The symphony of maize signaling quartet defending against gray leaf spot
Ping Wang, Ping He

TL;DR
The paper explains how a group of four proteins in maize work together to defend against a common fungal disease, showing how plant immunity is conserved.
Contribution
The study identifies a conserved maize signaling quartet that governs resistance to gray leaf spot, a major fungal disease.
Findings
A maize signaling module comprising ZmWAKL-ZmWIK-ZmBLK1-ZmRBOH4 governs resistance to gray leaf spot.
This signaling quartet is conserved in plant immunity across species.
The module activates NADPH oxidases to generate reactive oxygen species (ROS) for defense.
Abstract
In plant immunity, a well-orchestrated cascade is initiated by the dimerization of receptor-like kinases (RLKs), followed by the phosphorylation of receptor-like cytoplasmic kinases (RLCKs) and subsequent activation of NADPH oxidases for ROS generation. Recent findings by Zhong et al. illustrated that a maize signaling module comprising ZmWAKL-ZmWIK-ZmBLK1-ZmRBOH4 governs quantitative disease resistance to grey leaf spot, a pervasive fungal disease in maize worldwide, unveiling the conservation of this signaling quartet in plant immunity.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
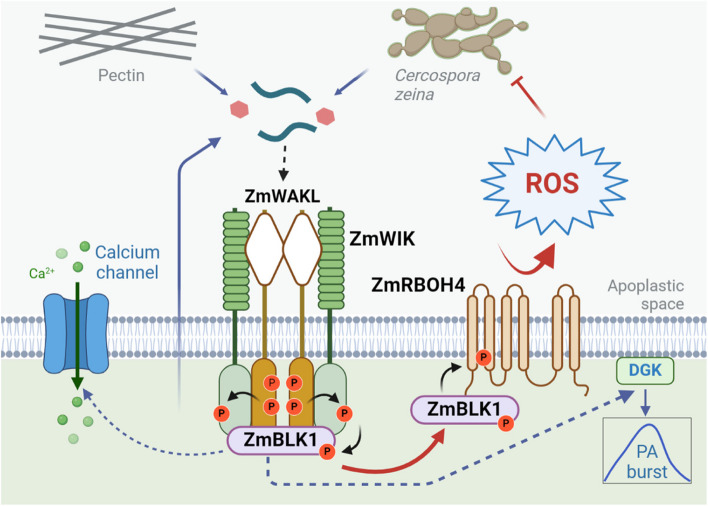 Figure 1
Figure 1- —http://dx.doi.org/10.13039/501100001809National Natural Science Foundation of China
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant-Microbe Interactions and Immunity · Plant Pathogenic Bacteria Studies · Plant pathogens and resistance mechanisms
Main text
Plant receptor-like kinases (RLKs) are important cell surface pattern recognition receptors (PRRs), recognizing a plethora of microbe- and danger-associated molecular patterns (MAMPs/DAMPs), ultimately orchestrating pattern-triggered immunity (PTI). Plant genomes contain hundreds of RLKs with diverse ectodomain (ECD) architectures, such as leucine-rich repeat (LRR), lysine motif (LysM), lectin motif, and malectin-like domain (Hou et al. 2021). Wall-associated kinases (WAKs) and WAK-like kinases (WAKLs) are a unique group of RLKs with extracellular galacturonan-binding (GUB) domains, which likely link to the cell wall (Stephens et al. 2022). WAKs and WAKLs emerge as formidable guardians against pathogens with diverse lifestyles (Stephens et al. 2022). For example, cotton GhWAK7A mediates resistance to Verticillium and Fusarium wilts by modulating the fungal chitin sensory complex (Wang et al. 2020).
A recent study in Nature Genetics by Zhong et al. illustrated a saga of maize WAKL-mediated defense against gray leaf spot (GLS), caused by Cercospora zeae-maydis and Cercospora zeina, a major fungal disease in maize worldwide (Zhong et al. 2024). Through the meticulous fine mapping of the major quantitative disease resistance (QDR) locus and comprehensive transgenic assays, ZmWAKL was found to be the major gene for conferring resistance against GLS, with ZmWAKL^Y^ as the resistant isoform and ZmWAKL^Q^ as the susceptible isoform. While the intracellular domain (ICD) of ZmWAKL^Y^ and ZmWAKL^Q^ remains highly conserved, their ECDs, particularly the GUB domain, diverge significantly. ECDs in RLKs play a crucial role in ligand binding. The data suggest that ZmWAKL^Y^ and ZmWAKL^Q^ likely have differences in perceiving MAMPs from pathogens or DAMPs from plants. This hypothesis was corroborated by an elegant chimeric gene experiment in which the fuse of ECD from ZmWAKL^Y^ and ICD from ZmWAKL^Q^ conferred the GLS resistance (Zhong et al. 2024). Similarly, the variation between resistant and susceptible isoforms of Rlm9, an oilseed WAKL involved in race-specific resistance against Leptosphaeri maculans, is concentrated in the ECD GUB domain (Larkan et al. 2020). In addition, unlike ZmWAKL^Q^, ZmWAKL^Y^ was self-associated via the GUB domain, suggesting the formation of homodimers of ZmWAKL^Y^. The ICD of ZmWAKL^Y^, not ZmWAKL^Q^, exhibited kinase activity both in vitro and in vivo, substantiating the importance of ZmWAKL^Y^ in transducing immune signaling (Zhong et al. 2024). Notably, the phosphorylation of ZmWAKL^Y^ was enhanced after C. zeina infection, supporting the physiological importance of ZmWAKL^Y^ phosphorylation.
