The Impact of Phenological Gaps on Leaf Characteristics and Foliage Dynamics of an Understory Dwarf Bamboo, Sasa kurilensis
Chongyang Wu, Ryota Tanaka, Kyohei Fujiyoshi, Yasuaki Akaji, Muneto Hirobe, Naoko Miki, Juan Li, Keiji Sakamoto, Jian Gao

TL;DR
This paper explores how phenological gaps in forest canopies affect the growth and leaf characteristics of the dwarf bamboo Sasa kurilensis.
Contribution
The study reveals how S. kurilensis adapts to phenological gaps through changes in leaf morphology and biomass.
Findings
Phenological gaps increase leaf area, thickness, and carbon content per unit area in S. kurilensis.
Phenological gaps do not significantly affect photosynthetic rate, leaf lifespan, or branch characteristics.
Leaf morphology is identified as a key adaptive trait in response to phenological gaps.
Abstract
Phenological gaps exert a significant influence on the growth of dwarf bamboos. However, how dwarf bamboos respond to and exploit these phenological gaps remain enigmatic. The light environment, soil nutrients, leaf morphology, maximum photosynthetic rate, foliage dynamics, and branching characteristics of Sasa kurilensis were examined under the canopies of Fagus crenata and Magnolia obovata. The goal was to elucidate the adaptive responses of S. kurilensis to phenological gaps in the forest understory. The findings suggest that phenological gaps under an M. obovata canopy augment the available biomass of S. kurilensis, enhancing leaf area, leaf thickness, and carbon content per unit area. However, these gaps do not appreciably influence the maximum photosynthetic rate, total leaf number, leaf lifespan, branch number, and average branch length. These findings underscore the significant…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
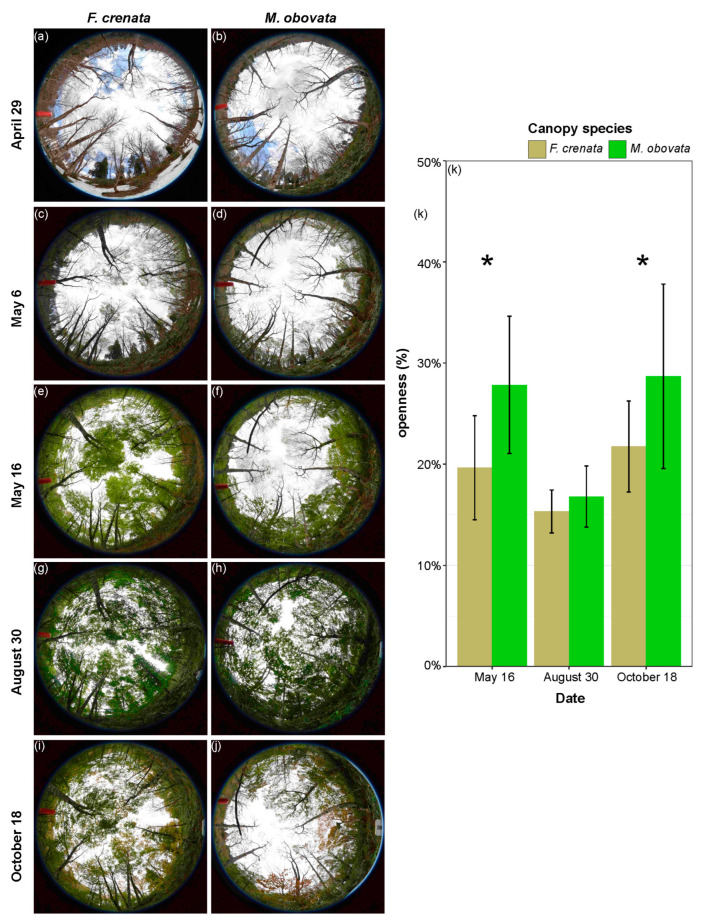 Figure 1
Figure 1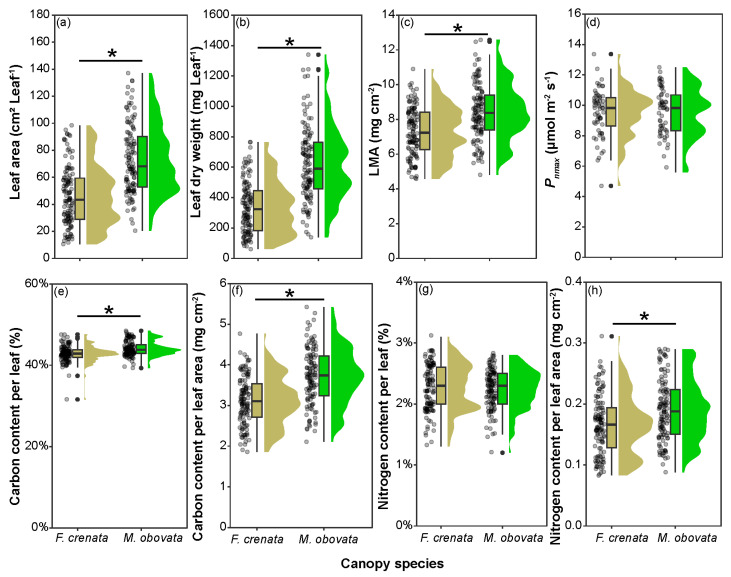 Figure 2
Figure 2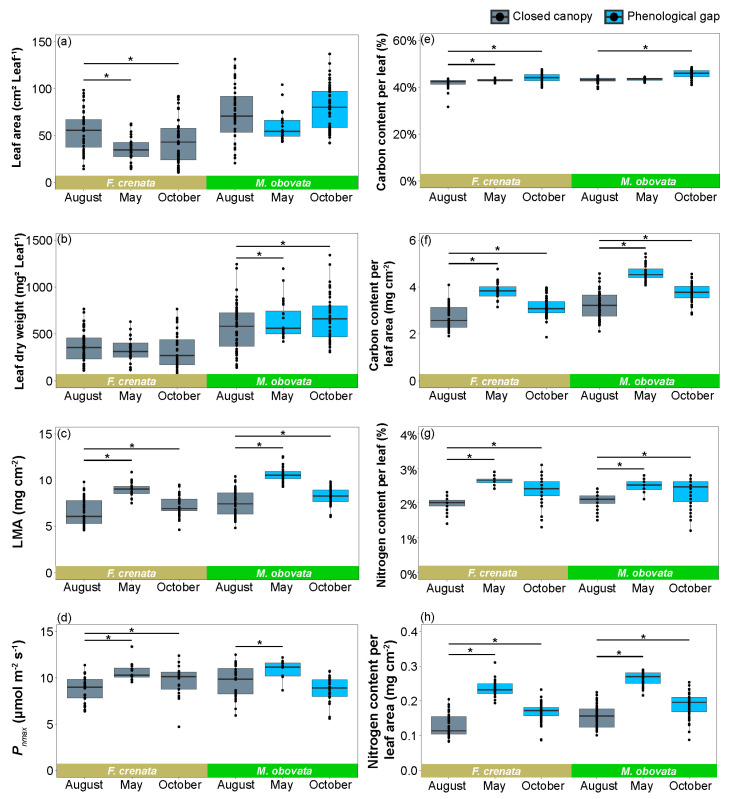 Figure 3
Figure 3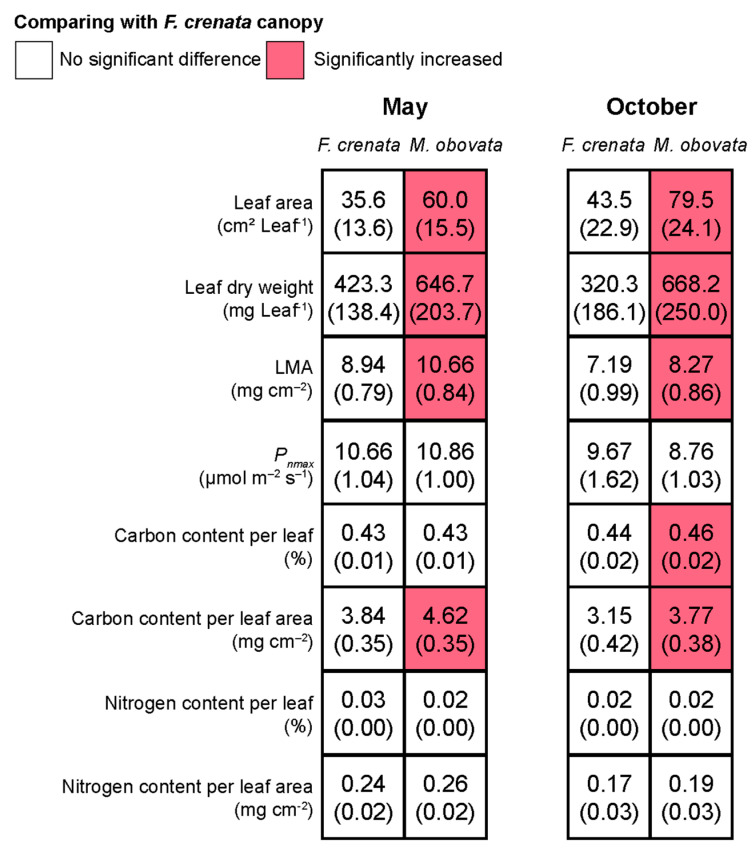 Figure 4
Figure 4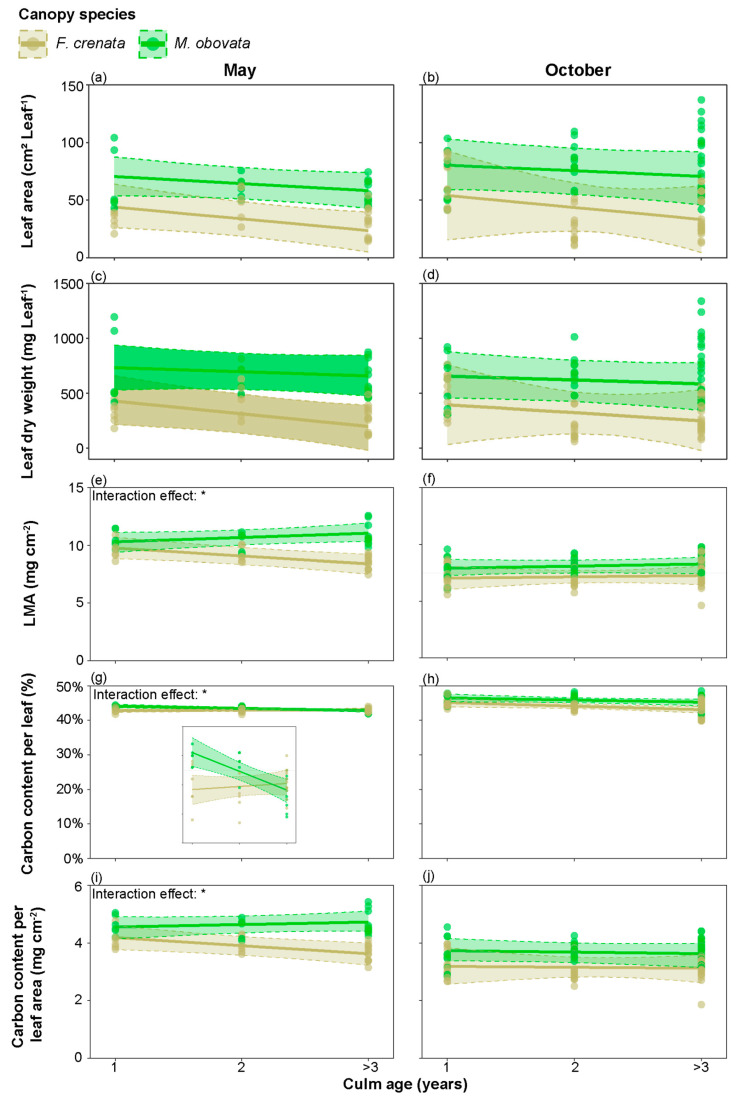 Figure 5
Figure 5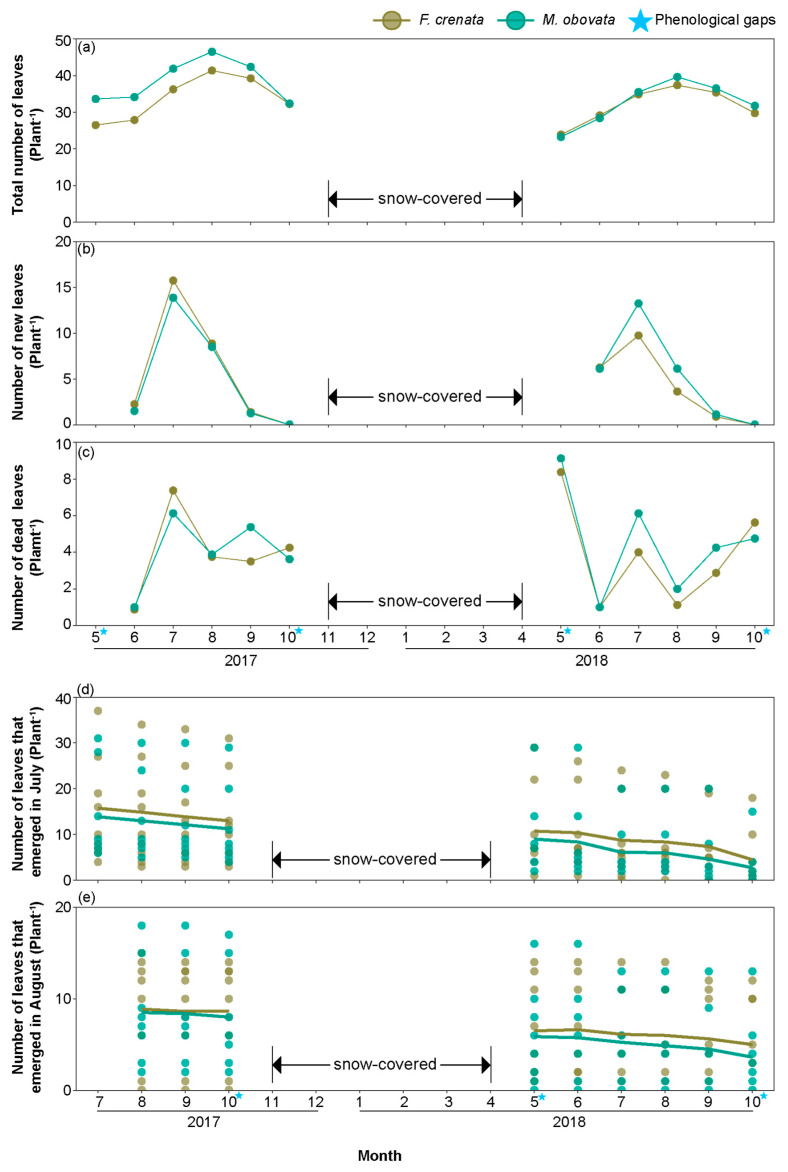 Figure 6
Figure 6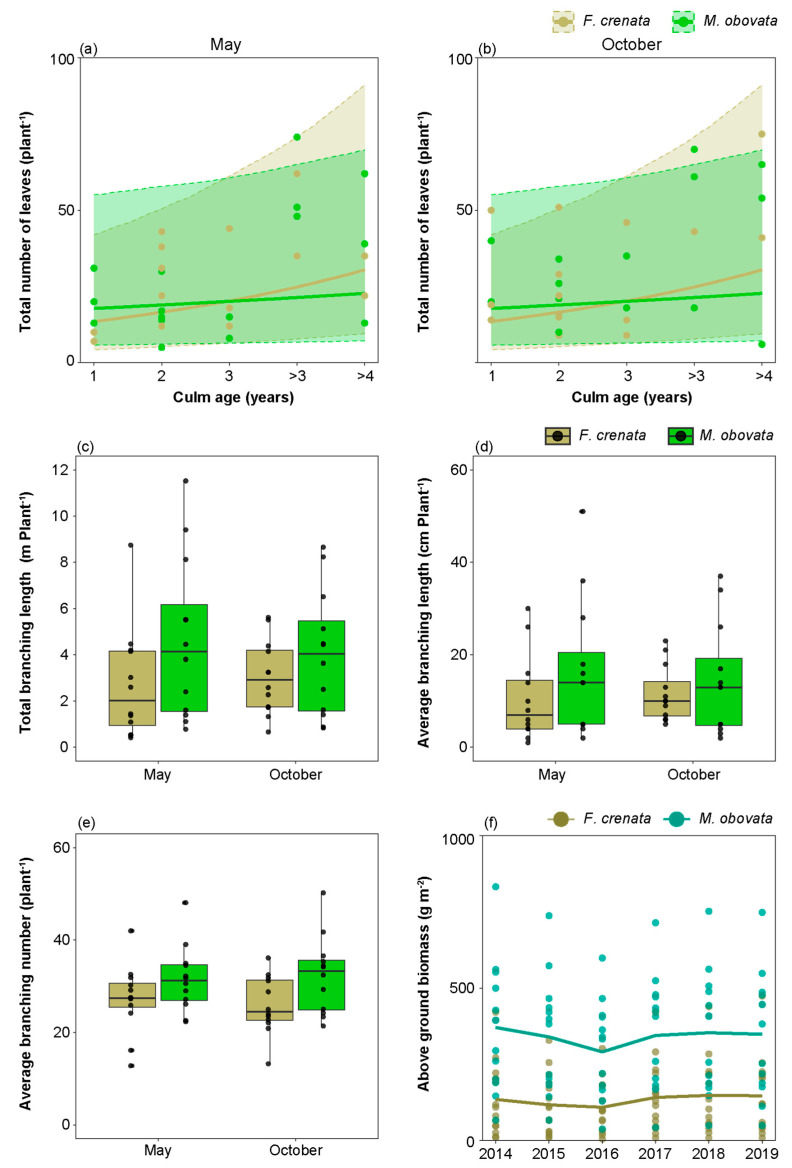 Figure 7
Figure 7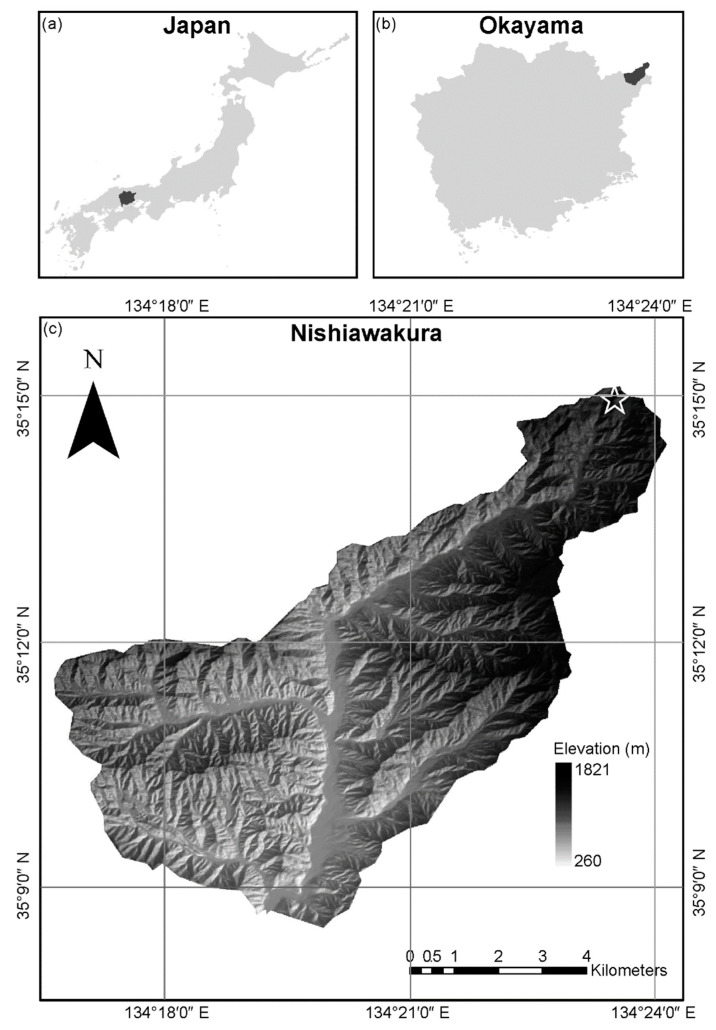 Figure 8
Figure 8- —National Natural Science Foundation of China
- —Japan Society for the Promotion of Science
- —National Key Research and Development Program of China
