Oleispirillum naphthae gen. nov., sp. nov., a bacterium isolated from oil sludge, and proposal of Oleispirillaceae fam. nov
Chenghui Peng, Xue Zhang, Jiang Li, Min Yang, Shichun Ma, Hui Fan, Lirong Dai, Lei Cheng

TL;DR
Scientists discovered a new spiral-shaped bacterium from oil sludge and classified it as a new species, genus, and family.
Contribution
The discovery and classification of a novel bacterium, Oleispirillum naphthae, and its placement in a new family Oleispirillaceae.
Findings
Strain Y-M2T has unique growth conditions and fatty acid composition.
The bacterium possesses a nitrogen-fixation cluster similar to methanogens.
Phylogenetic and genomic analyses confirmed it belongs to a new family in Rhodospirillales.
Abstract
A microaerophilic, Gram-negative, motile, and spiral-shaped bacterium, designated Y-M2T, was isolated from oil sludge of Shengli oil field. The optimal growth condition of strain Y-M2T was at 25 °C, pH 7.0, and in the absence of NaCl. The major polar lipid was phosphatidylethanolamine. The main cellular fatty acid was iso-C17 : 0 3-OH. It contained Q-9 and Q-10 as the predominant quinones. The DNA G+C content was 68.1 mol%. Strain Y-M2T showed the highest 16S rRNA gene sequence similarity to Telmatospirillum siberiense 26-4bT (91.1 %). Phylogenetic analyses based on 16S rRNA gene and genomes showed that strain Y-M2T formed a distinct cluster in the order Rhodospirillales. Genomic analysis showed that Y-M2T possesses a complete nitrogen-fixation cluster which is phylogenetically close to that of methanogene. The nif cluster, encompassing the nitrogenase genes, was found in every…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
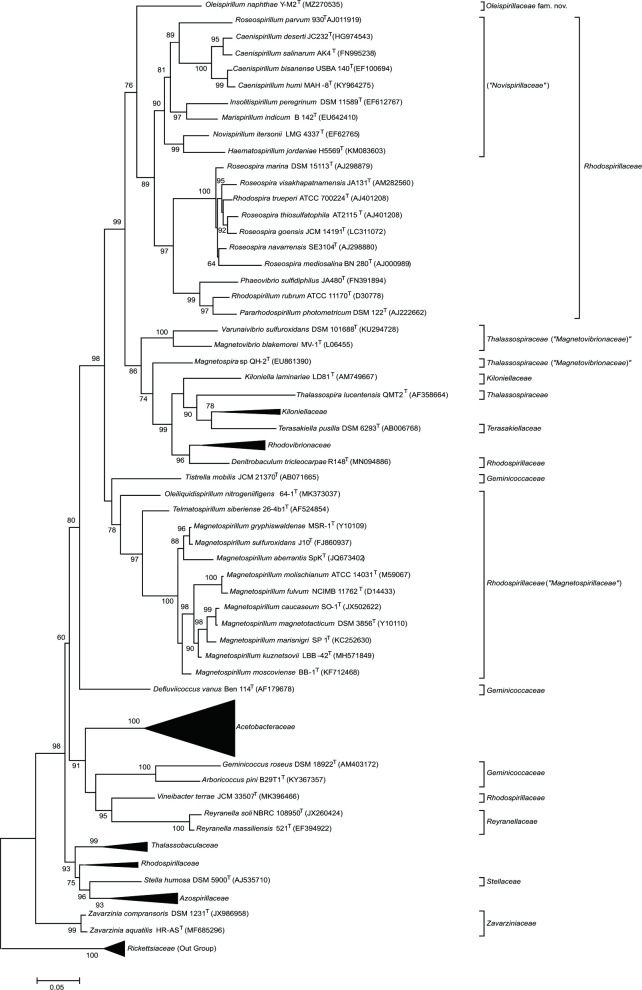 Fig. 1
Fig. 1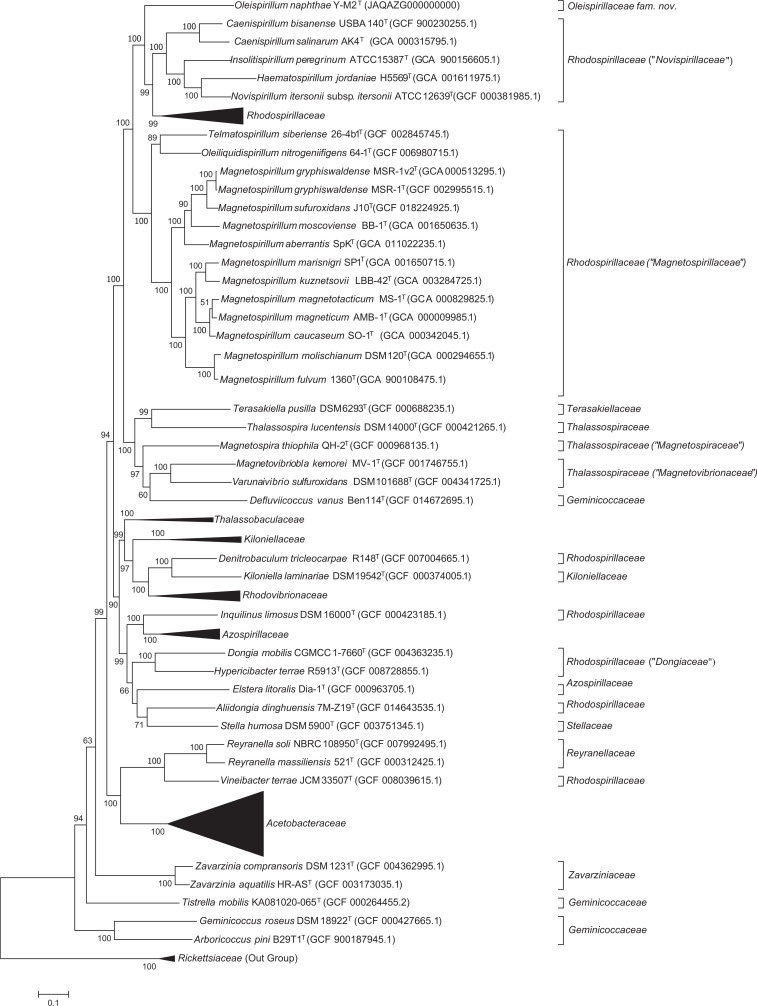 Fig. 2
Fig. 2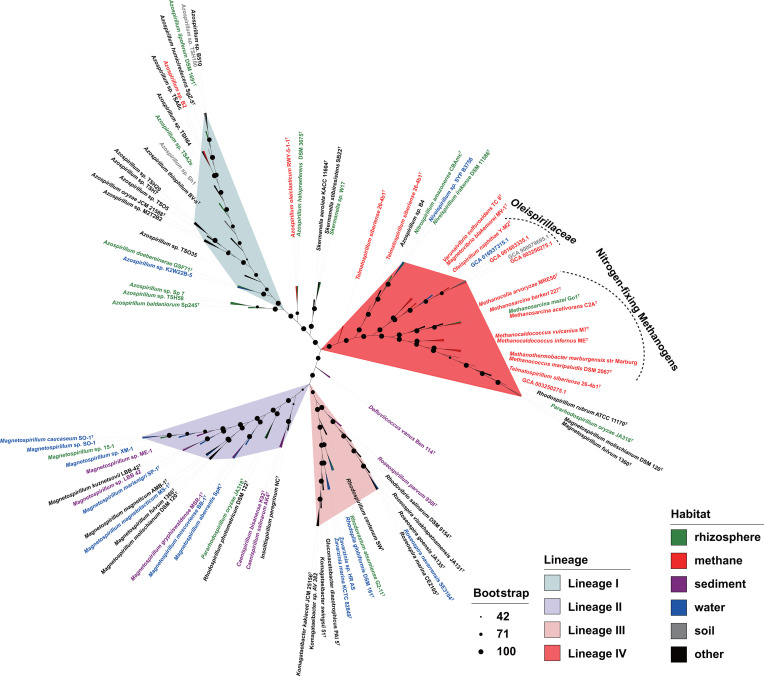 Fig. 3
Fig. 3| Characteristic | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 |
| Habitat | Oil sludge | Mesotrophic fen | Mud | Vinegar plant | Wheat | Biofilter | Marine macroalga | Freshwater | Salterns and salt lakes | Soil, freshwaters | Marine shellfish | Coastal seawater | Sea water | Mud |
| Gram reaction | Negative | Negative | Negative | Negative | Negative | Negative | Negative | Negative | Negative | Negative |
| Negative | Negative | Negative |
| Morphology | Spiral | Vibrio, spirilla | Helical spirilla | Rod | Vibroid | Coccoid | Spiral | Rod | Vibrioid, spiral | Six-pronged stars | Spiral | Slightly curved rod | Vibrioid, spiral | Straight rod, curved rod |