By performing an immunoprecipitation-mass spectrometry assay, Zhong et al. further demonstrated that ZmWIK, an LRR-RLK, interacted with ZmWAKL and was important in conferring GLS resistance. Both the ECD and ICD of ZmWIK preferentially interacted with ZmWAKL^Y^ compared to ZmWAKL^Q^ (Zhong et al. 2024). Similar to other RLK complexes, the transphosphorylation was observed between ZmWIK and ZmWAKL. Apparently, ZmWIK was more strongly phosphorylated than ZmWAKL when these two proteins were mixed, suggesting a potential role for ZmWIK in amplifying ZmWAKL phosphorylation in the signal relay process. With RLK receptor and coreceptor dimerization and transphosphorylation serving as the cornerstone of plant PTI (Hou et al. 2021), the authors proposed that ZmWIK is a coreceptor of ZmWAKL. Yet, intriguing questions persist, including the regulation of ZmWIK-ZmWAKL dimerization in response to pathogen invasion and the potential divergence in ZmWIK sequences and expression between GLS-resistant and susceptible maize varieties.
Receptor-like cytoplasmic kinases (RLCKs) are convergent signaling regulators downstream of multiple PRRs in relaying multiple intracellular signaling events, including ROS burst, calcium influx, and MAPK activation (Lin et al. 2013; Liang and Zhou 2018). Similarly, maize RLCK ZmBLK1 was associated with both ZmWIK and ZmWAKL. Apparently, the association between ZmBLK1 and ZmWIK was stronger than the ZmBLK1-ZmWAKL association, supporting the role of ZmWIK being the coreceptor (Zhong et al. 2024). Moreover, ZmBLK1, ZmWAKL, and ZmWIK form a complex at the plasma membrane, underscoring their collaborative efforts in immune signaling. Furthermore, ZmWIK strongly phosphorylated ZmBLK1, and this phosphorylation was further increased when ZmWAKL^Y^ was present (Zhong et al. 2024). This aligns with the interaction and phosphorylation of Arabidopsis RLCK BIK1 by the PRR coreceptor BAK1 (Lin et al. 2014).
A key function of RLCKs in plant immunity is to regulate ROS generation by phosphorylating NADPH oxidases, specifically the respiratory burst oxidase homologs (RBOHs) (Liang and Zhou 2018). Zhong et al. found that ZmRBOH4, one of the six RBOHs in maize, was highly expressed and induced by pathogens in leaves. The ZmRBOH4 knocked-out lines exhibited compromised C. zeina-induced ROS levels and were susceptible to GLS, paralleling the vulnerability observed in Zmwik null mutants (Zhong et al. 2024). Biochemistry assays revealed that ZmBLK1 interacted and phosphorated the N-terminal region of ZmRBOH4. Moreover, the phosphorylation of ZmRBOH4 by ZmBLK1 was enhanced in the presence of ZmWAKL^Y^ and ZmWIK, supporting that ZmRBOH4 is a component of ZmWAKL^Y^-ZmWIK-ZmBLK1 complex orchestrating GLS resistance in maize (Zhong et al. 2024).