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBamboo properties and applications · Plant Water Relations and Carbon Dynamics · Ecology and Vegetation Dynamics Studies
1. Introduction
Dwarf bamboos are widely found in the subalpine forests and certain subtropical mountain forests of Japan, South America, and Southwest China [1,2,3,4,5,6]. Dwarf bamboos typically occur as a dominant species covering forest floors with high density; as clonal plants, dwarf bamboos can share photosynthetic products between different culms through underground rhizomes, with culms spanning hundreds of square meters potentially belonging to the same genotype [7]. The shading effect and slowly decaying litter can inhibit the regeneration of some tree species, acting as ecological filters [8,9]. Dwarf bamboo forests also play other significant roles in forest ecosystems. For instance, more than 50% of the fine-root biomass in the surface soil layer was attributed to dwarf bamboos, which played a crucial part in the nutrient cycle [10]. Furthermore, when trees are felled, dwarf bamboos can mitigate nitrogen leaching from soil to rivers [11]. The interactions of dwarf bamboo with various organisms are also important factors affecting forest ecosystems.
One of the most important causes of the dominance of dwarf bamboo communities in covering forest floors is their strong adaptability to changing light environments, whether under closed or open canopies with limited or abundant light intensity, respectively. The culm density, culm height, and biomass of Sasa senanensis were found higher in open sites than under closed canopies [12], while the net photosynthetic rate and leaf dry mass per unit area (LMA) were significantly lower in leaves under closed canopies than in canopy gaps [13]. The maximum photosynthetic rates of Pleioblastus pubescens were lower and the leaf lifespan was longer in shaded conditions than in open conditions [14,15]. The net photosynthetic rate and LMA were significantly lower in leaves under closed canopies than in canopy gaps in Sasa kurilensis, and the C/F ratio of ramets (the ratio of non-photosynthetic to photosynthetic parts) under a closed canopy was lower than in canopy gaps. The above research elucidates the plasticity of dwarf bamboo between high- and low-light environments from multiple perspectives.
