Spindle checkpoint activation by fungal orthologs of the S. cerevisiae Mps1 kinase
Amy Fabritius, Anabel Alonso, Andrew Wood, Shaheen Sulthana, Mark Winey, Michael Polymenis, Michael Polymenis, Michael Polymenis

TL;DR
Researchers explored using Mps1 kinase from pathogenic fungi in a budding yeast system to develop new antifungal drugs.
Contribution
The study adapts a budding yeast system to express Mps1 orthologs from pathogenic fungi for antifungal drug screening.
Findings
Overexpression of Mps1 orthologs from pathogenic fungi caused cell cycle arrest in budding yeast.
Two fungal Mps1 orthologs produced phenotypes similar to budding yeast Mps1, validating the assay system.
The system shows potential for identifying inhibitors of Mps1 enzymatic activity as antifungal agents.
Abstract
There is an ongoing need for antifungal agents to treat humans. Identification of new antifungal agents can be based on screening compounds using whole cell assays. Screening compounds that target a particular molecule is possible in budding yeast wherein sophisticated strain engineering allows for controlled expression of endogenous or heterologous genes. We have considered the yeast Mps1 protein kinase as a reasonable target for antifungal agents because mutant or druggable forms of the protein, upon inactivation, cause rapid loss of cell viability. Furthermore, extensive analysis of the Mps1 in budding yeast has offered potential tactics for identifying inhibitors of its enzymatic activity. One such tactic is based on the finding that overexpression of Mps1 leads to cell cycle arrest via activation of the spindle assembly checkpoint. We have endeavored to adapt this assay to be based…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Fig 1
Fig 1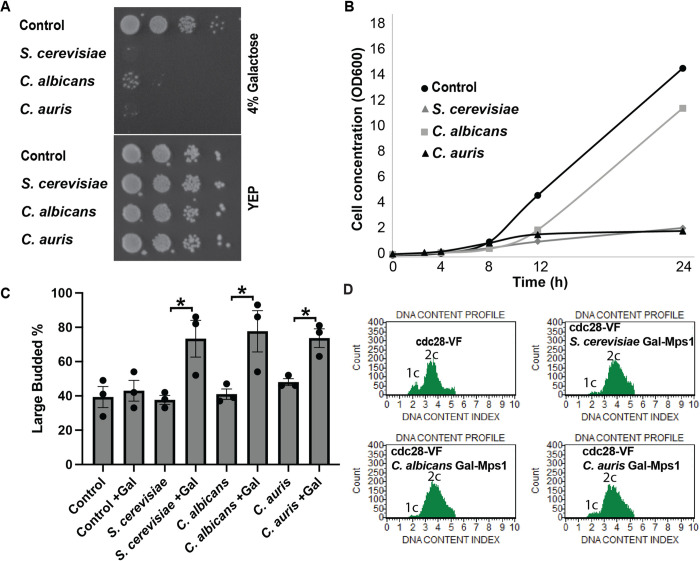 Fig 2
Fig 2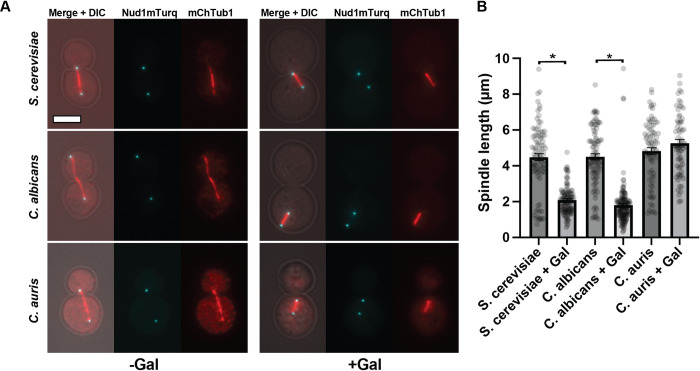 Fig 3
Fig 3- —http://dx.doi.org/10.13039/100000057National Institute of General Medical Sciences
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsFungal and yeast genetics research · Plant-Microbe Interactions and Immunity · Plant Reproductive Biology
Introduction
Fungal infections are on the rise for a variety of reasons and pose a serious health risk, especially for the increasing population of immune compromised patients. Currently, the rise in Candida auris hospital infections is raising alarms (CDC News Release: https://www.cdc.gov/media/releases/2023/p0320-cauris.html). Furthermore, fungal infections may be contributing to some disease states, such as cancers, in ways that have not been previously appreciated [1]. However, the collection of antifungal agents is limited, and few new antifungals have been reported in recent years. Antifungal targets generally fall into two classes: fungal specific (cell wall and ergosterol) and conserved, with enough divergence that the host is protected (e.g. microtubules), with most falling within the first class. Most new antifungals treatments are combination therapies or next generation formulations of current antifungals that attempt to alleviate side-effects, drug-drug interactions, or medication delivery issues [2]. However, some novel “first in class” compounds are being tested [3].
