The endogenous Mtv8 locus and the immunoglobulin repertoire
Helen A. Beilinson, Steven A. Erickson, Tatyana Golovkina

TL;DR
This study investigates whether a specific endogenous retrovirus, Mtv8, affects the diversity of immune receptors in mice.
Contribution
The study experimentally demonstrates that Mtv8 does not influence the B cell receptor repertoire in mice.
Findings
Mtv8-deficient mice were generated to test the role of Mtv8 in BCR repertoire formation.
The absence of Mtv8 does not alter the BCR repertoire in mice.
ERV-driven transcription does not affect V(D)J recombination in this context.
Abstract
The vast diversity of mammalian adaptive antigen receptors allows for robust and efficient immune responses against a wide number of pathogens. The antigen receptor repertoire is built during the recombination of B and T cell receptor (BCR, TCR) loci and hypermutation of BCR loci. V(D)J recombination rearranges these antigen receptor loci, which are organized as an array of separate V, (D), and J gene segments. Transcription activation at the recombining locus leads to changes in the local three-dimensional architecture, which subsequently contributes to which gene segments are utilized for recombination. The endogenous retrovirus (ERV) mouse mammary tumor provirus 8 (Mtv8) resides on mouse chromosome 6 interposed within the large array of light chain kappa V gene segments. As ERVs contribute to changes in genomic architecture by driving high levels of transcription of neighboring…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
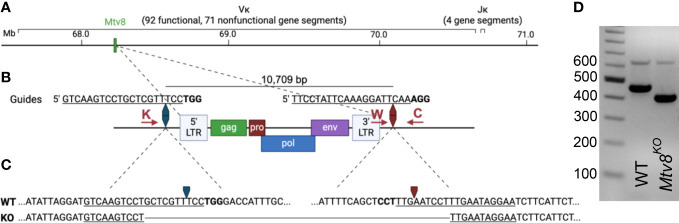 Figure 1
Figure 1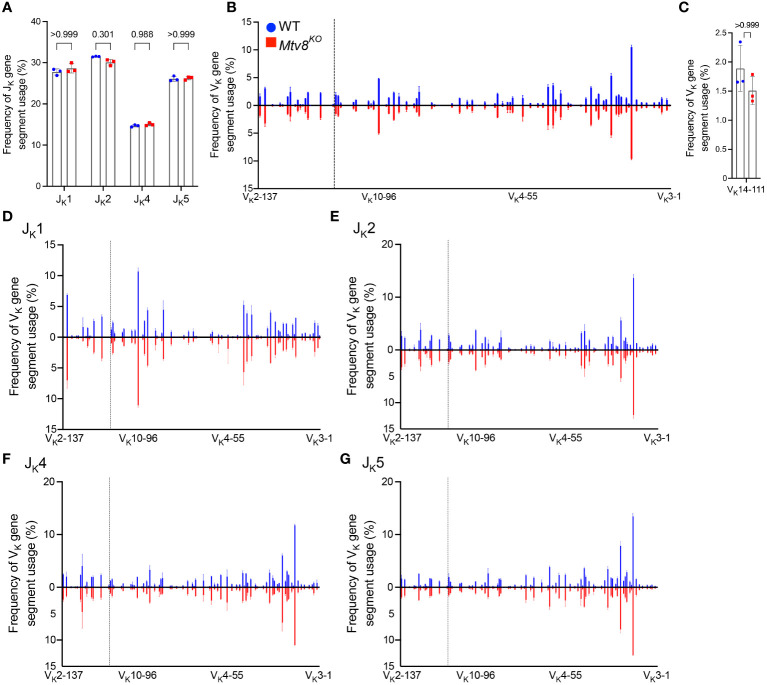 Figure 2
Figure 2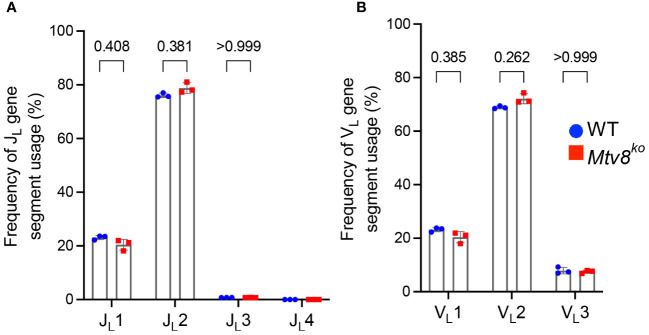 Figure 3
Figure 3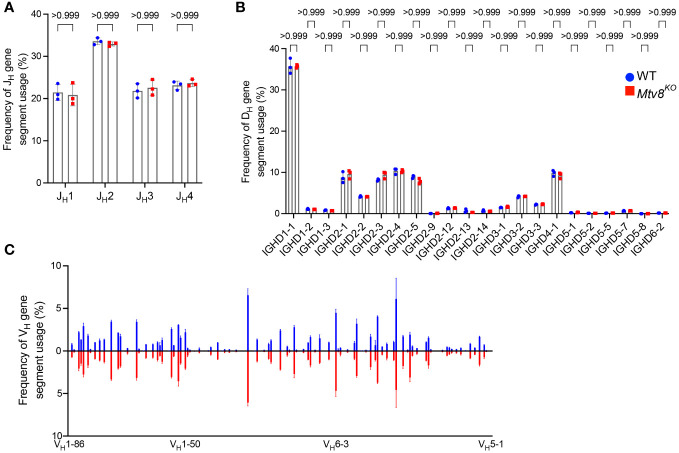 Figure 4
Figure 4Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsT-cell and B-cell Immunology · Immune Cell Function and Interaction · Immunotherapy and Immune Responses
Introduction
The extensive diversity of the jawed vertebrate adaptive immune response depends on the programmed assembly and hypermutation of antigen receptor (AgR) genes (1). The first stage of AgR assembly is V(D)J recombination, initiated by lymphocyte-specific recombination activating genes 1 and 2 (RAG1 and RAG2), during which immunoglobulin (Ig) and T cell receptor (TCR) genes are recombined from discrete variable (V), diversity (D), and joining (J) gene segments (2).