Currently, research on the physiological and ecological characteristics of dwarf bamboos has mainly focused on differences in low- and high-light environments. However, a phenological phenomenon exists in deciduous broad-leaved forests that has not yet received widespread attention. In these forests, the canopy of Fagus crenata completes leaf-emergence earlier in early spring and leaf-fall later in autumn than that of Magnolia obovata. This results in an increase in temporal light availability under the M. obovata canopy during these periods. [16,17,18,19]. Tomita and Seiwa (2004) have shown that S. kurilensis and Sasa palmata show a higher Sasa quantity index under M. obovata canopies than under F. crenata canopies [18]. The culm density and biomass of S. kurilensis are greater under M. obovata and Betula grossa canopies than under F. crenata canopies [16]. Only a few studies have indicated the impact of the phenological gap on the community structure of dwarf bamboo growing on the forest floor.
The ability of plants to adapt to heterogeneous light environments is crucial for their survival and reproduction. This adaptation is achieved primarily through adjustments in leaf morphology [20,21,22,23], photosynthetic performance [23,24], and changes in leaf chemical composition [21,24]. There have been no investigations into the adaptations of leaf characteristics to such temporally heterogeneous light environments in mixed-species canopies. The objectives of this study are as follows: (1) to confirm whether phenological gaps affect the leaf characteristics, leaf dynamics, and branching characteristics of S. kurilensis; (2) to investigate whether these effects vary with the culm age of S. kurilensis; and (3) to discuss the plasticity of S. kurilensis leaves and branches in response to phenological gaps and understand their survival strategies under the forest canopy.
2. Results
2.1. Phenological Gaps and Microhabitat Conditions under Different Canopy Species
Figure 1a–j shows the process of how phenological gaps occur from April to October using hemispherical photos. Both F. crenata and M. obovata canopies were still leafless on 24 April, while the ground was still covered with snow. By 6 May, the F. crenata canopy had started to leaf out, while the M. obovata canopy remained leafless. On 16 May, the F. crenata canopy had fully leafed out, while the M. obovata canopy was just beginning to leaf out. We believe that at this time, a phenological gap had formed beneath the M. obovata canopy, with significantly higher values of canopy openness compared to those beneath the F. crenata canopy (Figure 1k; Supplementary Table S1). By 30 August, both the F. crenata and M. obovata canopies had closed, with no significant differences in canopy openness beneath the canopies. Another phenological gap occurred around 18 October, as the M. obovata canopy had more leaf-drop than the F. crenata canopy, resulting in a significant difference in canopy openness under the trees of these two species (Figure 1k; Supplementary Table S1).
In addition to the light environment, the slope, convexity, soil water content, soil carbon content, and soil nitrogen content of all grids were compared to confirm whether there were other micro-environmental differences between the two types of canopies surveyed here (Table 1, Supplementary Table S2). No significant differences were observed in these other micro-environments. To minimize the impact of other unknown factors between different sub-plots on the dependent variable, a number of sub-plots were treated as random effects in our mixed-effects models. This approach allowed group-level differences to be accounted for while reducing the influence of these unknown factors on the outcome variable.
2.2. Leaf Characteristics under Different Canopy Species
We compared the leaf area, leaf dry weight, LMA, maximum net photosynthetic rate (P_nmax_), carbon content per leaf, carbon content per leaf area, nitrogen content per leaf, and nitrogen content per leaf area of S. kurilensis under each of the two canopy species. Except for P_nmax_ and nitrogen content per leaf, S. kurilensis grown under an M. obovata canopy had significantly higher values than those under an F. crenata canopy (Figure 2; Supplementary Table S3). We then analyzed whether differences had been observed in these leaf characteristics during different seasons based on when the phenological gaps occurred and when canopies were closed (Figure 3; Supplementary Table S4). The results showed that compared to August, LMA, carbon content per leaf area, nitrogen content per leaf, and nitrogen content per leaf area of leaves in May and October had increased significantly under both F. crenata and M. obovata canopies. However, the leaf area in May and October significantly decreased under the F. crenata canopy but not under the M. obovata canopy. The leaf dry weight increased significantly in May and October under the M. obovata canopy, but not under the F. crenata canopy. Similarly, P_nmax_ and carbon content per leaf also showed significant increases in May and October under an F. crenata canopy, but with different tendencies observed under an M. obovata canopy.
2.3. The Influence of Phenological Gaps on Leaf Characteristics
Although various leaf characteristics such as leaf area, leaf dry weight, LMA, and others exhibited significant differences in different seasons, it was unclear whether these differences were a result of intrinsic phenological changes in S. kurilensis or the influence of phenological gaps. Therefore, the differences in leaf characteristics were further compared between the two canopy species in May and October when phenological gaps appeared (Figure 4; Supplementary Table S5). The results showed that in May and October, leaf area, leaf dry weight, LMA, and carbon content per leaf area under an M. obovata canopy were significantly higher than those under an F. crenata canopy. In October, the carbon content per leaf under an M. obovata canopy was also significantly higher than that under an F. crenata. This indicates that these leaf characteristics of S. kurilensis are influenced by the presence of phenological gaps.
Figure 5 shows the interaction effects of canopy species and the culm age of S. kurilensis on five leaf characteristics in May and October (Supplementary Table S6). The results were used to assess whether different culm ages affected the promotion of leaves in phenological gaps. The results showed that in May, the promotion effects on LMA, carbon content per leaf, and carbon content per leaf area under M. obovata were influenced by culm age. Specifically, LMA and carbon content per leaf area increased significantly with culm age, while carbon content per leaf decreased significantly. However, in October, no similar trends were observed in these leaf characteristics.
2.4. Foliage Dynamics, Branching Characteristics and Above Ground Biomass under Different Canopy Species
Figure 6a–c depicts the monthly variations in the total number of leaves, new leaves, and dead leaves of S. kurilensis under the two canopy species analyzed here over two years (Supplementary Table S7). No significant differences were observed in the total number of leaves, new leaves, and dead leaves among different canopy types in all months. However, compared to an M. obovata canopy, the total number of leaves under an F. crenata canopy significantly increased with culm age in May and October (Figure 7a,b; Supplementary Table S8). A greater number of leaves of S. kurilensis emerged in July and August than in other months, and we visualized the changes in the quantities of the leaves that emerged in July and August over two years (Figure 6d,e; Supplementary Table S7). It can be seen that the mortality curves of leaves of the same age were almost identical under different canopies, indicating that almost no difference in leaf lifespan existed between the F. crenata and M. obovata canopies.