A challenge in development of new antifungals is the fact that fungi are eukaryotes such that host toxicity is a serious concern. However, the availability of genome sequences for numerous pathogenic fungi gives investigators the opportunity to test the endogenous forms of antifungal targets with the hope of tuning the agent for a target pathogen’s protein while limiting its recognition of the human ortholog if there is one. Additionally, most current antifungals suppress growth, but are not lethal to the fungi, which is one likely factor leading to the increase in drug resistance [4].
The yeast Mps1 kinase may be a good antifungal target because when inactivated by mutation or as a druggable form, the loss of activity leads to rapid cell death. This phenotype of Mps1 mutant strains is known to arise from the fact that the kinase is required at multiple points during the cell cycle to promote the assembly of the mitotic spindle and it is required in the spindle assembly checkpoint that arrests cell cycle progression in response to defects in the mitotic spindle. On the assembly side, Mps1 is needed for SPB duplication and for kinetochore assembly. On the checkpoint side, Mps1 contributes to mitotic checkpoint complex (MCC) formation by regulating localization and catalyzation of the MCC assembly (along with the SAC scaffolding proteins, BUB1 and MAD1-MAD2), blocking the APC/C and anaphase onset until spindle microtubules are properly attached at the kinetochores (Reviewed in [5]). Mps1 inactivation leads to failure in these dual roles leading to catastrophic events wherein cells do not form functional bipolar spindles yet fail to arrest in mitosis in response to this defect such that the cells continue through a failed mitosis that generates inviable aneuploid cells. The recognition that cells can be rapidly killed by inactivating Mps1 has led to it being considered a potential target for cancer chemotherapies in addition to our and others’ [6] interest in adapting it as an antifungal target.
In addition to showing that inactivation of Mps1 is lethal to yeast cells, we have also shown that overexpression of Mps1 inhibits cell growth. Analysis of the cells whose growth was inhibited by Mps1 overexpression revealed that the cells arrested in mitosis as large-budded cells with metaphase mitotic spindles [7]. This phenotype is the result of Mps1 overexpression constitutively activating the spindle checkpoint causing a mitotic arrest. As discussed above, Mps1 is known to act in the checkpoint. This “gain-of-function” phenotype for Mps1 offers the opportunity to test potential inhibitors in a simple growth assay wherein the inhibition of overexpressed Mps1 would restore growth to the cells. We have demonstrated the utility of this concept using a druggable allele of Mps1. Mps1-as1 (analog-sensitive) has an engineered ATP binding site that allows binding of an ATP analog that will not bind other protein kinases [8]. Overexpression of Mps1-as1 arrests cell growth as expected, and treatment of these cells with the inhibitor restores growth. Using the system, we have validated this positive-growth, whole cell assay for Mps1 activity.
