Targeting bone marrow mechanosensation in myelofibrosis
Francesca Vinchi

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
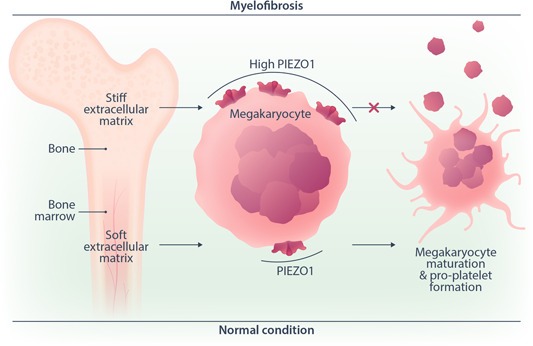 Figure 1
Figure 1- —None
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsErythrocyte Function and Pathophysiology · Myeloproliferative Neoplasms: Diagnosis and Treatment · Hemoglobinopathies and Related Disorders
Primary myelofibrosis (PMF) is a chronic myeloproliferative neoplasm characterized by progressive accumulation of fibrosis in the bone marrow.1 PMF results from the neoplastic transformation of a pluripotent hematopoietic stem cell. PMF progeny cells stimulate bone marrow fibroblasts—which are not part of the neoplastic transformation—to produce excessive collagen. As scar tissue increases, bone marrow failure eventually occurs, with consequent anemia and thrombocytopenia.1
Mutations of the Janus kinase 2 (JAK2) gene are present in a high proportion of cases of PMF. JAK2 is a member of the class I type tyrosine kinase family of enzymes and is involved in signal transduction initiated by erythropoietin, thrombopoietin, and granulocyte colony‐stimulating factor receptors among other stimuli. Mutations of the thrombopoietin receptor gene (MPL) or the calreticulin (CALR) gene also may be the cause of PMF.
PMF hallmarks are megakaryocytosis, with atypical immature megakaryocytes and the accumulation of cross‐linked collagen and reticulin fibers in the bone marrow, which gradually alter the architecture and stiffness of the extracellular matrix.1 With the progression of the disease, patients with PMF often present with thrombocytopenia and high‐grade fibrosis, which are indicative of a more advanced disease state.
The mechanical properties of the extracellular matrix within the bone marrow microenvironment exert a crucial role in various aspects of megakaryocyte development, including lineage commitment, polyploidization, and proplatelet formation. A softer environment likely supports these differentiation and maturation processes. Indeed, megakaryocytes cultured on soft substrates exhibit enhanced maturation and platelet production compared to stiffer surfaces.2, 3 However, how this sensing occurs and the membrane mechanosensor that mediates it and plays a role in megakaryocytes have remained unknown so far. A recent study published in the American Journal of Hematology unveiled the involvement of the mechanosensor PIEZO1 in the regulation of megakaryocyte maturation and proplatelet biogenesis under physiological and pathological conditions.
Mechanosensitive ion channels are integral membrane proteins that play a crucial role in fast signaling during mechanosensory transduction processes in living cells. As molecular transducers of mechanical force, mechanosensitive channels are activated by mechanical stimuli exerted on cellular membranes, upon which they rapidly and efficiently convert these stimuli into electrical, osmotic, and/or chemical intracellular signals. PIEZO1 is a cation channel that senses and transduces externally applied mechanical forces acting on the lipid plasma membrane and influences cell fate and tissue homeostasis in multiple cell types and tissues. PIEZO1 has a role in different blood cells, including red blood cells, macrophages, and platelets.4, 5, 6, 7 In erythrocytes, PIEZO1 controls cell differentiation and hydration status following mechanical activation4, 5; in macrophages, it modulates macrophage polarization and stiffness sensing6; in platelets, PIEZO1 activation by shear stress regulates calcium flux and thrombotic activity.7
Abbonante et al.8 recently proved a key role for PIEZO1 in megakaryopoiesis by taking advantage of mice deficient of the Piezo genes in the megakaryocyte lineage. Mice lacking PIEZO in megakaryocytes showed an increased number of circulating platelets both at steady state and upon myeloregeneration, suggesting a role for PIEZO activation in restraining platelet production. Indeed, the use of PIEZO1 pharmacological agonists on mouse and human cells increased immature megakaryocytes and decreased mature ones, reducing proplatelet formation via impaired megakaryocyte maturation.