| Motility | − | + | + |
|
|
| + | − | + | − | + | + | + | + |
| Cells (μm) | 0.8–3.0×0.4–0.6 | Diameter 0.2–0.6 | 0.2–0.4×3–4 | 0.6–0.9×1.0–4.0 | 1.0–1.7 | 1.5–2.1 | 0.5–0.6×2.5–5.0 | 0.68–0.92×1.16–1.91 | 0.8–0.9×1.0–3.5 | Diameter 0.7–3.0 | 0.3–0.5×1.2–4.0 | 0.3–0.5×1.3–1.5 | 3–5×0.6 | 0.7–0.9×0.5–2.1 |
| Temperature range (optimum) for growth (°C) | 20–30 (25) | 4–30 (25-28) | 18–38 ( | 15–45 (30–35) | 4–40 (25) | 18–37 (30–35) | 20–45 (42) | 6–40 (32) | 10–35 (30–35) | 4–40 ( |
| |||
| pH (optimum) for growth | 7.0 (6.0–7.5) | 4–7 (5.7–6.0) | 1.5–1.5 ( | 5.7–6.8 ( | 5.5–11.0(8) | 3.5–9.5 (3) |
| Neutral | 6.0–9.0 ( | 7–9 (8) |
|
| ||
| NaCl range (optimum) for growth (%,w/v) | 0 ( |
|
|
|
|
| 0.3–10.0 ( |
| 3–24 (8–12) | Up to 1 | 0.5–8.0 ( | 1–10 ( | 2–10 ( |
|
| DNA G+C content (mol%)* | 68.07 | 61.5–64.0 | 67.2 | 56.2–57.2 | 69–70 | 60.2–60.4 | 51.1 |
| 69.1 | 69.3–72.9 | 48/51 | 68 | 54.7 | 66.1 |
| API 20NE assay: | ||||||||||||||
| + |
|
|
|
|
|
|
|
|
|
|
|
|
| |
| − | − |
| − | + (to nitrite) | + | + (to N2O) | + |
|
| + (to nitrite) | + (to nitrite) |
|
| |
| + |
|
|
|
| + | − |
|
|
|
|
|
|
| |
| + |
| − |
|
|
|
| + |
|
| − |
| − | − | |
| Oxidase | − | − | + | − | + | + | + | + |
| + | + | + | + |
|
| Catalase | − | − | + | + |
| + | + | − |
| + | − | + | + |
|
| Fatty acid | Summed feature 8, summed feature 3, iso-C15 : 0 3-OH, iso-C17 : 0 3-OH | C18 : 1 | Summed feature 8, summed feature 3, C16 : 0 |
|
|
| C18 : 1 |
| C18 : 1, C18 : 0 |
| C14 : 0 3-OH, C18 : 1, C16 : 1, C16 : 0 | C16 : 0, C18 : 1 | C14, C16 : 0 3-OH, C16 : 1 | C18 : 1, C16 : 0, C18 : 0 3-OH, C16 : 0 3-OH |
| Quinone | Q9, Q10 |
| Q10 | Q-9 |
|
|
|
| Q-10, MK-10 |
| Q-10 | Q-10 |
| Q-10 |
| Characteristics |
|
|
|
|
| Genera | 1 | 14 | 5 | 5 |
| Oxygen requirement | Microaerophilic | Strictly aerobic, aerobic or facultatively anaerobic | Anaerobic | Aerobic or microaerobic |
| Shape | Spiral | Rod, curved rod, vibrioid or spiral | Helical | Spirillum, vibrioid to spiral |
| Motility | + | +/− | + | +/− |
| Gram stain | − | − | − | − |
| Flagellation | Single polar | Single polar or bipolar tufts of flagella (two to five fibrils) | Single polar or bipolar | Polar or subpolar flagella |
| Oxidase | − | +/− | +/− | +/− |
| Catalase | − | +/− | + | +/− |
| Growth temperature (°C): | ||||
| 20–30 | 10–45 | 10–47 | 4–55 | |
| 25 | 25–40 | 25–37 | 25–45 | |
| NaCl concentration for growth (%): | ||||
| 0 | 0–15 | 0–12 | 0–3 | |
| 0 | 0–8 | 0.5–8.0 | − | |
| pH for growth: | ||||