AgR assembly is a sequential process during lymphocyte development. In B cells, the Ig heavy-chain (Igh) locus recombines in early and pro B cells prior to the kappa and lambda light chain (Igk, Igl) loci in pre B cells (2, 3). Igh recombines in two phases: first, D_H_-to-J_H_ rearrangements occur in lymphoid progenitors; second, V_H_-to-DJ_H_ rearrangements occur in pro-B cells (2, 4). After successful (i.e. in-frame without premature stop codons) Igh recombination, Igk undergoes V_K_-to-J_K_ recombination (3). If neither Igk allele successfully rearranges, the Igl locus recombines in a V_L_-to-J_L_ fashion. During V(D)J recombination, RAG (a heterotetramer composed of RAG1 and RAG2 molecules) accumulates at recombination centers (RCs) that encompass J or DJ gene segments (for D-to-J/V-to-J and V-to-DJ recombination, respectively) (4). In RCs, RAG binds to a recombination signal sequences (RSS) that flanks the rearranging gene segment (4). Then, a partner RSS of the second gene segment is brought into the RC for synapsis and RAG-mediated cleavage.
V(D)J recombination determines the V, (D), and J gene segments used in a particular AgR gene and is dependent on three-dimensional chromosomal architecture. Specifically, V(D)J recombination is constrained to AgR loci by chromatin loops, the bases of which are defined by CCCTC-binding factor (CTCF)-bound CTCF-Binding Elements (CBEs) (5–9). The number of loops formed is dependent on the recombining locus (10). In addition to chromatin loops defining the AgR, chromosomal architecture also defines the mechanism by which recombining gene segments are brought into the RC (11).
The Igk locus is comprised of 92 functional (163 total) V_K_ and four functional J_K_ spread across 3.2 Mb (Figure 1A) and contracts into a recombination-competent chromosomal structure in developing pro- and pre-B cells (12, 13). Once contracted, the Igk locus forms a rosette-like structure with five V_K_-containing loops and one loop with J_K_ and C_K_ gene segments (10). Igk rearrangement occurs predominantly through the collision of, and subsequent recombination between, the J_K_-containing and a V_K_-containing loop (10). While the Igk loops are thought to be predominantly shaped by the CBEs throughout the locus, it is unknown whether other factors are involved.
