy Park, Quechan Park, Mayflower County Park, and Hidden Beaches (33.6566, −114.51738). The third sampling was carried out on 16 July 2023, specifically from the 2 sites (Quechan Park, and Hidden Beaches) where many snails were found during the second sampling. The collection of snails involved using handled kitchen strainers, which were swept through aquatic vegetation near the shoreline, or snails outside of shorelines at or above the waterline were picked using hands. Snails were washed, isolated in individual wells of 24-well plates, and left overnight in the dark before screening to identify individual snails shedding cercariae. Snails were re-screened the next morning. Snails that did not shed cercariae were maintained in the laboratory and re-evaluated every day for two weeks. Snails releasing cercariae were photographed, and both the individual snails and their cercariae were subjected to molecular identification.
2.2. DNA Extraction for Molecular Identification of Snails and Cercariae
Snail populations from individual surveys were grouped based on shell morphology, and DNA extraction was performed on each morphotype for molecular identification. Total DNA was extracted from the foot tissue of the snails. Each snail was placed under the microscope, and a small piece of foot tissue was cut using a sharp scalpel. The tissue was then placed in 30 μL of extraction buffer in a 200 µL microcentrifuge polymerase chain reaction (PCR) polypropylene tube, mixed with 27 μL of 10 mM Tris, 1 mM EDTA pH 8.0 (Thermo Fisher Scientific, Fair Lawn, NJ, USA), 1.5 μL of 2% Triton X, and 1.5 μL of Proteinase K (20 mg/mL, New England Biolabs, Ipswich, MA, USA). The snail tissue was disrupted by freezing and thawing five times using liquid nitrogen, and then the tissue was incubated overnight at −20 °C. The following day, the frozen lysate was incubated at 56 °C for 1 h, followed by 95 °C for 10 min. DNA from cercariae was extracted using a similar technique as described above. Approximately 100 cercariae were placed in 20 μL of extraction buffer in a 200 µL microcentrifuge PCR tube, mixed with 18 μL of 10 mM Tris, 1 mM EDTA pH 8.0 (Thermo Fisher Scientific, Fair Lawn, NJ, USA), 1 μL of 2% Triton X, and 1 μL of Proteinase K (20 mg/mL, New England Biolabs, Ipswich, MA, USA). A similar protocol was followed to disrupt the cell and extract DNA as described for snail tissues above.
2.3. DNA Amplification and Sequencing
Amplification of multiple loci in snail and cercariae DNA was performed using different primer sets. We used 1 µL of the DNA extract as a template for a 25 µL PCR reaction, using 12.5 µL of Taq polymerase (Promega, Madison, WI, USA), 1 µL each of forward and reverse primer, and 9.5 µL of nuclease-free for all the PCR tests. Snail ITS1 was amplified using Lim1657 (forward) 5′-CTGCCCTTTGTACACACCG-3′, ITS1-RIXO (reverse) 5′-TGGCTGCGTTCTTCATCG-3′ [20]. Snail ITS2 was amplified using NEWS2 (forward) 5′-TGTGTCGATGAAGAACGCAG-3′ and ITS2-RIXO (reverse) 5′-TTCTATGCTTAAATTCAGGGG-3′ [39]. Thermocycling conditions for snail ITS1 (Lim1657, ITS1RIXO) were as follows: 94 °C for 6 min; 3 cycles for each annealing temperature, 60–56 °C then 20 cycles at 55 °C with denaturation at 94 °C for 30 s and extension at 72 °C for 2 min, and a final extension at 72 °C for 7 min. Similarly, the PCR conditions for ITS2 consisted of an initial denaturation step at 94 °C for 2 min. This was succeeded by 32 cycles, with intervals of 30 s at 94 °C, 30 s at 50–58 °C, and 30 s at 72 °C. The process concluded with a final extension step of 7 min at 72 °C. The COX 1 gene of the snails was amplified using LCOI490 (forward) 5′-GGTCAACAAATCATAAAGATATTGG-3′ and HCO2198 (reverse) 5′-TAAACTTCAGGGTGACCAAAAAATCA-3′ [40]. The PCR condition for the amplification of the COX I gene was denaturation at 94 °C for 2 min, 30 cycles of 30 s at 94 °C, 30 s at 52 °C, and 2 min at 72 °C, followed by a final 7 min extension at 72 °C.
