Detection and characterization of the Pichia manshurica biofilm on the traditionally produced homemade apple vinegar
Zeynep İŞLEK KÖKLÜ, Pınar AKKUŞ SÜT, Esra ESKİHORAN ÜÇÜNCÜOĞLU, Sadık KALAYCI, Fikrettin ŞAHİN

TL;DR
This study examines how Pichia manshurica biofilms affect homemade apple vinegar, showing they can spoil the product by forming harmful structures.
Contribution
The study identifies and characterizes P. manshurica biofilms in homemade vinegar for the first time.
Findings
P. manshurica was found to form biofilms on the cellulosic film in vinegar.
Contaminated biofilms showed higher polysaccharide content and altered structure.
The presence of P. manshurica negatively impacts vinegar quality during fermentation.
Abstract
Pichia yeasts are capable of forming biofilms during vinegar production and causing spoilage in various beverages. In addition, there exists a significant likelihood of encountering yeast contamination which can prevent vinegar production. The present study investigates the detection and characterization of the Pichia manshurica (P. manshurica) biofilm on traditionally produced homemade apple vinegar. The unique characteristics of vinegar were analyzed with a focus on the constituent, known as the “mother of vinegar”, whose composition is comprised of cellulosic biofilm and acetic acid bacteria, including Gluconobacter oxydans (G. oxydans) Briefly, P. manshurica was isolated from apple vinegar and characterized in terms of the effect of biofilm formation on the surface of the cellulosic film on vinegar production. Microbial identification of vinegar with/without contamination by P.…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