In this study, we have asked whether the Mps1 overexpression assay could be recapitulated with the overexpression of Mps1 orthologs from other organisms. In total, we tested overexpression of eight different fungal Mps1 genes, four of which are the Mps1 orthologs in human pathogens. We also tested the Mps1 orthologs from two yeasts that used in research, one of which is a plant pathogen, and two chytrid fungi. The Mps1 orthologs from two Candida strains, C. auris and C. albicans, were found to be toxic when overexpressed in budding yeast, but the others were not. This first analysis of heterologous Mps1 expression suggests that this approach has promise in that some Mps1 orthologs could recapitulate the cell cycle arrest seen with endogenous Mps1.
Results
Identification of Mps1 orthologs
Orthologs of the Mps1 protein kinase are found throughout most of the eukaryotic world (Fig 1). The general organization of these proteins is a long N-terminal extension that exhibits little conservation across phylogenies, a conserved protein kinase domain and a short C-terminal extension. The Mps1 protein kinase domain has some signature motifs and is in the general family of S/T/Y kinases [9]. For this study, we intended to focus on Mps1 protein kinase encoding genes from pathogenic yeast. The genomes of many such organisms are available and were searched as described in the materials and methods. The apparent Mps1 ortholog was identified in seven pathogenic yeasts or other fungi. The multiple alignment of the protein kinase domain in these genes is shown in S1 Fig.
Mps1 kinase phylogenetics.Cladogram (guide) comparison of Mps1 kinase domains * indicates toxicity when overexpressed.
DNA sequences encoding each of the Mps1 orthologs, codon-optimized for expression in S. cerevisiae, were synthesized and inserted into a plasmid, enabling expression in yeast from the inducible galactose promoter (see materials and methods). Expression of these genes from integrated constructs was confirmed by immunoblotting for the GFP tag and measuring GFP fluorescence intensity (S2 Fig). While GFP-Mps1 expression among different constructs and backgrounds was variable, the expression was consistently increased in the +Gal condition.
Functional analysis of Mps1 orthologs in S. cerevisiae
As described above, a widely conserved function of Mps1 is its role in the spindle assembly checkpoint. The observation that Mps1 overexpression in S. cerevisiae led to a mitotic cell cycle arrest was an early indication of this function. The arrest is dependent on the spindle assembly checkpoint and arises from activation of the pathway which leads to high CDK kinase activity by inhibiting cyclin degradation by the APC. However, the arrest is transient because the cells are able to inhibit CDK (Cdc28) kinase activity via Swe1-mediated inhibitory phosphorylation. Overexpression of S. cerevisiae Mps1 in cdc28-VF strains, in which Cdc28 is insensitive to Swe1 inhibition is lethal [10] (Fig 2). We wished to determine whether any of the Mps1 orthologs were toxic in cdc28-VF strains upon overexpression. We found that, in addition to S. cerevisiae Mps1, overexpression of the Mps1 orthologs from either Candida albicans or Candida auris was toxic in this S. cerevisiae strain in a plate assay (Fig 2A). The continued presence of some minimal growth (Fig 2A and 2B) in the C. albicans Mps1 strain may be due to cell variability and possibly loss of Mps1 overexpression in a subset of cells.
*Overexpression of Mps1 is toxic and causes a cell-cycle arrest in the cdc28-VF background.All in cdc28-VF background, A. Colony growth on 4% galactose plates (Mps1 overexpression) and YEP plates (-galactose control)–dilution series. B. Growth curve of cells overexpressing Mps1 (4% galactose) of indicated species grown in liquid media. C. Percentage of large budded cells when overexpressing Mps1 of indicated species. D. Flow cytometric analysis of DNA content in cdc28-VF cells overexpressing indicated Mps1, showing cell-cycle arrest as single (2c) peak measured at 5 hours. p<0.05.