Generation of Mtv8 KO mice. (A) Schematic of B6J murine Igk locus on chromosome 6. Location of Mtv8 is highlighted in green. (B) Schematic of Mtv8 and guides used for CRISPR/Cas9 targeting of Mtv8 to generate Mtv8KO mice. LTR, long-terminal repeat; gag, group-specific antigen; pro, protease; pol, polymerase; env, envelope. Red arrows: genotyping primers (K: KOF, W: WTF, C: CommonR). (C) Genomic sequence of WT and KO Mtv8 alleles. Only the flanking sequnces are shown for the WT allele. For (B, C), underlined text indicates guide sequence, bold text indicates PAM sequence, arrows indicate Cas9-cut site. (D) Gel image of genotyping representative WT and homozygous Mtv8KO mice. Mice were genotyped using a three-primer PCR with the KOF, WTF, and CommonR primers. WT product: 479 bp, KO product: 419 bp.
The endogenous retrovirus (ERV) mouse mammary tumor virus (MMTV), Mtv8 was mapped to the Igk locus on chromosome 6 (Figure 1A) (14–16). Mtv8 is a provirus with all open-reading frames (ORFs) intact, namely, gag, pro, pol, env, and superantigen (SAg) (Figure 1B). The provirus does not produce infectious virions and is silent in the mammary glands, the targeted tissue of all MMTVs, potentially due to hypermethylation of its promoter region (17–19). The Mtv8 SAg has a specificity for Vβ11 and Vβ12 (20). Mice strains harboring Mtv8 lack Vβ11^+^ and Vβ12^+^ T cells (21), indicating that Mtv8 is expressed in antigen presenting cells, such as dendritic cells and B cells which present SAg to cognate T cells (22). As Mtv8 is mapped within the Igk locus, it was hypothesized that it may contribute to Igk recombination by driving high levels of transcription in its vicinity (23). It was suggested that recombination to the first V_K_ gene segment downstream of Mtv8 (V_K_14-111, formerly V_K_9M) could allow for enhancer activation of the Mtv8 promoter (23). To identify whether Mtv8 affects the Igk repertoire, the frequency of recombination of the J_K_ gene segments to V_K_14-111 was analyzed in inbred mouse strains with [BALB/c, C58.C, A/J, and C57BL/6J (B6J)] and without (C58, C.C58, NZB, and PERA/Ei) Mtv8 (23). These experiments demonstrated that mice inheriting Mtv8 have higher recombination between V_K_14-111 and all J_K_ gene segments compared to strains without Mtv8. While these analyses suggested differences in the usage of V_K_14-111, Ig repertoire comparisons between inbred mouse lines is not an ideal approach, as polymorphisms within the loci other than Mtv8 could also influence the repertoires.
To address whether Mtv8 shapes the repertoire of mice with this ERV, we used CRISPR/Cas9 technology to generate B6J mice lacking Mtv8 and compared their Ig repertoire to that of wild-type (WT) B6J mice. We found that the absence of Mtv8 had no significant effects on the Igh, Igk, and Igl gene segment usage. Thus, contribution of Mtv8 to the mouse Ig repertoires can be definitively ruled out.
Methods
Mice
Mice utilized in this study were bred and maintained at the animal facility of The University of Chicago. The studies described herein have been reviewed and approved by the Animal Care and Use Committee at the University of Chicago, which is accredited by the Association for Assessment and Accreditation of Laboratory Animal Care (AAALAC International).
C57BL/6J (B6J) mice were purchased from The Jackson Laboratory. *Mtv8^KO^
- B6J mice were generated using CRISPR/Cas9 technology. Two guide RNAs targeting the flanking up- and downstream sequences of Mtv8 (5’ guide: 5’-GTCAAGTCCTGCTCGTTTCC; 3’ guide: 5’- TTCCTATTCAAAGGATTCAA) were co-injected into a single cell B6J embryos along with Cas9 (Figure 1B). Founder mice and subsequent offspring in which Mtv8 was eliminated were identified using PCR with primers flanking the guide cut sites (KOF: 5’-GAATTTGGGTGCTCTTGCAT; CommonR: 5’-AACACAAATGGAGGCAAAGC; KO product size: 419 bp). To identify mice with the WT allele, a separate PCR was used using the CommonR primer and a WT-specific reverse primer that lies in the excited region (WTF: 5’- CAGTCCTAACATTCACCTCT; WT product size: 479 bp). A founder line was established in which Mtv8 was eliminated with a 10,688 bp deletion (Figure 1C). The founder mouse was bred to a WT B6J mouse and the resulting F1 offspring were intercrossed to generate a homozygous KO line (Figure 1D). The deletion was confirmed at the DNA level by sequencing the KO allele using the KO-specific PCR primers.