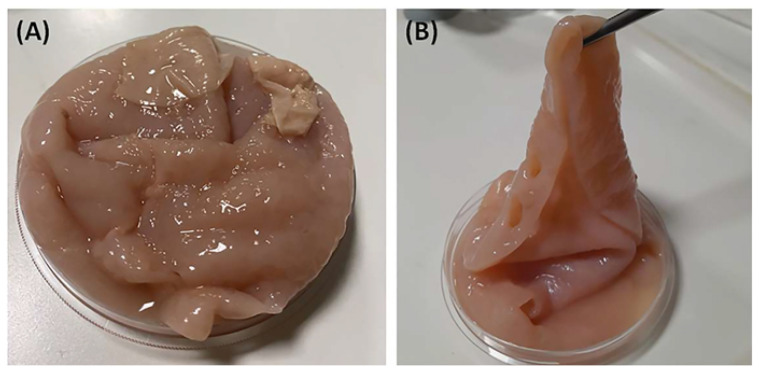 Figure 1
Figure 1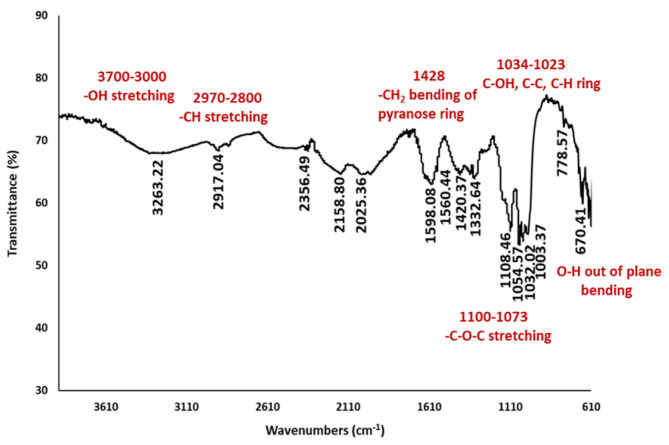 Figure 2
Figure 2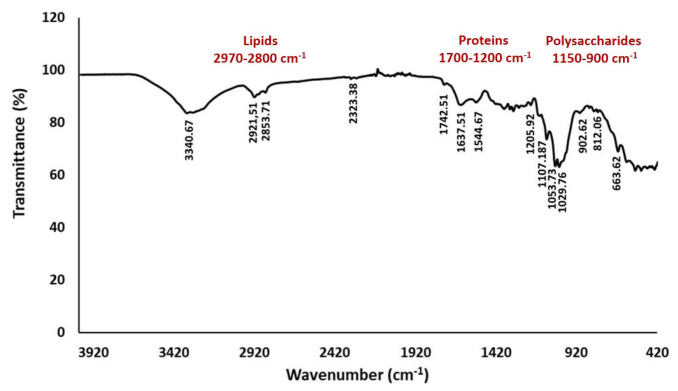 Figure 3
Figure 3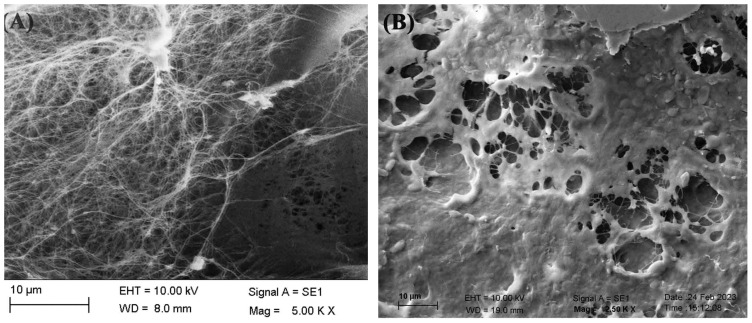 Figure 4
Figure 4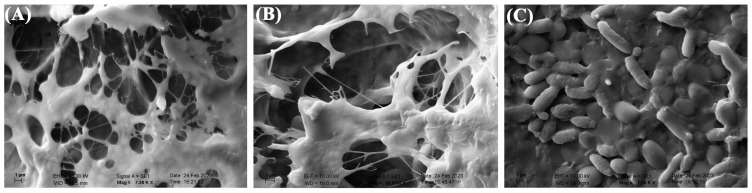 Figure 5
Figure 5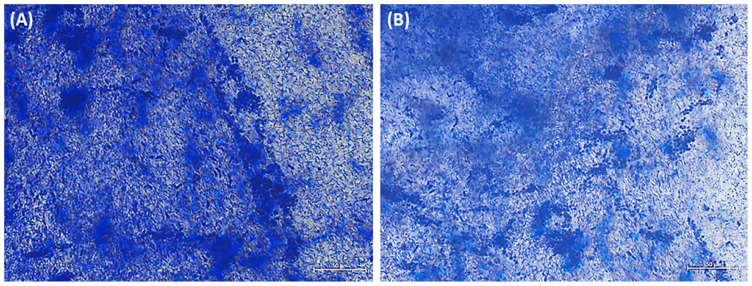 Figure 6
Figure 6- —Department of Genetics and Bioengineering, Yeditepe University
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsFermentation and Sensory Analysis · Botulinum Toxin and Related Neurological Disorders · Microbial Metabolism and Applications
1. Introduction
Vinegar is a popular condiment that is widely used in cooking, food preservation, and as a natural cleaning agent. It can be produced from a wide variety of fermentable carbohydrate sources, such as fruits, honey, grains, and starchy vegetables. It is commonly used as a food preservative, and according to the Food and Drug Administration (FDA), it contains 4% acetic acid that is produced through the fermentation of sugary or alcoholic materials [1,2]. Vinegar comprises diverse quantities of stable fruit acids, salts, pigments, and volatile components, such as esters and phenolics, which give it its characteristic aroma and flavor [3]. Apple cider vinegar is widely known for its therapeutic properties and health advantages. It is rich in trace minerals and vitamins, such as vitamins A, C, E, and B, beta carotene, and bioflavonoids, which play a crucial role in cellular function [4]. Vinegar possesses various properties, including antiglycemic, antihypertensive, antimicrobial, antioxidant, and antitumor activities. Additionally, vinegar serves as a fundamental component in Ayurvedic preparations [4].