| 7.0 | 5.0–11.0 | 6.5–10.0 | 4.0–10.0 | |
| 6.0–7.5 | 7.0–9.0 | 7.0–8.0 | 5.7–7.5 | |
| Gelatin hydrolysis | + | +/− | − | − |
| Urease | + | +/− | +/− | − |
| H2S production |
| − | +/− | + |
| Electron acceptors (+) |
| Sulphate and nitrate | Nitrate | Nitrate |
| Utilization of sugars | + | + | + | + |
| Utilization of amino acids | + | + | + | − |
| Utilization of organic acids | + | + | + | + |
| Respiratory quinones | Q9, Q10 | Q-7, MK-7, Q8, RQ8, Q9, Q10 | Q-9, Q-10 | Q-9, MK-9, Q-10 |
| Genome size (Mb) | 3.2 | 2.1–5.4 | 2.4–4.6 | 3.8–6.2 |
| DNA G+C content (mol%) | 68.1 | 59.1–69.3 | 62.4–70.0 | 61.6–66.4 |
- —National Nature Science Foundation of China
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGenomics and Phylogenetic Studies · Microbial Community Ecology and Physiology · Metal Extraction and Bioleaching
Introduction
Members of the order Rhodospirillales, affiliated with the class Alphaproteobacteria within the phylum Pseudomonadota, are ubiquitous in soil [12], air [3], water [45], anode biofilms [6], crude oil-contaminated soil [78], respiratory secretions [9], and other niches [1013]. The order Rhodospirillales comprises 12 validly published families which were reclassified by Hördt et al. in 2020 [14]. However, six more novel families were separated from the existing families within this order according to the phylogenomic analyses [15]. The family Rhodospirillaceae was split into family ‘Dongiaceae’ (including genera Dongia, Aliidongia, and Hypericibacter), ‘Oceanibaculaceae’ (Oceanibaculum), and current Rhodospirillaceae (Pararhodospirillum, Phaeovibrio, Rhodospira, Rhodospirillum and Roseospirillum) [15]. Most members of this order are chemoorganoheterotrophic and/or photoorganoheterotrophic [1618]. It also contains free-living nitrogen-fixing bacteria, such as the genus Azospirillum in the family Azospirillaceae [131920]. Most of them were obtained from plant root-related environments, including the facultative anaerobes Azospirillum oryzae COC8^T^ [19] and Azospirillum lipoferum Sp 7^T^ [621]. There are several nitrogen-fixing bacteria within Azospirillum (Azospirillum palustre B2^T^, Azospirillum oleiclasticum RWY-5-1-1^T^ and Azospirillum rugosum IMMIB AFH-6^T^) and aerobic Oleiliquidispirillum nitrogeniifigens 64-1^T^ in the family Rhodospirillaceae isolated from oil-related environments, including oil-contaminated soil and oil production mixtures [7813], indicating that members of Rhodospirillales are probably involved nitrogen cycling in oilfield.