RNA isolation from splenic B cells
CD19^+^ splenocytes were isolated from three WT and three *Mtv8^KO^
- 10.5-week-old B6J female mice. Red blood cell lysed splenocytes were labeled with microbeads conjugated to monoclonal anti-mouse CD19 antibodies (Miltenyi Biotec, Bergisch Gladbach, Germany) and positively sorted as detailed by the manufacturer. RNA was isolated from sorted cells using guanidine thiocyanate extraction and CsCl gradient centrifugation (24).
Library preparation
Immunoglobulin libraries were generated from 1 μg of RNA using the NEBNext Immune Sequencing Kit (New England Biolabs, Ipswich, Massachusetts, USA) according to manufacturer’s instructions, specifically enriching for B cell receptor (BCR) chains during the first PCR step using the mouse NEBNext IS BCR primers included in the kit (Mus-IgGb: 5’-GATGGGGCTGTTGTTKTRGC; Mus-IgGa: 5’-GTGTCGTTTTGGCCTG; Mus-IgE: 5’-GGTTCCTGATAGAGGC; Mus-IgD: 5’-GTTCCTTTTTATCACC; Mus-IgM: 5’-TGACTCTCCTGMRGARAC; Mus-IgA: 5’-GTGGGTAGATGGTGGG; Mus-IgK: 5’-RCATCAGCCMGGTWT; Mus-IgL: 5’-ATGGHGWRGMCTTGGG). The libraries generated from the six individual mice were pooled in equimolar amounts and sequenced by paired-end 300 bp sequencing on an Illumina MiSeq by The University of Chicago Genomics Facility.
B cell receptor sequence processing and analysis
Preprocessing of BCR sequences was performed using the open-source workflow pRESTO NEBNext Immune Sequencing Kit Workflow (v3.2.0) on Galaxy (25). Reads with a Phred quality score <20 were removed for quality control. Reads that did not match to the constant region primer (maximum error rate 0.2) were removed. Reads that did not match to the template switch sequence (maximum error rate 0.5) were removed. The first 17 bp following the template switch site were a unique molecular identifier (UMI) on each read. Sequences with identical UMIs were collapsed into consensus sequences with sequences found in less than 60% of reads removed. Positions with more than 50% gap sequences were removed. Mate-pairs were assembled with a minimum of 8 bp overlap (maximum error rate of 0.3). Assembled reads were assigned isotype-constant region identities based on local alignment of the 3’ ends of the reads (maximum error rate of 0.3). Using the Mus musculus reference C57BL/6J genome (GRCm38/mm10), V, (D), and J gene segments were assigned using MiGMAP mapper (Galaxy Version 1.0.3+galaxy2). Subsequent analyses were done using R Studio. Statistically significance was calculated using unpaired Welch’s t test with Bonferroni correction. Graphing was done using GraphPad Prism version 10.0.1 for Mac (GraphPad Software, Boston, Massachusetts, USA).
Results
Generation of Mtv8-deficient B6J mice
Mtv8 knockout (KO*)* B6J mice were generated using a CRISPR/Cas9 approach. To target Mtv8 without disturbing the V_K_ gene segments in its vicinity, we designed two guides to precisely delete the ERV: one 645 bp upstream of the 5’ LTR and one 79 bp downstream of the 3’ LTR (Figure 1B). A founder was identified using PCR with primers flanking the predicted deleted region and it was determined that a 10,688 bp deletion occurred, resulting in the elimination of the entire Mtv8 locus (Figures 1C, D). The founder was crossed to a wildtype (WT) B6J mouse and heterozygous, mutant allele-carrying progeny were interbred to generate a homozygous Mtv8-deficient line (Figure 1D).