For the parasite DNA, amplification of mitochondrial cytochrome oxidase subunit 1 (CO1) was conducted using Cox1_schist_5 (forward) 5′-TCTTTRGATCATAAGCG-3′, Cox1_schist_3 (reverse) 5′-TAATGCATMGGAAAAAAACA-3′ [41] and CO1F15 (forward) 5′-TTTNTYTCTTTRGATCATAAGC-3′, and CO1R15 (reverse) 5′-TGAGCWAYHACAAAYCAHGTATC-3′ [42]. The amplification of the CO1 gene used the same PCR conditions as described above for the amplification of the CO1 gene in the snails. Amplification of the large subunit region was performed using the primers C1 (forward) 5′-ACCCGCTGAATTTAAGCAT-3′ and D2 (reverse) 5′-TGGTCCGTGTTTCAAGAC-3′ [43].
PCR results were visualized on 1% agarose gel stained with 0.0003% ethidium bromide, along with a 1 kb plus DNA ladder (New England Biolabs, Beverly, MA, USA). Gel purification of the PCR products was performed using the QIAquick^®^ Gel Purification Kit (Qiagen, Germantown, MD, USA), following the manufacturer’s protocol. Subsequently, the purified PCR products were sent for Sanger sequencing using both the forward and reverse strands at the UCR Core Instrumentation Facility, according to the manufacturer’s protocol. The chromatogram sequence uncertainties in both the forward and reverse sequences for each locus underwent visual inspection and assembly through SeqManII software (DNASTAR Inc., Madison, WI, USA). The sequences generated as part of this work have been deposited in NCBI GenBank with their corresponding accession for snail and H. americana sequences (Supplementary Materials).
2.4. Identification and Phylogenetic Analysis
Identification of closely related species for each amplicon from snails and Heterobilharzia americana was conducted using the Basic Local Alignment Search Tool (BLAST) within the National Center for Biotechnology Information (NCBI) database. Based on the blast result, different closely related species of snails and H. americana species were used to construct the phylogeny. All sequences were aligned using MAFFT [44] with default parameters and the aligned sequences were visualized using Clustal W in MEGA 1; any alignment inconsistencies were corrected manually [45]. The IQ-Tree program [46] was used to choose the best model utilizing the ‘find best model’ function and to construct a maximum likelihood (ML) phylogenetic tree, incorporating ultrafast bootstrap branch support with 1000 replicates.
3. Results
3.1. Survey Results
From the first sampling, we collected 524 snails. Those snails were divided into four different groups based on the shell morphology (Figure 2). For all our surveys, the first and predominant category (comprising more than 90%) of snails were right-handed cone-shaped snails with very high shell spires, round and unobtrusive tentacles, and a foot that was yellowish to light olive. The second group of snails had spiral whorls, giving the shell a uniform wood-brown or fawn-colored appearance, with the last whorl swollen at the end. The third group of snails was composed of left-handed cone-shaped snails with a size like that of the snails in the first group but with longer tentacles. The fourth group of snails had larger shell sizes compared to all the snails collected during our survey, with a large, inflated body whorl and a short shell. During our second and third surveys, we collected 664 and 788 snails, respectively. Since the only recorded hosts of H. americana are cone-shaped snails, our focus was solely on this type of snail, and we exclusively gathered specimens of this kind.
For the second survey, we extracted DNA from four randomly selected snails at each of the three locations (Hidden Beach, First Leavy Park, and Mayflower County Park) in group A. For group B snails, we randomly selected four individuals. In group C, all four left-handed snails collected during the survey underwent DNA extraction. Additionally, three snails were selected from group D. The amplification and sequencing of the cytochrome oxidase subunit I (COX1) genes generated fragments ranging in size from 594 to 680 base pairs (bp). The Basic Local Alignment Search Tool (BLAST) analysis of the sequenced genes revealed that the majority of snails from group A were identified as two snail species, Galba humilis and Galba cubensis, with identity ranging from 98.76% to 100%. All the snails from group B were identified as Polygyra spp., with identities ranging from 96.74% to 99.83%. Snails from group C were identified as Physella acuta, with identities ranging from 98.23% to 99.56%. Similarly, snails from group D were identified as Succinea spp., with identities ranging from 99.50% to 99.83% (Supplementary Materials).