Biological nitrogen fixation, in which micro-organisms convert atmospheric nitrogen gas (N_2_) to ammonia (NH_3_), is an important mechanism to support bio-available nitrogen for organisms. Additionally, it plays a critical role in the global nitrogen cycle. It has been shown that this process is mainly mediated by three different nitrogenases, molybdenum–iron nitrogenase (Nif, Mo-Fe), vanadium–iron nitrogenase (Vnf, V-Fe), and iron-only nitrogenase (Anf, Fe-Fe) [2223]. Nif has two protein components: an electron delivery component (NifH, encoded by nifH) and a catalytic component (NifD and NifK, encoded by nifD and nifK, respectively) [23]. Nif is the most important nitrogen-fixation protein and is widely distributed in archaea (phylum Euryarchaeota) and 13 phyla in the bacterial domain, primarily including Bacillota, Bacteroidota, Cyanobacteria and Pseudomonadota [23], which have been found in a variety of environments ranging from marine [2425], rhizosphere [2627], freshwater sediments [24], and oilfields [828]. Genomic studies have revealed that nitrogen fixation-related genes constitute a nitrogen fixation cluster, such as the 17 nif genes (nifQ, nifA, nifL, nifH, nifD, nifK, nifT, nifY, nifE, nifN, nifX, nifS, nifU, nifW, nifZ, nifM, and nifF) distributed in a 49 kb region in Stutzerimonas stutzeri A1501^T^ [2930]. Previous studies have demonstrated that all nitrogen fixation bacteria in the order Rhodospirillales carry nif gene clusters, and no vnf or anf cluster has been identified in these species [68,13, 19, 28, 31].
Isolation
Strain Y-M2^T^ was isolated from oil sludge collected from Shengli oil field in Shandong province, PR China (38° 39′ 28″ N 104° 04′ 59″ E) using a traditional dilution method with a 96-well microplate as described previously [32]. Mineral medium containing 9.0 g NaCl, 0.5 g KCl, 0.3 g NH_4_Cl, 0.2 g KH_2_PO_4_, 3.0 g MgCl_2_·6H_2_O, 0.15 g CaCl_2_·2H_2_O, 0.5 g l-cysteine hydrochloride, 2 ml TE284 buffer, and 0.001 g resazurin in 1000 ml distilled water was used for isolation. The medium was boiled and cooled under 99.999 % N_2_ flow, dispensed into vials or tubes with butyl rubber stoppers, then sterilized at 121 °C for 30 min. The initial culture (0.5 ml) was diluted 10-fold by adding into 4.5 ml medium supplemented with short-chain fatty acids, glucose, yeast extract, and tryptone. After 7–30 days of incubation at 25 °C in rectangular jar using AnaeroGen (Thermo Scientific), pure cultures were isolated and transferred into fresh liquid medium. The purified strain Y-M2^T^ was cultured at 25 °C under microaerobic conditions (10 % oxygen) in modified Reasoner’s 2A (R2A) broth containing 0.25 g tryptone, 0.5 g casein hydrolysate, 0.5 g yeast extract, 0.5 g soluble starch, 0.3 g K_2_HPO_4_, 0.1 g MgSO_4_, 0.3 g sodium pyruvate, 0.25 g peptone, 0.5 g glucose, and 1000 ml distilled water. The medium was prepared and dispensed under N_2_ with the addition of 10 % oxygen, and sterilized at 121 °C for 15 min.
Genome sequencing
Strain Y-M2^T^ grew in 300 ml modified R2A medium at 28 °C for 48 h and 180 r.p.m. was harvested for genomic DNA sequencing. A commercial bacterial genomic DNA extraction kit (Magen) was used to extract the genomic DNA of strain Y-M2^T^. The draft genome of strain Y-M2^T^ was sequenced and assembled using the Illumina NovaSeq sequencing platform (Beijing Novogene Technology Co., Ltd., Beijing, PR China). The SPAdes strategy was used to assemble the genome, and the resulting N20, N50, and N90 values. Genome quality and single-copy marker genes were retrieved using CheckM (version 1.1.0; (https://github.com/Ecogenomics/CheckM/wiki).