Inhibition of growth in a plate assay does not reveal the basis of the overexpressed gene’s toxicity. We previously showed that S. cerevisiae Mps1 overexpression inhibited cell growth by causing a large budded mitotic cell cycle arrest. We asked whether overexpression of the C. auris or C. albicans Mps1 orthologs caused the same phenotypes. Fig 2 shows that overexpression of these Mps1 orthologs does inhibit cell growth based on optical density (Fig 2B) and that the cells arrest with large buds (Fig 2C). Finally, the spindle assembly mediated mitotic arrest is accompanied by cells containing G2/M post DNA synthesis quantities of DNA. Fig 2D shows the DNA content of cell populations as determined by flow cytometry. The cdc28-VF cells alone grow normally, but they do exhibit a bias toward mitotic G2/M DNA content. Nonetheless, a small population of G1 cells can be detected (Fig 2D, upper left panel). Overexpression of S. cerevisiae Mps1 in these cells eliminates this small peak, resulting in all of the cells having G2/M DNA content as expected for a mitotic arrest (Fig 2D upper right panel). This same result is observed in the cells overexpressing the C. albicans or C. auris Mps1 orthologs (Fig 2D, lower panels).
The complete panel of plate growth tests for all the strains expressing the various orthologs is shown in S3 Fig. We show growth with Mps1 overexpression in cdc28-VF strains, in addition to controls in wild-type cells and in a strain containing fluorescent protein markers for components of the microtubule cytoskeleton. The majority of the expressed orthologs cause no growth deficiency on plates. Furthermore, these orthologs did not cause a large-budded arrest (S3B Fig) or inhibit growth in liquid culture as assessed by optical density (S3C Fig).
Based on previous studies, we expected S. cerevisiae Mps1 to cause an arrest phenotype, but it appears that Mps1 from near relatives, but not of other fungal species, is able to trigger the arrest in this system. It is unclear whether the lack of phenotype by the overexpression of other species of Mps1 protein is due to their overexpression being insufficient or due to the S. cerevisiae host’s inefficient recognition of the Mps1 substrates. However, because there was an arrest in some species, it suggests the system can work as a test for whether Mps1 could be a potential antifungal target for these species.
An Mps1 inhibitor has, in fact, been suggested as an antifungal for C. albicans. Use of the inhibitor decreased phosphorylation activity and reduced growth of fungal cells, while not affecting human Mps1 activity [6], an important factor in the design of antifungal treatments.
Cellular structure impact of Mps1 ortholog overexpression
As discussed above, overexpression of S. cerevisiae Mps1 leads to mitotic arrest, but we have also reported that overexpression of a kinase-dead allele of Mps1 is toxic [8]. Furthermore, overexpression of the inactive Mps1 kinase has severe detrimental effects on spindle pole bodies and microtubule organization [11]. To assess the impact of Mps1 ortholog overexpression on cells, we overexpressed these constructs in strains containing fluorescent protein tagged microtubules (mCherry-Tub1) and spindle pole bodies (Nud1-mTurq). As expected, the overexpression of the Mps1 ortholog from C. auris or C. albicans caused a bi-polar, short spindle arrest as expected based on the above phenotypes that correspond to a mitotic arrest (Fig 3A and 3B). While Cdc28 is still presumably active in this background (mCherry-Tub1;Nud1-mTurq), the arrests may be due to a synthetic effect between the Mps1 overexpression and tagged cytoskeletal components.
*Spindles in wild-type cells overexpressing Mps1 of indicated species.A. Examples of long (left) and short (right) spindles in cells overexpressing Mps1 of indicated species in cells expressing mCherry-Tub1 (tubulin) and Nud1-mTurquoise (SPB marker). B. Average spindle lengths (Nud1 pole to pole length) of cells overexpressing Mps1 of indicated species. Scale bar = 4μm. p<0.001.
Examination of mitotic spindles and spindle pole bodies in the strains not exhibiting toxicity or a cell cycle arrest upon overexpression of a Mps1 ortholog also did not reveal defects in spindle structures (e.g. Fig 3A).