During our third survey, we focused solely on collecting cone-shaped snails (Galba spp.) from group A, with the majority obtained from Hidden Beach (33.6566, −114.51738). We collected 788 snails, and among them, four were shedding schistosome cercariae. These four snails underwent DNA extraction and sequencing (Figure 3 and Supplemental Videos). The sequencing of the COX I gene produced sequences ranging in size from 486 to 642 bp. BLAST analysis identified three snails as Galba cubensis, with identities ranging from 98.14% to 98.68%. Another fragment, measuring 655 bp in length from the fourth snail, was identified as Galba humilis with an identity of 99.52% in the database. We performed DNA extraction from four cercariae, amplifying the partial large subunit (28S) and cytochrome oxidase subunit I (COX1) gene. The 28S fragment lengths ranged from 897 bp to 902 bp, and the COX I gene fragment lengths ranged from 463 bp to 955 bp. BLAST results of the 28S sequences showed high similarity to the Heterobilharzia americana Utah strain, with a percent identity ranging from 99.56% to 99.89%. Similarly, BLAST results of the COX I gene for the California cercariae were highly similar to the H. americana Utah strain, with a percent identity ranging from 97.69% to 99.17%.
3.2. Phylogenetic Analysis of the Snail
For the phylogenetic analysis involving the cytochrome oxidase subunit I (COX1) gene of snails, we incorporated sequence data from both the second and third surveys. In the second survey, we included snails from three locations, selecting 4 snails from each site, resulting in a total of 12 snail samples. In the third survey, we specifically considered the sequence information from four snails that were shedding cercariae. Utilizing the BLAST results from NCBI for the sequenced snails, we identified 60 distinct snail isolates for comparison in constructing the phylogeny. In total, 86 sequences from different species were used for sequence alignment using MAFFT, and the IQ-Tree program determined K3Pu+F+I+G4 as the most fitting model for constructing a maximum likelihood (ML) phylogenetic tree. In the COX1 phylogeny, Pseudosuccinea columella from South Africa, Maryland, and New Mexico, with accession numbers MN601428, MK308112, and MW233387, respectively, were employed as an outgroup (Figure 4).
Based on the COX1 phylogeny, all the lymnaeid from group A were divided into two clades: Galba cubensis and Galba humilis. Species of snails in the Galba cubensis clade collected from California are strongly supported, as they clustered with representatives of this species from geographically distant regions within the United States, Canada, and South America. Similarly, Galba humilis species collected during our study group with strong statistical support, aligning with other snail species collected in the United States, such as Utah, New Mexico, and North Carolina. This robust support for the two clades suggests that these freshwater snails coexist in the same habitat along the banks of the Colorado River in California (Figure 4).
In another phylogenetic analysis of snails, we sequenced the partial Internal Transcribed Spacer 1 (ITS1) genes of four snails that were shedding cercariae. By utilizing the BLAST results from NCBI for these sequenced snails, we identified 23 distinct snail isolates for comparison in constructing the phylogeny. In total, 27 sequences from different species were used for sequence alignment using Mega 11, and the IQ-Tree program determined TPM2+G4 as the most fitting model for constructing a maximum likelihood (ML) phylogenetic tree based on the Bayesian Information Criterion (BIC). Different isolates of Ladislavella elodes, with accession numbers MW979408, MW879713, and MW879714 were employed as an outgroup (Figure 5).
A phylogenetic study of H. americana was conducted using the cytochrome oxidase subunit I (COX1) and the 28S region of the large subunit. For the phylogenetic tree based on COX1, we sequenced three isolates of cercariae and, based on the BLAST results in NCBI, identified 36 different isolates for the phylogenetic study. Based on the alignment of 39 sequences, the IQ-Tree program determined GTR+F+I+I+R4 as the best fitting model for constructing a maximum likelihood (ML) phylogenetic tree, based on the Bayesian Information Criterion (BIC) (Figure 6). The determination of the outgroup was conducted based on the previous analyses [6].
Heterobilharzia americana cercariae collected during our study clustered together and were placed next to H. americana collected in Utah. Furthermore, all the strains of H. americana were clustered together in the same clade next to Schistosomatium douthitii, suggesting close relationships between H. americana and S. douthitti. The results also revealed that isolates of H. americana from California are more similar to those from Utah and Texas than to an isolate from Louisiana (Figure 6).
Sequencing of the 28S region of H. americana and BLAST analysis on GeneBank resulted in the identification of 28 different species of Schistosomatidae. A total of 31 sequences were aligned for the study of their interrelationship. The IQ-Tree program determined TVM+F+G4 as the most fitting model for constructing a maximum likelihood (ML) phylogenetic tree based on the Bayesian Information Criterion (BIC). The outgroup was determined as Chimaerohemecus trondheimensis based on a previous study [41]. All the isolates of the H. americana were placed together in the same clade next to Schistosomatium douthitii, confirming their close relationship and validating the maximum likelihood tree based on the COX1 gene (Figure 7).