Genome features
The genome shotgun project of strain Y-M2^T^ has been deposited at DDBJ/ENA/GenBank under the accession JAQAZG000000000. The N20, N50, and N90 values of strain Y-M2^T^ were 461 460, 216 590, and 128 336, respectively. The draft genome sequence of strain Y-M2^T^ was 3 214 613 bp, contained 2903 open reading frames, 27 contigs, three rRNAs (one each of 5S rRNA, 16S rRNA, and 23S rRNA), 48 tRNA, and 69 ncRNA. The DNA G+C content was 68.1 mol % (Table S1, available in the online version of this article).
Phylogeny
The 16S rRNA gene sequence was retrieved from the genome (GenBank accession number MZ270535.1) and compared with the sequence obtained by amplifying with the universal bacterial primers 27F (5′-AGAGTTTGATCMTGGCTCAG-3′) and 1492R (5′- TACGGYTACCTTGTTACGACTT-3′) [33] to confirm the purity of strain Y-M2^T^. The 16S rRNA gene sequences of strains with a close taxonomic relationship were downloaded from the NCBI (www.ncbi.nlm.nih.gov) and LPSN databases (http://www.bacterio.net/). All sequences were aligned using muscle (version 3.8.1551) [34], and the phylogenetic trees were reconstructed using the software FastTree (version 2.1.11) with default parameters [35], and iq-tree (version 2.0.3) with ‘-m TEST -bb 1000 -alrt 1000’ [36].
The 16S rRNA gene sequence of strain Y-M2^T^ containing 1487 bp nucleotides was used for phylogenetic analyses, and the gene sequence similarity revealed that strain Y-M2^T^ was most closely related to T. siberiense 26-4b1^T^ (91.1 %) and Magnetospirillum gryphiswaldense MSR-1^T^ (90.6 %) in the family ‘Magnetospirillaceae’ (formerly belonging to Rhodospirillaceae) of the order Rhodospirillales. Based on the thresholds of sequence identity of 16S rRNA genes for separating new genera (94.5 %) [37], strain Y-M2^T^ is proposed to represent a novel genus in order Rhodospirillales. The maximum-likelihood phylogenetic tree based on 16S rRNA gene sequences showed that strain Y-M2^T^ located in the clade containing species of the family ‘Novispirillaceae’ and Rhodospirillaceae, but formed an independent branch which was well separated from all published families of the order Rhodospirillales (Fig. 1). The separation of strain Y-M2 ^T^ from all other valid families also occurred in the neighboir-joining and minimum-evolution phylogenetic trees (Fig. S1A and B). The average 16S rRNA gene sequence similarities from phylogenetic analysis between strain Y-M2^T^ and species within ‘Magnetospirillaceae’, ‘Novispirillaceae’, and Rhodospirillaceae were ≤90.8 % (Table S2), which was close to the minimum sequence identity for distinguishing families (87.7 %). Phylogenomic trees reconstructed using GTDB-tk (Fig. 2) and using concatenated alignment of 92 core genes (Fig. S1C) further confirmed that strain Y-M2^T^ is independent of ‘Magnetospirillaceae’, ‘Novispirillaceae’, Rhodospirillaceae, as well as other Rhodospirillales families, indicating it may represent a new family in order Rhodospirillales.
Phylogenetic trees of strain Y-M2T and Rhodospirillales members based on 16S rRNA genes. Species of the genus Rickettsia within the order Rickettsiales were used as an outgroup. Bootstrap percentages are based on 1000 replications.
Phylogenomic analyses of Rhodospirillales members based on 120 marker genes reconstructed by GTDB-tk.
Paired genomic average nucleotide identity (ANI) and average amino acid identity (AAI) were calculated using OrthoANIu [38] and Compare M (https://github.com/dparks1134/CompareM), respectively. Percentages of conserved proteins (POCP) between two microbial genomes were calculated according to the method described previously [39]. The AAI values between strain Y-M2^T^ and the genera of ‘Magnetospirillaceae’, ‘Novispirillaceae’, and Rhodospirillaceae were ≤60.2 % (Table S2), close to the family delineating threshold (60.0 %) of the order Rhodospirillales [15], supporting the proposal of a novel family. ANI and POCP values were less than 71.8 and 48.0 %, respectively (Table S2).
Comparative genomics
For genomic comparison analyses, a total of 447 genomes belonging to the order Rhodospirillales and 23 genomes of type strains belong to the order Rickettsiales in class Alphaproteobacteria were downloaded from the NCBI database. These genomes were predicted by Prodigal (version 2.6.3) [40] and annotated by using KofamKOALA with default parameters [41].