In Türkiye, there are various types of local vinegar that are specific to different regions. Apple vinegar, which has a history dating back more than 3500 years, is a typical and characteristic product in Türkiye. Traditionally, vinegar has been produced through a slow, natural, and spontaneous fermentation process involving acetic acid bacteria [5]. Besides, fruit juices have conventionally been used as a source for vinegar production in both household and industrial settings, in a two-stage procedure. Initially, fruit sugars undergo fermentation by yeasts, predominantly Saccharomyces ellipsoideus (S. ellipsoideus), and S. cerevisiae, resulting in the production of alcohol. Subsequently, acetic acid bacteria facilitate the oxidation of ethanol and any remaining or supplemented sugars into acetic acid [4,6]. These gram-negative or gram-variable organisms are ellipsoidal to rod-shaped, 0.6–0.8 μm × 1.0–0.4 μm in size [4]. It is possible to find pleomorphic forms that are spherical, elongated, distended, curved, or filamentous. They can convert substrates like glucose, ethanol, lactate, and glycerol into acetic acid. The classification of bacteria into two primary genera, namely Acetobacter and Gluconobacter, is based on their capacity to overoxidize acetate or lactate, as well as the arrangement of their flagella. Additionally, these are known to produce a film or pellicle composed of cellulose. The acetic acid bacteria and yeasts in the fermentation system become intertwined with the cellulosic pellicle to create the “mother of vinegar” [4]. Pichia manshurica (P. manshurica) is a yeast species that has been reported in various fermented foods, such as table olives, cocoa beans, wine, and vinegar. The documented spoilage of wine is attributed to the release of volatile phenols and cadaverine. P. manshurica exhibits a high degree of adaptation to the wine environment, although it is vulnerable to cleaning agents. Aerobic yeast species, including Pichia, which are deemed undesirable, have the capability to develop a film layer at the interface of air and wine upon exposure to air. This microorganism is capable of colonizing filter surfaces, thereby leading to contamination in wineries, as has been documented in previous studies [7,8]. P. manshurica poses a significant challenge due to its resistance to organic acids, resulting in the production of undesirable sensory attributes such as odors, tastes, and products. During the process of vinegar production utilizing spontaneous fermentation, there exists a heightened possibility of contamination with the development of biofilm and malodor on the surface of the vinegar. The yeasts that participate in the alcohol fermentation stage may lead to complications by catalyzing the oxidation of alcohol to carbon dioxide and water, as has been previously noted [7]. Therefore, the utilization of yeast strains such as P. manshurica in the process of vinegar manufacturing may give rise to complications and is not recommended.
Vinegar production, on the other hand, involves spontaneous fermentation and traditional techniques that are influenced by factors such as water, soil, and climate. These factors determine the quality of the vinegar, and the presence of P. manshurica could impact the flavor and quality of the final product [9–12]. In the previous study, yeasts, including Saccharomyces, Pichia, and Candida (C.), are categorized into two types based on their function: alcohol-producing and aroma-producing, and are known genera involved in vinegar fermentation. S. cerevisiae is the most common alcohol-producing yeast species and dominates the alcoholic fermentation process. Besides, aroma-producing yeasts are less abundant but significantly affect the quality and flavor of alcoholic beverages and vinegar [9]. Therefore, it is important to improve information about the characterization of yeast biofilms such as P. manshurica, and to develop methods for their detection in vinegar production to prevent spoilage and ensure the production of high-quality vinegar traditionally produced by cellulose.
In this study, we aimed to explore the microbial detection and characterization of cellulose produced by Gluconobacter oxydans (G. oxydans), and P. manshurica-contaminated cellulosic biofilms in vinegar production, using MALDI-TOF mass spectrometry (MALDI-TOF MS), Fourier transform infrared spectroscopy (FT-IR) analysis, scanning electron microscopy (SEM), and crystal violet staining. Therefore, besides highlighting the detection of P. manshurica contamination in the cellulosic biofilms, it sheds light on the comparative characterization of bacterial cellulose used in vinegar production with the biofilm formed as a result of contamination.
2. Materials and methods
2.1. Materials
The sweet apples were collected from Edremit in Türkiye. Tryptic soy agar and malt extract agar were purchased from Sigma-Aldrich, Germany. HCCA Matrix was obtained from Bruker, United States. Crystal violet stain was purchased from Fluka, US.
2.2. Production of apple vinegar
To prepare the vinegar, 1.75 kg of sweet apples were separated from the stems and kept with 2.5% honey, chickpeas, and drinking water in a 10-L glass jar. The mixture was stirred for 2 weeks using a wooden spoon until the apples sank to the bottom. The jar was then covered with cheesecloth and stored at 25 °C and in a dark environment for 3 to 6 months to be fermented. Over time, it was observed that the water remained on top, while the apples sank to the bottom. As fermentation progressed, a membrane-like cellulose biofilm began to form on the apples, gradually thickening. Once fermentation was complete, the mixture underwent filtration, resulting in the separation of the pulp and vinegar constituents.