4. Discussion
In 2023, the Los Angeles County of Public Health issued an advisory confirming 11 cases of canine schistosomiasis in dogs from three Southern California counties: Los Angeles, Orange, and Riverside, between 2018 and 2023. The parasite had not previously been reported as endemic to Southern California. Travel history of the infected dogs indicated that before their diagnosis, all 11 dogs had been in the Colorado River [37]. This finding led us to conduct our survey [12,14,17,18,22,31,32,35]. Based on our study, we were able to recover H. americana cercariae from two naturally occurring snails, Galba cubensis and Galba humilis, coexisting together along the banks of the Colorado River. This finding helps to support the notion that the infected dogs likely contracted H. americana from the Colorado River. This finding is also the most westerly report of this endemic North American schistosome, suggesting that this parasite has a wider range than previously speculated and is expanding its distribution beyond what was previously reported [6].
Initially, G. cubensis was exclusively considered to be the natural host of H. americana [18,20]. Another species, G. humilis from Michigan, showed marginal susceptibility upon experimental exposure to H. americana [12]. However, in 2021, G. humilis was discovered to be naturally infected with H. americana in a human-made pond in Moab, Utah. This discovery was recorded at Mulberry Grove Pond, which receives water from Mill Creek, one of the tributaries of the Colorado River. This pond is approximately 280 feet from Mill Creek, and the shore of the Colorado River is approximately 2.17 miles away from the pond [6]. The newly identified natural host had a 77.5% positive infection rate following experimental exposure of H. americana, suggesting increased compatibility with G. humilis, which is widely distributed across North America [6]. Recently, another snail species Galba schirazensis was newly reported to be a compatible host of H. americana [21]. Our study provides the first report of the coexistence of two snail species, G. cubensis and H. humilis, in the same location during multiple samplings, both naturally infected with H. americana. This validates a wider distribution of H. americana than previously speculated (Figure 6). Galba is a genus of small-shelled freshwater snails that primarily originated in the Americas and subsequently invaded Asia, Africa, and Europe [47,48,49,50,51]. Their capacity to endure periods of drought and reproduce through self-fertilization enables them to spread across extensive distances and establish new populations from individual snails [47,52]. Their high invasiveness may be one reason for the broader expansion of the parasite associated with these snails [53]. Our phylogenetic analysis, which revealed other Galba species closely related to G. cubensis and G. humilis, combined with the identification of diverse snail species in our survey, indicates the potential for other snail species to be potential hosts of H. americana. Our finding of H. americana on the shore of the Colorado River in the southernmost part of the United States leads us to hypothesize that the spread of this parasite is throughout the Colorado River and among its tributaries. This is because the Colorado River and its watershed area harbor a diversity of mammals that can serve as hosts for this parasite, contributing to the maintenance of H. americana populations [54]. In-depth studies are required to investigate the extent of parasite infestation along the banks of the Colorado River and its tributaries in the United States and Mexico to gain a clear understanding of the parasite’s infection (Figure 8).
Our primary sampling site is located on the banks of the Colorado River in a popular spot for recreation. The area is also abundant in vegetation, attracting various mammals such as raccoons, bobcats, and rabbits. The identification of H. americana from this location suggests that these naturally occurring mammals may be infected with this parasite, contributing to its population maintenance. The extent of infection in the natural mammal population in this area remains unknown. Further investigation is needed to understand the impact of this parasite on our biodiversity, as it poses a potentially severe threat. A survey in Texas emphasized the severity of H. americana infection among raccoons, indicating a prevalence of up to 47% in raccoons of all ages and up to 85% in older raccoons [55]. This high prevalence was suspected to be the cause of H. americana infection in horses in the same locality [36].
The study of H. americana biology presents a unique challenge due to limited information about the identification and distribution of the intermediate host, coupled with scant details about the schistosome itself. Schistosoma, a genus of blood flukes, is responsible for causing schistosomiasis—a widespread and challenging disease affecting both humans and animals. Approximately 200 million people are infected with schistosomes, and an estimated 500–600 million individuals in various tropical and subtropical countries are at risk of contracting the disease [56]. Furthermore, the penetration of cercariae from animal schistosomes into human skin can result in irritating outbreaks of cercarial dermatitis, commonly known as swimmer’s itch [57]. There are at least 13 identified genera and more than 100 species of schistosomes to date. The larval stage of these schistosomes infects intermediate host snails and parasitizes a variety of organisms, including birds and mammals [58,59].