Mtv8 does not contribute to light and heavy chain recombination
To investigate whether Mtv8 alters Ig gene segment usage, we analyzed the Ig repertoires of WT and *Mtv8^KO^
- B6J mice (Figures 2-4, Supplementary Table 1). Accordingly, RNA isolated from CD19^+^ splenocytes was used to prepare Ig heavy and light chain-specific 5’-RACE libraries, which were analyzed to determine the BCR repertoire using the pRESTO toolkit using the Mus musculus reference C57BL/6J genome (GRCm38/mm10) (25). The repertoire of productive transcripts was analyzed. Productive transcripts were defined as in-frame without premature stop codons and are likely those transcripts that are translated into expressed Ig proteins.
Igk repertoire of CD19+ splenocytes of Mtv8KO mice. Frequency of JK (A) and VK (B) gene segments in productive Igk transcripts from CD19+ splenocytes. Productive recombination events are in-frame without premature stop codons. (C) Frequency of VK14-111 (previously termed VK9M) in and productive Igk transcripts. (D-G) Frequency of VK gene segments in total Igk transcripts recombined to JK1 (D), JK2 (E), JK4 (F), and JK5 (G). (B, D-G) VK gene segments are arranged from 5’ distal to 3’ proximal to JK gene segments and the vertical dotted line represents chromosomal location of Mtv8. n=3 mice per group. Data presented as mean with error bars indicated SD. Statistical significance was determined by unpaired Welch’s t test with Bonferroni correction (numbers above bars indicate adjusted P value).
To determine whether Mtv8 influences Igk recombination, we examined the J_K_ and V_K_ usage in productive Igk transcripts from CD19^+^ splenocytes (Figure 2). There was no difference in J_K_ usage (Figure 2A). Furthermore, the loss of Mtv8 led to no alterations in the total V_K_ repertoire (Figure 2B). Notably, there was no change in usage of V_K_ gene segments mapped in the close proximity of Mtv8 (dotted line; Figure 2B). An increased usage of the V_K_ gene segment, V_K_14-111 (formerly called V_K_9M) directly downstream of Mtv8 in *Mtv8^+^
- mouse strains compared to *Mtv8^-^
- strains was previously reported (23). We observed no difference in frequency of V_K_14-111 usage in Igk transcripts between WT and *Mtv8^KO^
- B6J mice (Figure 2C). Thus, the previously observed disparities in V_K_14-111 usage between *Mtv8^+^
- and *Mtv8^-^
- mice from distinct genetic backgrounds are independent of Mtv8.
The same Igk locus can undergo multiple rounds of recombination. While Mtv8 does not affect the overall frequency of V_K_ usage, we wanted to test the possibility that it might affect either the first or subsequent recombination in distinct ways. Recombination of any V_K_ to J_K_1 can only occur during a primary recombination event, as recombination to any other J_K_ gene segment would remove J_K_1 from the Igk locus. Recombination to J_K_2, J_K_4, or J_K_5 can either occur during a primary or secondary recombination event. As such, we calculated the frequency of V_K_ gene segment usage in total recombination events with each J_K_ gene segment. We found that, in line with globally unaffected J_K_ gene segment usage, frequency of any particular V_K_ gene segment recombining to J_K_1, J_K_2, J_K_4, and J_K_5 was unchanged with the loss of Mtv8 (Figures 2D–G). Taken together, these data show that the Igk repertoire of *Mtv8^KO^
- B6J mice does not differ from WT B6J and that Mtv8 does not influence the Igk recombination.
We also found no difference in the J_L_ and V_L_ usage among productive Igl transcripts between WT and *Mtv8^KO^
- B6J mice (Figure 3). Similarly, we found no alterations in J_H_, D_H_, and V_H_ gene segment usage between WT and *Mtv8^K^
^O^ mice (Figure 4).
Igl repertoire of CD19+ splenocytes of Mtv8KO mice. Frequency of JL (A) and VL (B) gene segments in and productive Igl transcripts from CD19+ splenocytes. Productive recombination events are in-frame without premature stop codons. n=3 mice per group. Data presented as mean with error bars indicated SD. Statistical significance was determined by unpaired Welch’s t test with Bonferroni correction (numbers above bars indicate adjusted P value). No P value is indicated for JL4 as values for all samples are 0.