2.3. Collection of the vinegar samples
Homemade apple vinegar was traditionally produced without any pasteurization process or additives (Figures 1A and 1B). Each sample was collected in bottles and stored in laboratory conditions at 4 °C for 24 h.
2.4. Microbial identification of the apple vinegar
2.4.1. Isolation of microorganisms from the apple vinegar
The microbiological properties of the vinegar were determined using MALDI-TOF MS. Initially, the samples of the apple vinegar were enriched with alkaline peptone water. Subsequently, serial dilutions to 10^−1^ and 10^−6^ were prepared, and the diluted samples were plated on Tryptic Soy Agar and Malt Extract Agar. These plates were then incubated at 30 °C for 48, and 72 h. Suspicious microorganism colonies were selected from each agar and ultimately purified by reseeding onto fresh Tryptic Soy Agar and Malt Extract Agar plates, followed by additional incubation at 30 °C for 48 and 72 h. The purified colonies were then identified using MALDI-TOF MS (Bruker Daltonik GmbH, Bremen, Germany).
2.4.2. Identification of microorganisms within the apple vinegar using MALDI-TOF MS
Following 48 and 72 h of incubation, the Petri dishes were examined and a single colony structure was identified by matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (MALDI-TOF MS) (Bruker Microflex LT, Bruker Daltonik GmbH, Bremen, Germany). Briefly, a single colony from each Petri dish was transferred onto a MALDI-TOF MS target plate, covered with 1 μL of α-cyano-4 hydroxycinnamic acid (i.e. matrix solution) dissolved with 50% acetonitrile-2.5% trifluoroacetic acid (HCCA matrix), and left to dry at room temperature before protein analysis by MS. Subsequently, it was covered with the target, and analyzed by the mass spectra represented in term of the mass change ratios (m/z) of each microorganism. According to the library matching, the MS spectrum of unknown microbial isolates was compared with the MS spectra of known microbial isolates contained in the MALDI-TOF MS database. In order to detect the species level identification of microbes, a typical mass range m/z of 2–20 kDa was used by Flex Control software, in which mainly ribosomal proteins were represented together with housekeeping proteins.
2.5. Characterization studies of the biofilm
2.5.1. FT-IR analysis
Dried biofilm samples were analyzed by FT-IR spectrometer (Thermo Scientific, NICOLET iS50 FT-IR Spectrometer, USA) within the spectral region of 4000–400 cm^−1^. Data collection and processing were performed with OMNIC software, which was provided along with the instrument.
2.5.2. Morphological analysis by SEM
SEM (Zeiss EVO40, Germany, USA) was used to visualize the surface structure of the biofilm samples at an accelerated voltage of 10 kV. Briefly, the dried samples were placed onto a metallic stub and subjected to gold palladium sputtering (EM ACE200, Leica) for 45 s. The prepared samples were then examined under SEM (Zeiss EVO40). The experiments were repeated with three independent studies.
2.6. Crystal violet staining
Crystal violet staining solution (0.5%) was prepared in accordance with the previously described procedure [13]. Briefly, 0.5 grams of crystal violet powder was dissolved in 20 mL of methanol and the ultimate volume was adjusted to 100 mL with distilled water. Afterwards, small pieces were cut from the biofilm structure with the help of a scalpel. Biofilm pieces were treated with a crystal violet dye solution for 10 min. After staining, the biofilm pieces were washed 3 times for 5 min to remove excess dye. The stained biofilm pieces were taken on a glass slide and visualized with bright field imaging using an inverted microscope (ZEISS Axio Vert.A1).
3. Results
3.1. Microbial identification using MALDI-TOF MS
In accordance with the library database of MALDI-TOF MS, the MS spectrum of unknown microbial isolates was identified as G. oxydans ssp*. B544 UFL*, G. oxydans ssp*. B540 UFL*, and P. manshurica with a ratio of 2.01, 1.94, and 1.70, respectively.