Conclusion
We report a cell-based assay for the function of Mps1 protein kinases encoded by orthologs from pathogenic yeasts. Conceptually similar to a “humanized mouse,” we are using the innocuous S. cerevisiae to test gene function from pathogens. The assay is based on understanding the function of Mps1 in the spindle checkpoint of budding yeast. We have previously reported that Mps1 over-expression inhibits cell cycle progression by ectopically activating the spindle assembly checkpoint. This phenotype can be reproduced by over-expression of Mps1 from Candida species that are closely related the budding yeast, Saccharomyces cerevisiae. These Candida Mps1 orthologs include the gene from C. auris, a pathogenic yeast of growing concern as an infectious agent.
Materials and methods
Ortholog identification and comparison
Orthologs of S. cerevisiae Mps1 were identified by doing a BLASTp search targeted to the database for the organism of interest, and a reciprocal best hit protein BLAST search was used to confirm the identity. Kinase domains were determined based on amino acid sequence alignment to the known kinase domain of S. cerevisiae Mps1 [9]. Clustal omega was used to create a guide cladogram, with a real branch lengths comparison of kinase domain sequences.
Gene synthesis and vector design
Double stranded DNA of codon optimized (idtdna.com/CodonOpt) mps1 of the indicated species was synthesized (IDT) and cloned into the XhoI and XbaI or XmaI sites of pSJ114 [12].
Yeast strains, transformation, culture
Yeast strains used in this study (Table 1) are derived from the w303 background. Plasmids were digested with ApaI and integrated into the URA3 locus.
Table 1: Strains and plasmid.
All yeast strains were grown at 30°C. Strains were grown to log phase in YEP containing 2% raffinose, then adjusted to OD600 of 0.1–0.4. For plate assays, tenfold serial dilutions were spotted onto glucose (repressing) or 4% galactose (expressing)(+Gal) plates. Plates were incubated at 30°C for 2–3 days.
For all other assays, galactose was added to 4% (Gal+) to induce GFP-Mps1 expression and cells were assayed after 5 hours of growth in galactose-containing medium.
Cell cycle profiling
The cell cycle stage was determined using the Muse Cell Analyzer instrument and Muse Cell Cycle Kit (Millipore Sigma) according to the manufacturer’s instructions.
For the budding index, cells were sonicated and were counted using a hemacytometer (N>50 cells per condition). Spindle length was measured as distance between Nud1mTurq foci (N>50 cells per condition). Statistical tests were done using a Student’s t-test and error bars show the SEM.
Fluorescence microscopy
Briefly, cells were fixed in 4% formaldehyde in 100 mM sucrose. Cells were sonicated, then images were acquired on an inverted microscope Eclipse Ti-E (Nikon) fitted with a CFI60 Plan Apochromat lambda 100x oil immersion objective lens (N.A. 1.45) (Nikon) and an ORCA-Flash4.0 V3 sCMOS camera. Data were acquired using NIS-Elements software. Image projections were made in FIJI [13,14].
Fluorescence intensity of GFP of a 0.65x0.65 μm (10x10 pixel) region between SPBs of short spindles was measured using FIJI (N = 10 cells per condition). Statistical tests were done using a Student’s t-test and error bars show the SEM.
Immunoblot analysis
Cells were collected by low-speed centrifugation, washed, and resuspended in B150 breaking buffer. Cells were then lysed by vortexing with glass beads for 10 min at 4°C. Extracts were clarified by centrifugation, and samples were run on an 8% SDS-PAGE gel. The membrane was blocked in 0.2% Tropix I-block reagent (Applied Biosystems) with 0.1% Tween-20 in PBS overnight at 4°C. The GFP tag was detected using a mouse monocolonal anti-GFP antibody (Biolegend) [14].