The analysis of whether phenological gaps had an effect on the branching characteristics of S. kurilensis found no significant differences in the total branching length, average branching length, and average branching number under the two canopy species in both May and October (Figure 7c–e; Supplementary Table S9).
We estimated the above ground biomass of S. kurilensis in 28 sub-plots from 2014 to 2019 based on the ground diameter. The results showed that under both F. crenata and M. obovata canopies, no significant increasing or decreasing trend in the above ground biomass of S. kurilensis was observed over the six year period, indicating that the S. kurilensis community remained stable under canopies of both species (Figure 7f; Supplementary Table S10).
3. Discussion
3.1. Seasonal Variations in the Leaf Characteristics of S. kurilensis
Our observations and a previous study [18] indicated that the temporal persistence of phenological gaps lasts for approximately 10–20 days (Figure 1). Due to the variation in leaf phenology among many tree species [25], phenological gaps may be widespread in forests. Sasa kurilensis under two canopy species exhibited significant seasonal variations in leaf characteristics (Figure 3); an interesting point is that the seasonal changes in leaf area and leaf dry weight of S. kurilensis are completely different under F. crenata and M. obocata canopies. Compared to August, the LMA of S. kurilensis significantly increased in May and October under both the canopies. Concurrently, the leaf area of S. kurilensis significantly decreased under the F. crenata canopy, while the leaf dry weight of S. kurilensis significantly increased under the M. obovata canopy. Based on previous studies and the results of the present study, it appears that the available light resources and biomass for S. kurilensis under an F. crenata canopy must be less than those under an M. obovata canopy (Figure 1) [16,18]. Sasa kurilensis growing under an M. obovata canopy can enhance the LMA by increasing the allocation of biomass per unit leaf area, whereas S. kurilensis under an F. crenata canopy is constrained by allocatable resources and can only achieve the same result through a reduction in leaf area. Sasa kurilensis, which is covered by snow and experience more than five months of darkness, is highly likely to suffer from photoinhibition or other types of damage when it is suddenly exposed to the year’s strongest light environment during snowmelt [26]. The significant increase in LMA results in thicker leaves, which not only effectively mitigates the effects of photoinhibition [27] but also enhances carbon and nitrogen allocation per leaf area, thereby promoting improved photosynthetic performance [28]. A significant increase in LMA, P_nmax_, and carbon and nitrogen allocation per leaf and per unit leaf area of S. kurilensis were observed in May and October; this is a reasonable adaptation to their environment, regardless of whether under the canopy species of F. crenata or M. obovata.
3.2. The Impact of the Phenological Gap on the Leaf Characteristics of S. kurilensise
The effect of phenological gaps on leaf characteristics is concealed within the inherent seasonal variations of S. kurilensis. During May and October, the phenological gaps increase leaf area, leaf dry weight, LMA, and carbon content per leaf area significantly under an M. obovata canopy (Figure 4); this should be due to the differences in available light resources. We believe that the S. kurilensis under an M. obovata canopy, which benefits from phenological gaps, also aids the S. kurilensis under an F. crenata canopy through physiological integration. We make this inference because in pure forests of F. crenata, the LMA in S. kurilensis is 5.32 ± 0.20 (mg cm^−2^), which is much lower than the results (7.30 ± 1.40 mg cm^−2^) of this survey (Figure 2); this implies that in pure forests without phenological gaps, the leaves of S. kurilensis can become thinner [17]. The creation of phenological gaps improves the light environment, which in turn promotes the dominance of S. kurilensis across the forest floor. This dominance is contingent upon the plasticity of their leaves and their strong ability for physiological integration. In some plant communities, the age structure of the population and the functional roles of individuals can vary under different light environments [29]. Based on this, we considered whether the plasticity of leaves in response to phenological gaps varies with culm age. The results showed that as culm age increased, both leaf area and leaf dry weight decreased almost in parallel in both canopies. Under the F. crenata canopy, the LMA and carbon content per leaf significantly decreased with increasing culm age (Figure 5). For the light-limited environment under an F. crenata canopy, reducing material allocation to older culms to ensure larger leaf area and higher LMA in younger culms for photosynthesis production may be a cost-effective strategy. This capability allows for adaptation to diverse light environments, and studies have shown that S. kurilensis invades subalpine and alpine ecosystems through its high stress tolerance and adaptability to climate change, where significant differences exist in the growth rates of the current year and previous year culms across forest, subalpine, and alpine ecosystems. [30].
3.3. The Impact of Phenological Gaps on the Foliage Dynamics and Branching Characteristics of S. kurilensis
In this study, no significant difference was observed in the total number of leaves and number of new and dead leaves per culm under both canopy species over a two year period (Figure 6a–c). However, considering that the leaves under an M. obovata canopy were larger and thicker, the foliage biomass per plant under an M. obovata canopy was higher than that under an F. crenata canopy, which is consistent with earlier studies. Tobita et al. (2006) reported that dwarf bamboo grown in high-light environments has greater leaf biomass per unit area (g m^−2^) than those grown in low-light environments [13].