3.2. FT-IR analysis of the cellulosic biofilm samples
Cellulosic biofilm produced by G. oxydans was analyzed by FT-IR to identify the chemical structure, as shown in Figure 2. Accordingly, cellulose-specific characteristic peaks in the range of 4000–400 cm^−1^ were observed in the FTIR spectrum of bacterial cellulose. The cellulosic sample indicated the characteristic peaks at 3263 cm^−1^ for O-H stretching vibration, at 2917 cm^−1^ for C-H stretching vibration, at 1420 cm^−1^ for C-H bending vibration, at 1032 to 1054 cm^−1^ for C-O-C and C-O-H stretching vibration of the sugar, and at 670 cm^−1^ for the OH out-of-phase bending vibrations (Figure 2).
3.3. FT-IR analysis of the P. manshurica-contaminated cellulosic biofilm
The P. manshurica-contaminated cellulosic biofilm structure was examined by FT-IR spectroscopy within the spectral region range of 4000–400 cm^−1^, in which the presence of chemical groups belonging to important components such as proteins, lipids, and polysaccharides was investigated (Figure 3). The peaks in the range of 1150–900 cm^−1^ indicated high polysaccharide content, in which the band region was mainly formed as a result of stretching vibration of C–C and C–O bonds and deformation of C–O–H and C–O–C bonds (Figure 3). The stretching vibration of the lipids was observed between 2921.51 and 2853.71 cm^−1^ (Figure 3).
3.4. Morphological analysis of the biofilms
The SEM images of cellulosic biofilm samples indicated a three-dimensional porous network structure consisting of randomly arranged ribbon-shaped ultrafine fibers (Figure 4A). The filaments and reticulated structures of contaminated cellulosic biofilms from G. oxydans and P. manshurica differ morphologically (Figures 4 and 5). Accordingly, bacterial cellulose was observed to lose its reticulated structure and nanofiber network, and was replaced by the structure of the P. manshurica biofilm (Figures 4B, 5A, and 5B). As a result of the convergence by thick layers, nanofibers were unable to be observed, indicating that the topology of the pore network was highly interconnected. Furthermore, two different microbial populations were visualized under the SEM images, in which the spherical or ovoid shape of micrometers in size, occurring singly, in pairs, in chains, or in groups, corresponds to P. manshurica yeast populations, whereas G. oxydans was detected with ellipsoidal to rod-shaped bacterial properties (Figure 5C).
3.5. Crystal violet staining
Cellulosic biofilms contaminated with P. manshurica were observed under brightfield microscopy, revealing the development of a layer of positive cells that exhibited crystal violet staining on the surface of the cellulosic biofilm (Figure 6). In parallel to SEM images, there was a development of filamentous G. oxydans bacterial and P. manshurica yeast colonization, resulting in the formation of a complex filamentous structure that was abundantly dispersed without showing any selective binding between the different surfaces of the biofilm (Figures 6A and 6B).
4. Discussion
Cellulose, which is used extensively in the paper and textile industry, can be obtained from plant sources (containing 60–90% cellulose), as well as produced by microorganisms such as bacteria, fungi, and algae. The high purity, mechanical properties, and high water-holding capacity of bacterially produced cellulose are advantageous for its use in different areas. Since the reporting of Acetobacter xylinum as a producer of cellulose, many cellulose-producing bacterial strains have been described previously [14,15], including Azotobacter, Gluconacetobacter, Pseudomonas, and Rhodobacter [16]. In the present study, G. oxydans, which is known as one of the cellulose-producing microorganisms, was determined by MALDI-TOF MS. Previously, simple sugars in the apple such as glucose, fructose, and sucrose were found be as important substrates that contribute to traditionally produced apple vinegar bacterial cellulose production, owing to its high sugar content, thus enhancing a favorable environment for cellulosic “mother of vinegar” production [15]. Together with the sugar source of the apples, Gluconobacter spp. has taken advantage of cellulose production [4]. In the process of vinegar production, there is a high risk of specific yeast contamination, including Pichia spp., involved in the alcohol fermentation, resulting from the oxidation of alcohol to CO_2_ and H_2_O [7]. Pichia species can lead to biofilm formation and excessive foul odor on the surface of vinegar production [8]. For this reason, it is necessary to determine the microorganism in the contamination, characterize the biofilm formation from Pichia, and perform its instrumental and morphological analysis, including MALDI-TOF MS, FT-IR, SEM, and crystal violet staining.