The schistosome identified in our survey, H. americana, is primarily a parasite of domestic and wild mammals. The exact host range of this parasite is not known; further studies are required to validate the susceptibility range. A study conducted in 1967, where H. americana was experimentally introduced into humans, revealed that the parasite caused vesicles at the infection site, and the rash subsided 5–8 days postinfection. In infected rhesus monkeys, no eggs were recovered from the feces; however, young schistosomules were retrieved from the liver 21 days after exposure, and pre-adults were observed in the liver of a second monkey 45 days after exposure [22]. Another study revealed a variable range of susceptibility among primates [60]. There is much more to explore regarding the pathogenicity and host range of this parasite, highlighting the need for further experimentation.
5. Conclusions
In our study, we successfully confirmed the presence of Heterobilharzia americana for the first time along the shores of the Colorado River, infecting two species of snails, Galba humilis and Galba cubensis. This significant finding marks the westernmost record of this endemic North American schistosome in the United States. The identification of the parasite in an area with a documented history of canine schistosomiasis emphasizes the persistence and potential expansion of this parasitic threat. This parasite’s presence in the area suggests a broader distribution than previously reported, highlighting the need for a better understanding of its geographical range. Such information is critical given the potential impact on natural biodiversity, domestic pets, and human health. The threat posed by H. americana, with its complex life, warrants careful consideration and proactive measures. Future research should focus on understanding the severity of the infestation, the dynamics of the parasite’s life cycle, and its interactions with various hosts. An in-depth understanding of these aspects is crucial for formulating effective strategies to manage and control the spread of H. americana. To conclude, our study sheds light on the expanding range of H. americana. These findings underscore the need for continued research, surveillance, and strategic planning to mitigate the potential ecological, veterinary, and public health impact of this parasite.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Allen T. Murray K.A. Zambrana-Torrelio C. Morse S.S. Rondinini C. Di Marco M. Breit N. Olival K.J. Daszak P. Global Hotspots and Correlates of Emerging Zoonotic Diseases Nat. Commun.20178112410.1038/s 41467-017-00923-829066781 PMC 5654761 · doi ↗ · pubmed ↗
- 2Zhou P. Yang X.L. Wang X.G. Hu B. Zhang L. Zhang W. Si H.R. Zhu Y. Li B. Huang C.L. A Pneumonia Outbreak Associated with a New Coronavirus of Probable Bat Origin Nature 202057927027310.1038/s 41586-020-2012-732015507 PMC 7095418 · doi ↗ · pubmed ↗
- 3Gire S.K. Goba A. Andersen K.G. Sealfon R.S.G. Park D.J. Kanneh L. Jalloh S. Momoh M. Fullah M. Dudas G. Genomic Surveillance Elucidates Ebola Virus Origin and Transmission during the 2014 Outbreak Science 20143451369137210.1126/science.125965725214632 PMC 4431643 · doi ↗ · pubmed ↗
- 4Hosie M.J. Jasim S. The Case for Adopting a Combined Comparative Medicine and One Health Approach to Tackle Emerging Diseases Vet. Rec.20201872410.1136/vr.m 268633638551 PMC 7456687 · doi ↗ · pubmed ↗
- 5Laing G. Vigilato M.A.N. Cleaveland S. Thumbi S.M. Blumberg L. Salahuddin N. Abdela-Ridder B. Harrison W. One Health for Neglected Tropical Diseases R. Soc. Trop. Med. Hyg.202111518218410.1093/trstmh/traa 117PMC 784210233169163 · doi ↗ · pubmed ↗
- 6Loker E.S. Dolginow S.Z. Pape S. Topper C.D. Alda P. Pointier J.P. Ebbs E.T. Sanchez M.C. Verocai G.G. De Jong R.J. An Outbreak of Canine Schistosomiasis in Utah: Acquisition of a New Snail Host (Galba humilis) by Heterobilharzia americana, a Pathogenic Parasite on the Move One Health 20211310028010.1016/j.onehlt.2021.10028034258371 PMC 8254006 · doi ↗ · pubmed ↗
- 7Brinkerhoff R.J. Folsom-O’Keefe C.M. Tsao K. Diuk-Wasser M.A. Do Birds Affect Lyme Disease Risk? Range Expansion of the Vector-Borne Pathogen Borrelia burgdorferi Front. Ecol. Environ.2011910311010.1890/090062 · doi ↗
- 8Wasel S.M. Samuel W.M. Crichton V. Distribution and Ecology of Meningeal Worm, Parelaphostrongylus tenuis (Nematoda), in Northcentral North America J. Wildl. Dis.20033933834610.7589/0090-3558-39.2.33812910761 · doi ↗ · pubmed ↗