Since PMF is associated with a severely modified extracellular matrix in the bone marrow, mechanical sensing is likely altered in this pathological condition. Interestingly, megakaryocytes from JAK2 ^V617F^ mice as well as megakaryocytes and platelets from PMF patients showed an increased expression of PIEZO1 at messenger RNA and protein levels. In particular, PIEZO1 expression was higher in platelets from patients showing overt compared to early PMF, being characteristic of an advanced fibrotic disease. Indeed, PIEZO expression increased when human megakaryocytes were grown in vitro into a stiff compared to a soft three‐dimensional matrix, indicating that, in PMF alterations of bone marrow, stiffness results in PIEZO1 overexpression (Figure 1). Finally, the negative correlation between megakaryocyte PIEZO1 expression and blood platelet count in patients with PMF confirmed a role for PIEZO1 as a negative regulator of megakaryocyte maturation and platelet production. Overall, these findings support a role for PIEZO1 overexpression in the pathological megakaryopoiesis and reduced platelet production observed in PMF. Interestingly, the use of PIEZO1 inhibitors promoted PMF megakaryocyte maturation and proplatelet formation in vitro. This underscores PIEZO1 inhibition as a potential therapeutic strategy to mitigate the impaired megakaryocyte development underlying PMF pathophysiology. Future studies will determine whether the increased PIEZO1 expression observed in platelets from patients with PMF might contribute to the increased thrombotic risk that is described in these patients.
Overall, this study highlights the involvement of the mechanosensor PIEZO1 in regulating megakaryocyte maturation and proplatelet biogenesis8 (Figure 1). Upregulated PIEZO1 in PMF likely contributes to the aberrant megakaryopoiesis associated with this disease and negatively impacts platelet biogenesis. The discovery that PIEZO1 acts as a brake of megakaryocyte differentiation and platelet production opens up the opportunity to develop targeted therapies for megakaryocytes and/or platelet disorders associated with altered bone marrow microenvironment and impaired mechanosensation.
AUTHOR CONTRIBUTIONS
Francesca Vinchi conceived and wrote the Hematopic.
CONFLICT OF INTEREST STATEMENT
Dr. Vinchi is a member of the advisory board of Silence Therapeutics and a consultant for RallyBio. None of these relationships is relevant to the current publication.
FUNDING
Dr. Vinchi receives research funding from CSL Vifor, Silence Therapeutics, and PharmaNutra, none of these relevant for this work.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Zahr AA , Salama ME , Carreau N , et al. Bone marrow fibrosis in myelofibrosis: pathogenesis, prognosis and targeted strategies. Haematologica. 2016;101(6):660‐671.27252511 10.3324/haematol.2015.141283 PMC 5013940 · doi ↗ · pubmed ↗
- 2Abbonante V , Di Buduo CA , Gruppi C , et al. A new path to platelet production through matrix sensing. Haematologica. 2017;102(7):1150‐1160.28411253 10.3324/haematol.2016.161562 PMC 5566016 · doi ↗ · pubmed ↗
- 3Di Buduo CA , Wray LS , Tozzi L , et al. Programmable 3D silk bone marrow niche for platelet generation ex vivo and modeling of megakaryopoiesis pathologies. Blood. 2015;125(14):2254‐2264.25575540 10.1182/blood-2014-08-595561 PMC 4383799 · doi ↗ · pubmed ↗
- 4Caulier A , Jankovsky N , Demont Y , et al. PIEZO 1 activation delays erythroid differentiation of normal and hereditary xerocytosis‐derived human progenitor cells. Haematologica. 2020;105(3):610‐622.31413092 10.3324/haematol.2019.218503 PMC 7049340 · doi ↗ · pubmed ↗
- 5Cahalan SM , Lukacs V , Ranade SS , Chien S , Bandell M , Patapoutian A . Piezo 1 links mechanical forces to red blood cell volume. e Life. 2015;4:4.10.7554/e Life.07370 PMC 445663926001274 · doi ↗ · pubmed ↗
- 6Atcha H , Jairaman A , Holt JR , et al. Mechanically activated ion channel Piezo 1 modulates macrophage polarization and stiffness sensing. Nat Commun. 2021;12(1):3256.34059671 10.1038/s 41467-021-23482-5PMC 8167181 · doi ↗ · pubmed ↗
- 7Zhao W , Wei Z , Xin G , et al. Piezo 1 initiates platelet hyperreactivity and accelerates thrombosis in hypertension. J Thromb Haemostasis. 2021;19(12):3113‐3125.34411418 10.1111/jth.15504 · doi ↗ · pubmed ↗
- 8Abbonante V , Karkempetzaki AI , Leon C , et al. Newly identified roles for PIEZO 1 mechanosensor in controlling normal megakaryocyte development and in primary myelofibrosis. Am J Hematol. 2024;99(3):336‐349. 10.1002/ajh.271841 38165047 PMC 10922533 · doi ↗ · pubmed ↗