4.1. The chemical structural analysis of cellulosic biofilms by FT-IR
In the present study, the examination of microbial biofilm structures by FT-IR spectroscopy was carried out in the spectral region range of 4000–400 cm^−1^, which indicates the presence of chemical groups belonging to important components such as proteins, lipids, and polysaccharides. Previously, FT-IR biofilm analysis demonstrated that lipids, proteins, and polysaccharides were absorbed in the specific spectral ranges, including 3000–2800 cm^−1^, 1705–1600 cm^−1^, and 1200–950 cm^−1^, respectively [17,18]. Furthermore, in another study, the peaks of protein content were detected as amide I at 1600–1700 cm^−1^, amide II at 1500–1600 cm^−1^, and amide III at 1200–1350 cm^−1^ [17]. In addition, it has been reported that protein units exhibit IR band properties through C=O stretching at the amide I region, N–H bending and C–N stretching at the amide II region, and C–N bending and N–H stretching at the amide III region [17,19,20]. On the other hand, the peaks in the range of 1150–900 cm^−1^ indicated high polysaccharide content, in which the band region is mainly formed as a result of stretching vibration of C–C and C–O bonds and deformation of C–O–H and C–O–C bonds [21,22]. The peaks of symmetrical and asymmetrical C-H vibrations were observed between 2800 and 2970 cm^−1^, indicating the presence of lipid molecules in EPS due to the characteristic of lipids condensed in bacterial cell walls and membranes [23,24]. Similarly, in our biofilm sample, the stretching vibration of the lipids was observed between 2921.51 and 2853.71 cm^−1^ (Figure 3).
4.2. The morphological analysis of cellulosic biofilms by SEM
The SEM images of cellulosic biofilm samples were visualized as three-dimensional porous network structures and randomly arranged ribbon-shaped ultrafine fibers (Figure 4). Saxena et al. reported that cellulose synthesis was characterized by crystallization unidirectional growth, in which glucose molecules were bounded β (1–4) glucosidic bonds linearly [25,26]. Oriented microfibrils were formed by the union of glucosidic chains via intramolecular hydrogen bonds. The diameter of thin filaments was arranged between 34.34 and 39.16 nm, which is smaller than plant cellulosic fibrils [26,27]. The results were consistent with the previously reported cellulose structure [9,26,28]. The filaments and reticulated structures of contaminated cellulosic biofilms from G. oxydans and P. manshurica display differences morphologically (Figures 4 and 5), wherein the *P. manshurica-*contaminated cellulosic biofilm lost its reticulated structure, and nanofiber network, resulted in the formation of thick layers rather than nanofibers. Kim et al. (2006) stated that the vast surface area of cellulose was essential for the attachment of cellulose and the development of vascularization [29]. In addition, within the range of 10–100 nm, cotton fibers, wood slurry fiber, and synthetic fiber were indicated by Youshinaga et al. [30], which could be linked to the unique and fine structure of the cellulosic biofilms from G. oxydans.
4.3. The process of staining biofilm-populated biomaterial using crystal violet
The staining technique for the microscopic examination of biofilms offers the benefit of efficiently tracking the development of biofilms on physical substrates. The present investigation employs a combination of qualitative and semiquantitative methodologies to identify biofilms through an advanced stereomicroscope, which enables a comprehensive analysis of the structure and function of biofilms. In addition, the utilization of mosaic technology has the potential to be implemented for the examination of extensive surfaces extending square centimeters, in contrast to conventional methods which focus on considerably smaller regions.
Basically, crystal violet is a cationic dye commonly utilized in microbiology to label negatively charged surface molecules such as peptidoglycan and extracellular polysaccharides [31]. To mitigate the potential for manipulation and artifacts resulting from the bacteria and their biofilm, a direct application of crystal violet staining was employed on the biofilm surfaces. The prominent technique to evaluate biofilm formation generally involves the use of a 96-well microtiter plate assay [32], wherein the detection of the dye taken from the formerly stained biofilm was typically accomplished through colorimetric means. Although crystal violet staining is generally applied in a microtiter plate assay [33,34], it was carried out in this study by directly staining the biofilm sample taken from the vinegar-producing environment and examined using brightfield microscopy (Figure 6).