Igh repertoire of CD19+ splenocytes of Mtv8KO mice. Frequency of JH (A), DH (B), and VH (C) gene segments in productive Igh transcripts from CD19+ splenocytes. Productive recombination events are in-frame without premature stop codons. (B) DH gene segments are arranged from 5’ distal to 3’ proximal to JH gene segments. (C) Frequency of VH gene segments in productive Igh transcripts that are statistically different between WT and Mtv8 KO mice. VH gene segments are arranged from 5’ distal to 3’ proximal to DH gene segments. n=3 mice per group. Data presented as mean with error bars indicated SD. Statistical significance was determined by unpaired Welch’s t test with Bonferroni correction (numbers above bars indicate adjusted P value).
Discussion
It is now accepted that the collisions of the rosette-like loops of the Igk locus are the major mechanism of V_K_-to-J_K_ recombination (10). However, the identification of an ERV, Mtv8, in the middle of the V_K_ array on the Igk locus led to the hypothesis that it may influence the Ig repertoire (14–16). Differences in V_K_ usage in mouse strains with and without Mtv8 initially supported this hypothesis (23). Now, advancements in genome-editing technologies allow us to definitely address whether a genetic loss of Mtv8 can change the Ig repertoire within a mouse line. We show that Mtv8 has no effect on the Ig repertoire by analyzing the Igh, Igk, and Igl transcriptional repertoires of B cells isolated from WT and Mtv8 ^KO^ B6J mice. These data demonstrate that while Mtv8 is an intact ERV at the Igk locus, it has no influence on Igk recombination.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE248410, GSE248410.
Ethics statement
The animal study was approved by University of Chicago Animal Resources Center. The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
HB: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Writing – original draft, Writing – review & editing. SE: Methodology, Writing – review & editing. TG: Funding acquisition, Methodology, Project administration, Resources, Supervision, Writing – review & editing.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Cooper MD Alder MN. The evolution of adaptive immune systems. Cell. (2006) 124:815–22. doi: 10.1016/j.cell.2006.02.001 16497590 · doi ↗ · pubmed ↗
- 2Schatz DG Swanson PC. V(D)J recombination: mechanisms of initiation. Annu Rev Genet. (2011) 45:167–202. doi: 10.1146/annurev-genet-110410-132552 21854230 · doi ↗ · pubmed ↗
- 3Alt FW Zhang Y Meng FL Guo C Schwer B. Mechanisms of programmed DNA lesions and genomic instability in the immune system. Cell. (2013) 152:417–29. doi: 10.1016/j.cell.2013.01.007 PMC 438291123374339 · doi ↗ · pubmed ↗
- 4Schatz DG Ji Y. Recombination centres and the orchestration of V(D)J recombination. Nat Rev Immunol. (2011) 11:251–63. doi: 10.1038/nri 2941 21394103 · doi ↗ · pubmed ↗
- 5Jhunjhunwala Svan Zelm MC Peak MM Murre C. Chromatin architecture and the generation of antigen receptor diversity. Cell. (2009) 138:435–48. doi: 10.1016/j.cell.2009.07.016 PMC 272683319665968 · doi ↗ · pubmed ↗
- 6Zhang Y Zhang X Dai HQ Hu H Alt FW. The role of chromatin loop extrusion in antibody diversification. Nat Rev Immunol. (2022) 22:550–66. doi: 10.1038/s 41577-022-00679-3 PMC 937619835169260 · doi ↗ · pubmed ↗
- 7Davidson IF Bauer B Goetz D Tang W Wutz G Peters JM. DNA loop extrusion by human cohesin. Science. (2019) 366:1338–45. doi: 10.1126/science.aaz 3418 31753851 · doi ↗ · pubmed ↗
- 8Kim Y Shi Z Zhang H Finkelstein IJ Yu H. Human cohesin compacts DNA by loop extrusion. Science. (2019) 366:1345–9. doi: 10.1126/science.aaz 4475 PMC 738711831780627 · doi ↗ · pubmed ↗