Strain Y-M2^T^ contains the nif family genes nifH, nifD, and nifK, which are key genes related to nitrogen fixation. nifH encodes Fe protein, which is a homodimer bridged by an inter-subunit (4Fe-4S) cluster that serves as the obligate electron donor for the MoFe protein, nifDK encodes Mo-nitrogenase, which is composed of dinitrogenase (MoFe protein) and dinitrogenase reductase (Fe protein) [42]. nifH, nifD, and nifK act as marker genes for predicting the nitrogen fixation capabilities of microbes [4345]. Therefore, strain Y-M2^T^ was speculated to have the ability to fix atmospheric nitrogen. To elucidate the adaptive mechanisms of the nif gene of Y-M2^T^, a comparative genomic analysis of the chromosomal regions flanking the nif gene clusters contained within a contig. The nif cluster genes nifH, nifD, and nifK were present in all N_2_-fixing strains in the order Rhodospirillales, whereas partial genes of the anf and vnf clusters were only found in M. fulvum DSM 113^T^, M. molischianum DSM120^T^, Rhodospirillum rubrum ATCC 11170^T^, and T. siberiense 26-4b^T^. Further analysis revealed that the anfG gene mediates crosstalk between the gene nifH and nifK encoding nitrogenases components [46], for which the assignments were similar to the archaea Methanosarcina activorans C2A^T^ (Fig. 3).
Maximum-likelihood tree reconstructed by concatenation of nifHDK genes. This tree includes the genomes of 45 nitrogen-fixing Rhodospirillales species and eight archaea nitrogen-fixing species from a variety of environments. Nodes with bootstrap values are marked with different size black dots. The colours of the labels and clades indicate the habitat where the strain was isolated and different lineages of nifHDK concatenation, respectively.
The maximum-likelihood tree was reconstructed by concatenation sequence of nifHDK genes, and the type of nifHKD was distincted to four distinct lineages (Fig. 3). Lineages I, II and III were most related to the genera Azospirillum, Magnetospirillum, and Roseospirillum. Interestingly, strain Y-M2^T^, as well as several species of the family Thalassospiraceae and Rhodospirillaceae, were placed in the lineage IV, which also contains archaea belonging to the genera Methanosarcina, Methanocella, Methanocaldococcus, Methanococcus, and Methanothermobacter, implying the homology of their nitrogenases (Fig. 3). In addition, most of bacteria and archaea in lineage IV were isolated from anaerobic or methanogenic environments (Fig. 3). A previous study has revealed that the distribution of nitrogenase genes was crucially impacted by horizontal gene transfer (HGT) events, the acquisition of nif by Firmicutes possibly through HGT events with a methanogen ancestor [47]. Therefore, the phylogenetic relationships among strain Y-M2^T^, Thalassospiraceae, Rhodospirillaceae species, and archaea in the nifHDK phylogenetic tree suggested the HGT events possibly occurred between Rhodospirillales and methanogens in anoxic environments (Fig. 3).
To further analyse nifHDK lineage IV, we reconstructed the nif gene organization based on encoding genes sequences. The nif clusters in the lineage IV could be classified to three distinct types (Fig. S2). The type I nif gene cluster was the most common nitrogenase in bacteria. Compared to type I, type II had an additional anfG gene inserted between nifH and nifK, whereas type III had genes nifN and nifE which replaced nifT in type I (Fig. S2). Surprisingly, the organization of nif genes showed that * T. siberiense* 26-4b1^T^ contained all types of nif cluster described above, which indicates that it may play an important role in the evolution of the nif genes in the order Rhodospirillales.
Morphology, physiology and chemotaxonomy
For morphology tests, the strain Y-M2^T^ was incubated at 25 °C in R2A medium for 7 days. Cell size, shape, and flagella were assessed using a scanning electron microscope (jeol JEM-1400 Plus) and a transmission electron microscope (jeol JEM-1230). Gram-stain and spore-stain kits (Solarbio) were used to assess Gram staining and spore formation, respectively. Growth of strain Y-M2^T^ at different temperatures (15, 20, 25, 30, and 37 °C), pH values (pH 5.5–8.0 at intervals of 0.5 pH units), and NaCl concentrations (0–70 g l^−1^ at intervals of 10 g l^−1^ NaCl) was determined in R2A broth by measuring the OD_600_ value with a spectrometer (DU730, Beckman Coulter). The following sterilized anoxic buffers at a final concentration of 20 mM were added into the fresh medium: MES (pH 5.5, 6.0, 6.5), PIPES (pH 7.0, 7.5), Tris (pH 8.0). The pH was finally adjusted by addition of solutions of HCl or NaOH before incubation, which was determined with a pH meter (Horiba). Oxygen requirement was tested under anaerobic, 2 %, 10% and 20 % of O_2_ (v/v) following the procedure described previously [48]. Biochemical characteristics of strain Y-M2^T^ were determined using API 20NE, API ZYM and API 20E kits (bioMérieux) according to the manufacturer’s instructions. Microbial nitrogenase (NITS) ELISA scientific research kit (Jingmei) was used to determine the content of nitrogenase. All experiments mentioned above were performed in triplicates. Cultures of strain Y-M2^T^ and T. siberiense 26-4b^T^ incubated at 25 °C in R2A medium were harvested at logarithmic phase for fatty acids analyses. The protocols of Microbial Identification Inc. (midi) and gas chromatography (Agilent 6990) were used to identify fatty acids using Sherlock software (version 6.3) [49]. Respiratory quinones were detected in strain Y-M2^T^ using a previously described protocol [5051]. Polar lipids were extracted using a chloroform–methanol system and were analysed using one- and two-dimensional thin-layer chromatography following the method described by Kates et al. [52].