A previous study demonstrated that bacterial biofilm structures consist of an extracellular matrix containing long-chain polymeric molecules, mainly negatively charged exopolysaccharides and lipopolysaccharides, and secreted by related cells [35]. Negatively charged surface molecules in the EPS structure and cells were visualized using crystal violet staining to identify biofilm-producing organisms and further visualize the biofilm formations. Accordingly, the images indicated that the filaments were stacked on top of each other and formed a structure in which bacterial and yeast cells were observed locally, which could be distinguished considering their morphologies, including rod-shaped (i.e. P. manshurica yeast) and spherical-shaped (i.e. G. oxydans bacteria) (Figures 6A and 6B). In addition, P. manshurica biofilm formation was achieved to be observed from a vinegar-producing environment stained with crystal violet under the bright field microscope, according to the dense structure of biofilm and hence the dye density.
Given that SEM images and crystal violet staining are not able to identify bacterium strains as part of a biofilm, our technique was combined with MALDI-TOF MS and FT-IR analysis. However, the limitations of crystal violet staining in identifying bacterial strains within biofilms necessitate the use of molecular assays to achieve more accurate identification of the strains involved in our approach. Therefore, further investigation is required to establish a precise relationship between colony-forming unit counts and/or polymerase chain reaction analysis. This will enable the conversion of our qualitative read-out into more quantitative data.
5. Conclusion
The presence of Pichia species has been observed to result in the development of biofilm and malodorous compounds on the surface of the bacterial cellulosic film during production. As a result, yeast contamination may develop in the process of vinegar manufacturing and may give rise to complications. Based on the experimental results of the detection and characterization studies, including MALDI-TOF, FT-IR, SEM, and crystal violet staining, this study highlights the potential detrimental effect of Pichia proliferation on the quality of homemade vinegar during apple fermentation, due to its unique biofilm characteristics. Consequently, this study emphasizes that the proliferation of Pichia during apple fermentation can potentially have a negative impact on the quality of the homemade vinegar, owing to its distinct biofilm characteristics, and therefore necessitates identifying the detection and characteristic features belonging to the microbial source.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Hasan AN Antony U Bala GV Prabha DT Kavya C Study on Production Vinegar from Apple IOP Conference Series: Earth and Environmental Science 2021761101212510.1088/1755-1315/761/1/012125 · doi ↗
- 2Morales ML Tesfaye W García Parrilla MC Casas JA Troncoso AM Sherry wine vinegar: Physicochemical changes during the acetification process Journal of Science Food and Agriculture 200181761161910.1002/jsfa.853 · doi ↗
- 3Ho CW Lazim AM Fazry S Zaki UKHH Lim SJ Varieties, production, composition and health benefits of vinegars: A review Food Chemistry 20172211621163010.1016/j.foodchem.2016.10.12827979138 · doi ↗ · pubmed ↗
- 4Mathew B Agrawal S Nashikkar N Bundale S Upadhyay A Isolation of Acetic Acid Bacteria and Preparation of Starter Culture for Apple Cider Vinegar Fermentation Advances in Microbiology 20199610.4236/aim.2019.96034 · doi ↗
- 5Štornik A Skok B Trček J Comparison of cultivable acetic acid bacterial microbiota in organic and conventional apple cider vinegar Food Technology and Biotechnology 201654111311910.17113/ftb.54.01.16.408227904401 PMC 5105631 · doi ↗ · pubmed ↗
- 6Cleenwerck I De Vos P Polyphasic taxonomy of acetic acid bacteria: An overview of the currently applied methodology International Journal of Food Microbiology 2008125121410.1016/j.ijfoodmicro.2007.04.01718237808 · doi ↗ · pubmed ↗
- 7Hou X Wang J Mei Y Ge L Qian J Antibiofilm mechanism of dielectric barrier discharge cold plasma against Pichia manshurica Innovative Food Science & Emerging Technologies 20238510.1016/j.ifset.2023.103340 · doi ↗
- 8Lorenzini M Cappello MS Andreolli M Zapparoli G Characterization of selected species of Pichia and Candida for their growth capacity in apple and grape must and their biofilm parameters Letters in Applied Microbiology 2023761 ovac 02810.1093/lambio/ovac 02836688769 · doi ↗ · pubmed ↗