This trend has also been observed in other plant species [31]. Sasa kurilensis under the canopy of M. obovata invested in larger and thicker leaves to increase photosynthetic performance, rather than increasing the number of leaves. Sasa kurilensis does not produce new leaves repeatedly on the same branch, and every year leaves emerge on new branches. Although we did not make precise calculations, it is evident that this approach requires the plants to expend more energy and resources. Some components in leaves, such as nitrogen, can even be recycled with an individual plant according to demand [32], but it is unlikely that the material allocated to new branches can be recycled. Our analysis of branching characteristics also confirms this inference, because no difference was observed in total and average branch length alone with the number of branches between S. kurilensis under both the canopy types analyzed here (Figure 7c–e). Although the M. obovata canopy has phenological gaps that can temporarily result in an increase in available light resources, most of the time, M. obovata still has a closed canopy, as does F. crenata. Even if increasing branch length places leaves in more extended positions, little may be gained in the production of photosynthetic products during the closed-canopy period. Additionally, more branches may increase the risk of intraspecific competition and self-shading [33,34,35,36,37].
In general, thicker leaves are associated with longer lifespans [38] because the investment in leaf tissue needs to be recovered before the leaf dies. The results of the present study showed that the leaf lifespans of S. kurilensis under both canopy species were almost identical (Figure 6), and estimated to be around 2–3 years. Sasa kurilensis under an F. crenata canopy allocated fewer resources to leaves but achieved the same leaf lifespan as S. kurilensis under an M. obovata canopy. We considered two reasons for this: first, the return on investment under an F. crenata canopy was higher; second, physiological integration provided resource compensation, extending the lifespan of leaves under an F. crenata canopy. Evidence to support this has been found in other studies; for example, culm lifespan was significantly longer under low- than under high-light conditions [6]. The advantage of this strategy was that it allowed for the long-term recovery of the initial investment when the culm lifespan was long. During the years the present investigation was conducted, no signs of decline were observed in the S. kurilensis community under both canopy species analyzed here (Figure 7f). Sasa kurilensis under the M. obovata canopy expands its leaf area to adapt to the transient appearance of phenological gaps, thereby gaining more photosynthetic products. Our results also imply that these photosynthetic products might be shared with other culms experiencing a greater scarcity of light resources through physiological integration. While we cannot confirm this hypothesis at present, we believe that the strong plasticity of S. kurilensis leaves is one of the key reasons for its ability to occupy the forest floor.
4. Materials and Methods
4.1. Study Area
The study was conducted in an old-growth beech forest located in Wakasugi Forest Reserve, at 35°14′ N, 134°23′ E, at 1048 m above sea level, in the Chugoku Mountains near the village of Nishiwakura, in Okayama Prefecture, in western Japan (Figure 8). The soil is a brown forest soil on granite rock substrate. The site falls within the cool temperate zone, with a mean temperature of approximately 8.2 °C and an average annual precipitation of 2400 mm. In winter, S. kurilensis is completely covered by accumulated snow. The canopy layer of the forest is dominated by F. crenata, M. obovata, and B. grossa [39]. The forest floor is densely covered with S. kurilensis, with F. crenata, Acer japonicum, Acer mono, and Viburnum furcatum as the dominant species in the canopy layer.
4.2. Establishment of Study Plot
A rectangular 10 m × 120 m survey plot was established, which was subdivided into 48 sub-plots, each measuring 5 m × 5 m; in addition, a 2 m × 2 m sub-plot was established at the center of each larger 5 m × 5 m sub-plot. The assessments of the light environment and soil nutrients were conducted in the 5 m × 5 m sub-plots, while all other investigations were performed within the 2 m × 2 m sub-plots (Supplementary Figure S1).
4.3. Leaf Characteristics
In August 2017, October 2017, and May 2018, leaves that were not shaded, from culms aged one, two, and over three years, were randomly sampled in the sub-plots (Supplementary Figure 1). Throughout the study, a total of 238 leaves from 65 culms with different ages were randomly collected and taken back to the laboratory for measurements. The leaves were scanned and their area was determined using imaging software (LIA for Win 32 version 0. 3781 by K. Yamamoto, Nagoya Univ., Nagoya, Japan). Subsequently, the leaves were dried at 70 °C for more than 48 h using an oven, and their dry weight was measured. The LMA, leaf area, and dry weight per leaf were then calculated for each individual leaf. Carbon and nitrogen content in each leaf were determined using an elemental analyzer (2400 II CHNS/O, Perkin-Elmer, Norwalk, CT, USA).
4.4. Leaf Maximum Net Photosynthetic Rate
The experiments on the P_nmax_ of non-shaded S. kurilensis leaves were taken during the appearance of phenological gaps in May and October and during closed-canopy conditions in August in both 2016 and 2017 (Supplementary Figure S1). To ensure consistency, measurements for each season were completed within a consecutive five day period. The measurements were conducted on sunny days between 9:00 and 11:30 AM, ensuring that the leaves had been unshaded for over an hour under natural conditions before sampling. Prior to measurements, leaves were exposed to a light intensity of 1800 μmol m^−2^ s^−1^ for half an hour using an open gas exchange system (LI-6400, LI-COR, Lincoln, NE, USA) to fully open the stomata before measuring maximum photosynthesis rates (this intensity was confirmed in a preliminary experiment to not cause photoinhibition in S. kurilensis leaves). A total of 120 non-shaded leaves were measured under F. crenata and M. obovata canopies. The P_nmax_ was measured under the following conditions: a photosynthetic photon flux density of 1600 μmol m^−2^ s^−1^, a CO_2_ concentration of 370 parts per million, and a relative humidity of 65–75%. The leaf temperature was set to be the same as the air temperature.
4.5. Foliage Dynamics and Branching Characteristics
A two year survey (2017–2018) was conducted to investigate the foliage dynamics of S. kurilensis under two canopy species listed above (Supplementary Figure S1). For each species, we randomly selected eight non-shaded culms of different ages (one, two, three, and over four years). The study site, where S. kurilensis grows, experiences snow cover from December to April every year. The total number of leaves on each culm was recorded every month from May to October, while dead and new leaves were accurately tracked by marking them with different colored markers. In addition, we investigated the branching characteristics of 12 randomly selected culms under each canopy species, recording the number and length of all branches during both phenological gaps (May and October) and closed-canopy (August) conditions in 2018.
