Biological Effects of Corticosteroids on Pneumococcal Pneumonia in Mice and Humans
Hiroki Taenaka, Katherine D. Wick, Aartik Sarma, Shotaro Matsumoto, Rajani Ghale, Xiaohui Fang, Mazharul Maishan, Jeffrey E. Gotts, Charles R. Langelier, Carolyn S. Calfee, Michael A. Matthay

TL;DR
This study shows that corticosteroids reduce lung damage and inflammation in pneumococcal pneumonia in both mice and humans, without increasing bacterial load.
Contribution
The study identifies specific biological mechanisms by which corticosteroids benefit patients with pneumococcal pneumonia.
Findings
Dexamethasone reduced pulmonary edema, lung permeability, and hypoxemia in mice with pneumonia.
Steroid therapy altered inflammatory responses at both the gene and protein levels.
Findings in mice were consistent with transcriptomic changes observed in human patients.
Abstract
Streptococcus pneumoniae is the most common bacterial cause of community acquired pneumonia and the acute respiratory distress syndrome (ARDS). Some clinical trials have demonstrated a beneficial effect of corticosteroid therapy in community acquired pneumonia, COVID-19, and ARDS, but the mechanisms of this benefit remain unclear. The objective of this study was to investigate the effects of corticosteroids on the pulmonary biology of pneumococcal pneumonia in an observational cohort of mechanically ventilated patients and in a mouse model of bacterial pneumonia with Streptococcus pneumoniae. We studied gene expression with lower respiratory tract transcriptomes from a cohort of mechanically ventilated patients and in mice. We also carried out comprehensive physiologic, biochemical, and histological analyses in mice to identify the mechanisms of lung injury in Streptococcus pneumoniae…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
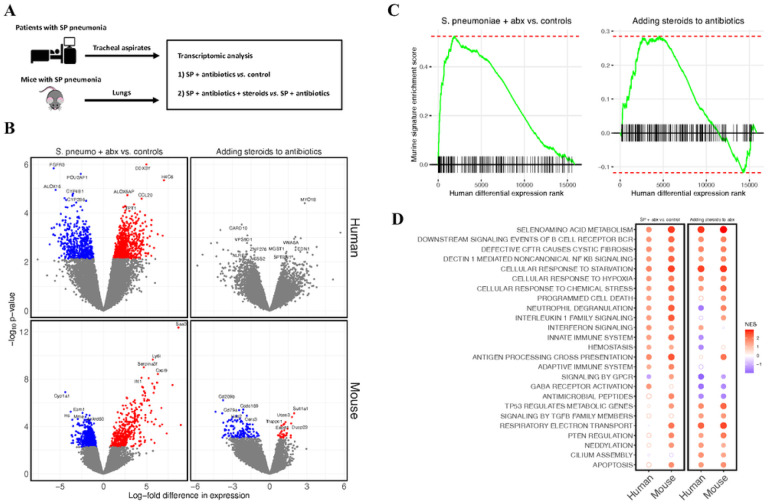 Figure 1
Figure 1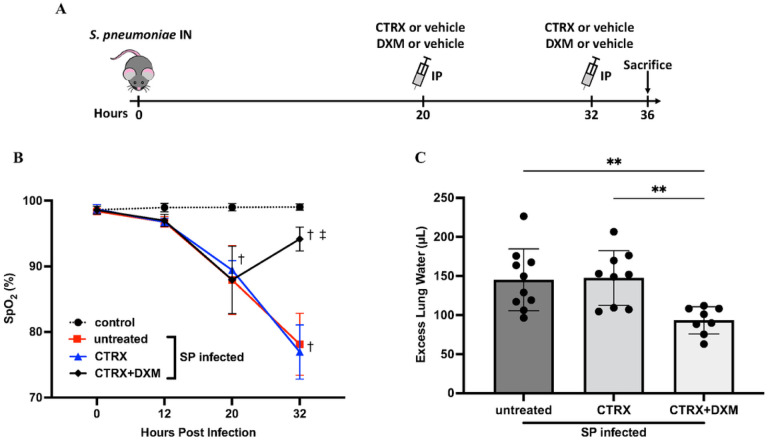 Figure 2
Figure 2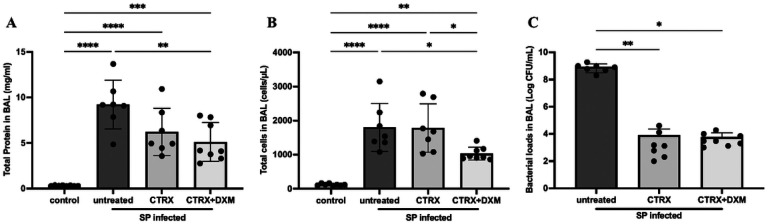 Figure 3
Figure 3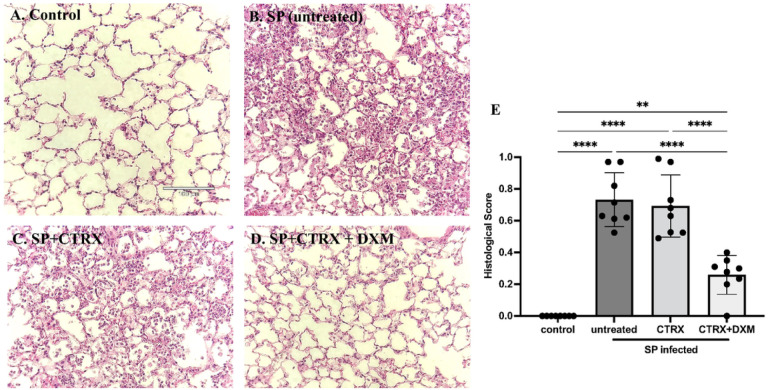 Figure 4
Figure 4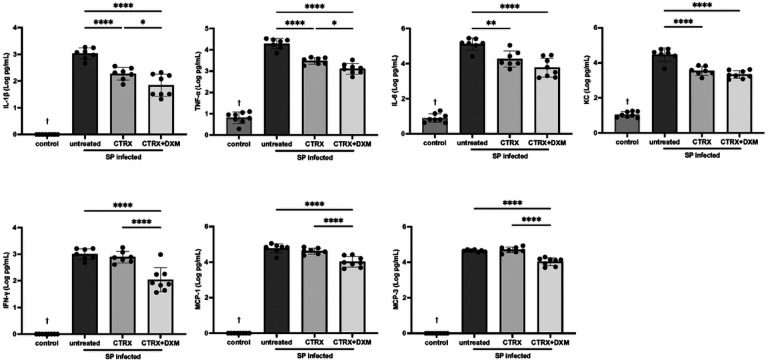 Figure 5
Figure 5Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPneumonia and Respiratory Infections · Lung Cancer Research Studies · Pleural and Pulmonary Diseases
INTRODUCTION
Bacterial pneumonia is a frequent cause of severe respiratory failure and acute respiratory distress syndrome (ARDS) [1]. The most common cause of bacterial community acquired pneumonia (CAP) is Streptococcus pneumoniae [2, 3], which directly damages the lung epithelium and induces the release of a substantial number of cytokines and chemokines from epithelial cells and macrophages, resulting in protein exudation and edema formation in the lung [1, 4]. Despite effective antibiotic therapy, the dysregulated pathogen-host interaction may still cause life-threatening acute respiratory failure [5].