Collectively, in this groundbreaking discovery, the authors uncover a conserved signaling quartet that serves as the linchpin in maize's defense against GLS. The quad, comprising ZmWAKL, ZmWIK, ZmBLK1, and ZmRBOH4, emerges as a masterful conductor orchestrating maize's immune defense mechanism (Fig. 1). By drawing parallels with analogous modules in other plant species, this study illuminates the evolutionary conservation of pivotal signaling pathways across diverse plant lineages. A pivotal aspect of this discovery lies in the recognition of ZmWAKL as a quantitative disease-resistance gene, a feat often hindered by the challenges associated with dissecting and verifying quantitative traits genetically. Nevertheless, by unraveling the intricate workings of the ZmWAKL–ZmWIK–ZmBLK1–ZmRBOH4 module, this study not only enhances our understanding of maize immunity but also unveils promising avenues for engineering crops with enhanced disease resistance.Fig. 1. The proposed model of the ZmWAKL–ZmWIK–ZmBLK1–ZmRBOH4-mediated immunity. Upon C. zeina invasion, the ligand molecule from the pathogen, plant cell wall pectin, or secreted protein from cytosol is sensed by the ZmWAKL homodimer and induces ZmWAKL phosphorylation. ZmWIK functions as a coreceptor of ZmWAKL and is phosphorylated by ZmWAKL. ZmBLK1 is the RLCK that interacts with and is phosphorylated by both ZmWAKL and ZmWIK for signaling relay. The NADPH oxidase ZmRBOH4 is the substrate of ZmBLK1 and generates the ROS burst upon ZmBLK1 phosphorylation, culminating in GLS resistance. It is possible that ZmBLK1 also activates other substrates, such as calcium channels and diacylglycerol kinases (DGKs), for calcium influx and phosphatidic acid (PA) burst, conferring broad resistance against pathogens
WAKs/WAKLs play versatile roles during plant–microbe interactions, ranging from the detection of potential patterns from plants or microbes, modulation of PRR complexes, function of signaling components, and biosynthesis of cellulose and secondary metabolite (Stephens et al. 2022). As such, probing the precise role of ZmWAKL in perceiving either MAMPs from C. zeina or DAMPs released or induced in plants upon infection is an intriguing avenue for future research. Similarly, the possibility of ZmWIK serving as the bona fide PRR within the ZmWAKL–ZmWIK–ZmBLK1–ZmRBOH4 module cannot be discounted. Furthermore, in addition to their involvement in regulating ROS burst, Arabidopsis RLCKs have been shown to modulate cytosolic calcium influx and phosphatidic acid burst in plant immunity (Fig. 1) (Tian et al. 2019; Kong et al. 2024). Exploring whether the ZmWAKL–ZmWIK–ZmBLK1–ZmRBOH4 module extends its regulatory reach to other signaling events in plant immunity could provide valuable insights into its contribution to broad-spectrum resistance.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Hou S Liu D He P Phytocytokines function as immunological modulators of plant immunity Stress Biology 20211810.1007/s 44154-021-00009-y 34806087 PMC 8591736 · doi ↗ · pubmed ↗
- 2Kong L Ma X Zhang C Kim SI Li B Xie Y Yeo IC Thapa H Chen S Devarenne TP Munnik T He P Shan L Dual phosphorylation of DGK 5-mediated PA burst regulates ROS in plant immunity Cell 20241873609623.e 2110.1016/j.cell.2023.12.03038244548 PMC 10872252 · doi ↗ · pubmed ↗
- 3Larkan NJ Ma L Haddadi P Buchwaldt M Parkin IAP Djavaheri M Borhan MH The Brassica napus wall-associated kinase-like (WAKL) gene Rlm 9 provides race-specific blackleg resistance Plant J 202010489290010.1111/tpj.1496632794614 PMC 7756564 · doi ↗ · pubmed ↗
- 4Liang X Zhou JM Receptor-like cytoplasmic kinases: central players in plant receptor kinase-mediated signaling Annu Rev Plant Biol 20186926729910.1146/annurev-arplant-042817-04054029719165 · doi ↗ · pubmed ↗
- 5Lin W Ma X Shan L He P Big roles of small kinases: the complex functions of receptor-like cytoplasmic kinases in plant immunity and development J Integr Plant Biol 2013551188119710.1111/jipb.1207123710768 PMC 4391744 · doi ↗ · pubmed ↗
- 6Lin W Li B Lu D Chen S Zhu N He P Shan L Tyrosine phosphorylation of protein kinase complex BAK 1/BIK 1 mediates Arabidopsis innate immunity Proc Natl Acad Sci U S A 20141113632363710.1073/pnas.131881711124532660 PMC 3948311 · doi ↗ · pubmed ↗
- 7Stephens C Hammond-Kosack KE Kanyuka KWA Ksing plant immunity, waning diseases J Exp Bot 202273223710.1093/jxb/erab 42234520537 · doi ↗ · pubmed ↗
- 8Tian W Hou C Ren Z Wang C Zhao F Dahlbeck D Hu S Zhang L Niu Q Li L Staskawicz BJ Luan SA calmodulin-gated calcium channel links pathogen patterns to plant immunity Nature 201957213113510.1038/s 41586-019-1413-y 31316205 · doi ↗ · pubmed ↗