Supporting information
S1 FigAmino acid sequence alignment.Amino acid sequence alignment of Mps1 kinase domains from indicated species.(TIF)
S2 FigProtein expression.Mps1 Protein expression with (+) or without (-) galactose induction shown by A. Western blot in indicated cell background. Anti-GFP showing overexpression of indicted Gal-GFP-Mps1 construct. * = GFP-Mps1, arrows = internal control. B. GFP fluorescence intensity in nucleus measured from microscopy of cells expressing GFP-Mps1 in mCh-Tub1;Nud1-mTurq background **p<0.001.(TIF)
S3 FigGrowth and viability with overexpression of Mps1.A. Growth on dilution plates of cells overexpressing indicated species of Mps1 (4% galactose) or without overexpression (YEP) in three different cell backgrounds (cdc28-VF, WT, or mChTub1;Nud1mTurq). B. Budding indices with (+Gal) or without overexpression of indicated species of Mps1 in either a wild-type or mChTub1;Nud1mTurq background. C. Growth curve (OD_600_) of cells overexpressing (4% galactose) indicated Mps1 in liquid culture in cdc28-VF background. *p<0.05.(TIF)
S1 Raw images(TIF)
S1 Raw data(PDF)
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Narunsky-Haziza L, Sepich-Poore GD, Livyatan I, Asraf O, Martino C, Nejman D, et al. Pan-cancer analyses reveal cancer-type-specific fungal ecologies and bacteriome interactions. Cell. 2022;185: 3789–3806.e 17. doi: 10.1016/j.cell.2022.09.005 36179670 PMC 9567272 · doi ↗ · pubmed ↗
- 2Jacobs SE, Zagaliotis P, Walsh TJ. Novel antifungal agents in clinical trials. F 1000 Research. 2022;10: 507. doi: 10.12688/f 1000 research.28327.2 35136573 PMC 8787557 · doi ↗ · pubmed ↗
- 3Treviño-Rangel R de J, González GM, Montoya AM, Rojas OC, Elizondo-Zertuche M, Álvarez-Villalobos NA. Recent Antifungal Pipeline Developments against Candida auris: A Systematic Review. J Fungi. 2022;8: 1144. doi: 10.3390/jof 8111144 36354911 PMC 9695453 · doi ↗ · pubmed ↗
- 4Lee Y, Puumala E, Robbins N, Cowen LE. Antifungal Drug Resistance: Molecular Mechanisms in Candida albicans and Beyond. Chem Rev. 2021;121: 3390–3411. doi: 10.1021/acs.chemrev.0c 00199 32441527 PMC 8519031 · doi ↗ · pubmed ↗
- 5Mc Ainsh AD, Kops GJPL. Principles and dynamics of spindle assembly checkpoint signalling. Nat Rev Mol Cell Biol. 2023 [cited 21 Apr 2023]. doi: 10.1038/s 41580-023-00593-z 36964313 · doi ↗ · pubmed ↗
- 6Tsuda K, Nishiya N, Umeyama T, Uehara Y. Identification of LY 83583 as a specific inhibitor of Candida albicans MPS 1 protein kinase. Biochem Biophys Res Commun. 2011;409: 418–423. doi: 10.1016/j.bbrc.2011.05.010 21586273 · doi ↗ · pubmed ↗
- 7Hardwick KG, Weiss E, Luca FC, Winey M, Murray AW. Activation of the budding yeast spindle assembly checkpoint without mitotic spindle disruption. Science. 1996;273: 953–956. doi: 10.1126/science.273.5277.953 8688079 · doi ↗ · pubmed ↗
- 8Jones MH, Huneycutt BJ, Pearson CG, Zhang C, Morgan G, Shokat K, et al. Chemical Genetics Reveals a Role for Mps 1 Kinase in Kinetochore Attachment during Mitosis. Curr Biol. 2005;15: 160–165. doi: 10.1016/j.cub.2005.01.010 15668173 · doi ↗ · pubmed ↗