Cells of strain Y-M2^T^ were Gram-negative, spiral-shaped with motility with monotrichous flagella, 0.8–3.0 µm long, 0.2–0.4 µm wide (Fig. S3), and non-spore-forming. Growth was observed at 20–30 °C, pH 6.0–7.5, without NaCl, and at 2–10 % O_2_. Optimal growth occurred at 25 °C, pH 7.0, without NaCl, and in the presence of 10 % O_2_.
Positive for acetoin (acetyl methylcarbinol) produced by glucose fermentation via the butylene glycol metabolic pathway and the production of the enzyme gelatinase which liquefies gelatin in API 20E test. Positive for arginine dihydrolase, urease, hydrolysis aesculin and β-galactosidase, negative for reduction of nitrates to nitrites or nitrogen in API 20NE tests (Table 1). In API ZYM tests, strain Y-M2^T^ presented positive reactions for alkaline phosphatase, esterase (C4), lipid esterase (C8), leucine arylaminase, acid phosphatase, naphthol AS-BI phosphate hydrolase, β-galactosidase, and β-glucosidase, and negative reactions for lipoid enzyme (C14), valine arylaminase, cystine arylaminase, trypsin, chymotrypsin, α-galactosidase, uronic acid glycosidase, α-glucosidase, N-acetylglucosamine enzyme, α-mannosidase, nd β-fucosidase. In addition, 335.7 ng l^−1^ nitrogenase was detected in about 0.1 ml of biomass of strain Y-M2^T^ (Fig. S4), indicating nitrogenase is positive in strain Y-M2^T^. Strain Y-M2^T^ contained Q-9 and Q-10 as the predominant quinones (14.2 and 85.2 %, respectively). The major fatty acids were iso C_17 : 0_ 3-OH (26.1 %), summed feature 8 (15.3 %), summed feature 3 (13.3 %), and iso-C_15 : 0_ 3-OH (12.6 %), which were different from the closest related strain containing C_18 : 1_* ω7c*, C_16 : 0_, and C_17 : 0_ as the major fatty acids (Table 1). The polar lipid profile comprised phosphatidylethanolamine, aminolipid, and aminophospholipidase as the main polar lipids (Fig. S5).
Comparative phenotypic characteristics between strain Y-M2^T^ and the closely related type strain T. siberiense 26-4b^T^ are presented in Table 1. The representative species of the order Rhodospirillales are mesophilic, and contain Q-9 and/or Q-10 as the predominant quinones, with the exception of Rhodovibrio salinarum DSM 915^T^, which contains MK-10. However, there were many differences that distinguish strain Y-M2^T^ from its closely related species T. siberiense 26-4b^T^, as well as the other families within Rhodospirillales, such as gelatinase activity and pyruvate utilization (Table 1). The cellular fatty acid compositions of Y-M2^T^ and T. siberiense 26-4b^T^ were different (Table 1). Furthermore, a physiological comparison revealed that strain Y-M2^T^ had differences in terms of sulphur and nitrate metabolism from the representative species of the families ‘Magnetospirillaceae’, ‘Novispirillaceae’, and Rhodospirillaceae (Table 2). Meanwhile, strain Y-M2^T^ could be distinguished in terms of urease from ‘Magnetospirillaceae’, and by cell shape from most representative species in Rhodospirillaceae. Therefore, on the basis of phylogenetic analyses, lower genomic indices, and physiological traits, we propose that strain Y-M2^T^ represents a novel family in the order Rhodospirillales.
In conclusion, strain Y-M2^T^ belongs to the order Rhodospirillales and represents a novel genus in a novel family based on the results of phylogenetic analyses, genome relatedness, as well as the observed differences in physiological traits, for which the name Oleispirillum naphthae gen. nov. sp. nov. within Oleispirillaceae fam. nov. is proposed. The type strain is Y-M2^T^.
Description of Oleispirillum gen. nov.
Oleispirillum (O.le.i.spi.ril’lum. L. neut. n. oleum oil; N.L. neut. dim. n. spirillum, a small spiral; N.L. neut. n. Oleispirillum small spiral associated with oil).
Gram-stain-negative, spiral, microaerophilic, mesophilic, motile, and non-spore-forming. The predominant cellular fatty acids are summed feature 3, iso-C_15 : 0_ 3-OH, summed feature 8, and iso-C_17 : 0_ 3-OH. Polar lipids include phosphatidylethanolamine, aminolipid and aminophospholipid. The predominant quinones are Q-9 and Q-10. The type species is Oleispirillaceae naphthae.