4.6. Estimation of Above Ground Biomass
We marked all individuals of S. kurilensis growing in each sub-plot and carried out a culm tracking survey every October from 2014 to 2019 (Supplementary Figure S1). The survey included ground diameter and the numbers of surviving, dead, and new culms. The above ground biomass of S. kurilensis in each of the 28 smaller sub-plots was estimated based on ground diameter measurements. The data of above ground biomass for the years 2014 and 2015, as well as the calculation method for biomass (Equation (1)), can be found in Wu et al. [16].
W: above ground biomass D: ground diameter.
4.7. Light Environment and Microhabitat Conditions
Canopy openness was measured at a height of 3 m at the center of each 28 sub-plot (Supplementary Figure S1). Hemispherical photographs were taken on 29 April, 6 May, 16 May, 30 August, and 18 October 2017 to document when phenological gaps occurred or disappeared. Canopy openness was calculated using LIA32 analysis software.
An investigation into the soil nutrient conditions of each grid was conducted in October 2018 (Supplementary Figure S1). Soil samples were collected from the four corners and the center of each 5 m × 5 m sub-plot, the A0 layer was removed, and approximately 50 g of hard soil was collected from a depth up to 50 cm; each set of five samples from one sub-plot were subsequently combined prior to analysis. Stones and plant roots were removed using a 2 mm sieve and the soil was weighed in its wet state. After drying for one week in a well-ventilated room, the soil was placed in an oven and dried at 70 °C for 48 h. The dry weight of soil sample was measured and the moisture content of each sample was calculated. The carbon and nitrogen content of soils were measured using a CN coder (JM1000CN, J-Science Laboratory, Kyoto, Japan). Slope and convexity in each of the 28 grids were derived from Akaji et al. [40].
4.8. Model Analysis
To investigate the impact of phenological gaps on the individual leaves, foliage, and branching characteristics of S. kurilensis, a generalized linear mixed model (GLMM) was constructed using the Bayesian Markov Chain Monte Carlo method. The use of Bayesian modelling methods present various benefits, such as allowing for distribution-type selection of the data, and providing effective data fit even with limited sample numbers. Hence, Bayesian analyses are widely utilized across numerous fields of study [41,42,43]. The dependent variables included leaf area (cm^2^ leaf^−1^), leaf dry weight (mg leaf^−1^), LMA (mg cm^−2^), P_nmax_ (μmol m^−2^ s^−1^), carbon and nitrogen content per leaf (%), carbon and nitrogen content per leaf area (mg cm^−2^), total number of leaves (plant^−1^), number of dead and new leaves (plant^−1^), total length of branches (m plant^−1^), average length of branches (cm), average number of branches (plant−1), and above ground biomass (g m^−2^). The independent variables were canopy species (F. crenata and M. obovata), season (May, August, and October), and culm age (one, two, three, over three, and over four years). For analyses of the photosynthetic performance of leaves, the random effect was set to individual S. kurilensis identifiers; for other analyses, the random effect was set to sub-plot identifiers. Supplementary Table S11 contains detailed information on all model structures. Topography of each sub-plot was set as a random effect when significant differences were observed between two canopy species. In the absence of such significant differences, the number of sub-plots was set as a random effect (Supplementary Table S12 and S13). For each model, we ran four independent chains with one interval, and obtained 5000 posterior samples from each chain after a burn-in of 5000 iterations using non-informative prior distributions since we did not have any prior information for our data. We considered a 95% credible interval that did not include zero to be statistically significant for the mean of the variables. We calculated the R-hat of all parameters and ensured that it was less than 1.1. All models were performed using the brms package [44] on R version 4.2.2 and R Studio version 1.3.1093.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Franklin J.F. Maeda T. Ohsumi Y. Matsui M. Yagi H. Hawk G.M. Subalpine coniferous forests of central Honshu, Japan Ecol. Monogr.19794931133410.2307/1942487 · doi ↗
- 2Nakashizuka T. Numata M. Regeneration process of climax beech forests: I. Structure of a beech forest with the undergrowth of Sasa Jpn. J. Ecol.198232576710.18960/seitai.32.1_57 · doi ↗
- 3Wada N. Dwarf bamboos affect the regeneration of zoochorous trees by providing habitats to acorn-feeding rodents Oecologia 19939440340710.1007/BF 0031711628313678 · doi ↗ · pubmed ↗
- 4Noguchi M. Yoshida T. Factors influencing the distribution of two co-occurring dwarf bamboo species (Sasa kurilensis and S. senanensis) in a conifer-broadleaved mixed stand in northern Hokkaido Ecol. Res.200520253010.1007/s 11284-004-0009-6 · doi ↗
- 5Kudo G. Amagai Y. Hoshino B. Kaneko M. Invasion of dwarf bamboo into alpine snow-meadows in northern Japan: Pattern of expansion and impact on species diversity Ecol. Evol.20111859610.1002/ece 3.922393485 PMC 3287379 · doi ↗ · pubmed ↗
- 6Tsunoda Y. Furukawa S. Mizunaga H. How does the longevity of Sasa kurilensis ramets respond to a light gradient? An analysis of ontogenetic changes to hydraulic resistance and carbon budget within a ramet Ecol. Res.20173211712810.1007/s 11284-016-1423-2 · doi ↗
- 7Matsuo A. Suyama Y. Makita A. The relationship between rhizome habits and spatial distribution pattern of genets in two dwarf bamboos Jpn. J. Ecol.201060818810.18960/seitai.60.1_81 · doi ↗
- 8ItôH. Hino T. Dwarf bamboo as an ecological filter for forest regeneration Ecol. Res.20072270671110.1007/s 11284-006-0066-0 · doi ↗