Description of Oleispirillum naphthae sp. nov
Oleispirillaceae naphthae (naph’thae. L. gen. fem. n. naphthae, of oil).
Cells are Gram-stain-negative, spiral, motile with flagella, and non-spore-forming. Cell size is 0.8–3.0×0.2–0.4 µm. Growth is observed at 20–30 °C (optimal temperature 25 °C), pH 7.0 (optimal pH 6.0–7.5) without NaCl under 2%–10 % oxygen. Positive reactions for alkaline phosphatase, esterase (C4), lipid esterase (C8), leucine arylaminase, acid phosphatase, naphthol AS-BI-phosphate hydrolase, β-galactosidase, and β-glucosidase. Negative reactions for lipoid enzyme (C14), valine arylaminase, cystine arylaminase, trypsin, chymotrypsin, α-galactosidase, uronic acid glycosidase, α-glucosidase, N-acetylglucosamine enzyme, α-mannosidase, and β-fucosidase in API ZYM tests. Positive for acetoin (acetyl methylcarbinol) produced by fermentation of glucose by bacteria utilizing the butylene glycol pathway and the production of the enzyme gelatinase which liquefies gelatin in API 20E tests. Positive for arginine dihydrolase, urease, hydrolysis aesculin and β-galactosidase, negative for reduction of nitrates to nitrites or nitrogen in API 20NE tests. The predominant quinones are Q-9 and Q-10. The predominant cellular fatty acids are summed feature 3, iso-C_15 : 0_ 3-OH, summed feature 8 and iso-C_17 : 0_ 3-OH. Polar lipids mainly include phosphatidylethanolamine, aminolipid, and aminophospholipid.
The type strain, Y-M2^T^ (=CCAM 827^T^=JCM 34765^T^), was isolated from oil sludge. The G+C content of the genomic DNA of strain Y-M2^T^ is 68.1 mol%.
The almost complete 16S rRNA gene sequence and draft genome of strain Y-M2^T^ was deposited to GenBank under the accession MZ270535 and JAQAZG000000000, respectively.
Description of Oleispirillaceae fam. nov
Oleispirillaceae (O.le.i.spi.ril.la.ce’ae. N.L. neut. n. Oleispirillum a bacterial genus; -aceae suffix to denote a family; N.L. fem. pl. n. Oleispirillaceae the Oleispirillum family).
The description of the family is based on the type genus Oleispirillum. This family is affiliated with the order Rhodospirillales in the class Alphaproteobacteria.
The type and only genus is Oleispirillum.
supplementary material
10.1099/ijsem.0.006292Uncited Supplementary Material 1.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Cordeiro AB Ribeiro RA Helene LCF Hungria M Rhizobium esperanzae sp. nov., a N 2-fixing root symbiont of Phaseolus vulgaris from Mexican soils Int J Syst Evol Microbiol 2017673937394510.1099/ijsem.0.00222528895521 · doi ↗ · pubmed ↗
- 2Divyasree B Lakshmi K Bharti D Sasikala C Ramana CV Caenispirillum deserti sp. nov., a spheroplast-forming bacterium isolated from a salt desert Int J Syst Evol Microbiol 2015653119312410.1099/ijsem.0.00039026296943 · doi ↗ · pubmed ↗
- 3Yoo S-H Weon H-Y Noh H-J Hong S-B Lee C-M et al Roseomonas aerilata sp. nov., isolated from an air sample Int J Syst Evol Microbiol 2008581482148510.1099/ijs.0.65385-018523199 · doi ↗ · pubmed ↗
- 4Yang Y Zhang R Feng J Wang C Chen J Azospirillum griseum sp. nov., isolated from lakewater Int J Syst Evol Microbiol 2019693676368110.1099/ijsem.0.00346031135333 · doi ↗ · pubmed ↗
- 5Baik KS Hwang YM Choi J-S Kwon J Seong CN Dongia rigui sp. nov., isolated from freshwater of a large wetland in Korea Antonie van Leeuwenhoek 20131041143115010.1007/s 10482-013-0036-924046206 · doi ↗ · pubmed ↗
- 6Zhou S Han L Wang Y Yang G Zhuang L et al Azospirillum humicireducens sp. nov., a nitrogen-fixing bacterium isolated from a microbial fuel cell Int J Syst Evol Microbiol 2013632618262410.1099/ijs.0.046813-023264502 · doi ↗ · pubmed ↗
- 7Young CC Hupfer H Siering C Ho M-J Arun AB et al Azospirillum rugosum sp. nov., isolated from oil-contaminated soil Int J Syst Evol Microbiol 20085895996310.1099/ijs.0.65065-018398202 · doi ↗ · pubmed ↗
- 8Wu D Zhang X-J Liu H-C Zhou Y-G Wu X-L et al Azospirillum oleiclasticum sp. nov, a nitrogen-fixing and heavy oil degrading bacterium isolated from an oil production mixture of Yumen oilfield Syst Appl Microbiol 20214412617110.1016/j.syapm.2020.12617133360414 · doi ↗ · pubmed ↗