Corticosteroids are sometimes prescribed as an adjunctive therapy for severe pulmonary infections and ARDS. The clinical benefit of adjunctive steroid therapy for pneumococcal infections has been previously demonstrated in meningitis [6]. Recent clinical trials have demonstrated therapeutic benefits of dexamethasone, a long-acting glucocorticoid, on mortality in severe COVID-19 infection [7], non-COVID ARDS [8], and in severe CAP [9–11]. However, the mechanisms by which steroids improve outcomes in patients with severe pneumonia or ARDS are not well established, and few studies have comprehensively characterized the effects of steroids in the respiratory tract of critically ill patients [12, 13] or clinically relevant animal models using bacterial pathogens treated with antibiotic therapy [14, 15].
Corticosteroids are typically used to potently suppress systemic immune responses, but they have additional effects that may be relevant to lung injury, including effects on wound healing, modifying lung fluid balance and the extent of pulmonary edema, and metabolism [16, 17]. There is emerging evidence that corticosteroids have distinct effects on systemic vs. pulmonary inflammation [18, 19]. Given that corticosteroids have pleiotropic and lung-specific effects, elucidating the effects of corticosteroids on the injured lung in bacterial pneumonia is important for establishing a biologic rationale for further clinical application of steroids in severe lung infections. Understanding the specific mechanisms of corticosteroid benefit should help identify the patients with pneumonia or ARDS most likely to benefit from steroids.
To investigate the effects of corticosteroids in bacterial pneumonia, we studied lower respiratory tract gene expression in an observational cohort of mechanically ventilated patients with and without S. pneumoniae pneumonia and in mice with an established mouse model of pneumococcal pneumonia. We also carried out comprehensive biochemical, physiological, and histologic studies in the mice to understand the effects of steroids in pneumococcal pneumonia.
METHODS
See the online supplement for more detailed methods.
Observational cohort
We studied tracheal aspirate gene expression from samples collected from an observational cohort of mechanically ventilated adults within 72 hours of admission to the ICU at UCSF Medical Center. The study was approved by the UCSF Institutional Review Board (17–24056). We included all subjects who had S. pneumoniae in a respiratory tract culture or met criteria for S. pneumoniae infection using a metagenomic sequencing-based model as previously described [20]. We also included as controls subjects who were mechanically ventilated for neurologic injury, were not immunocompromised, received no immunosuppressive medications, and had no evidence of respiratory disease on chest radiograph (Fig. 1A).
Experimental Model in Mice, Bacterial Infection, and Treatment:
The protocol was approved by the University of California, San Francisco Institutional Animal Care and Use Committee (No.AN189182). C57BL/6 mice were randomly divided into four groups: (1) Healthy control; (2) S. pneumoniae; (3) S. pneumoniae + ceftriaxone; (4) S. pneumoniae + ceftriaxone + dexamethasone. Mice in groups 2–4 were anesthetized and inoculated intranasally with 10^8^ CFU of live S. pneumoniae serotype 19F. Mice received 10 mg/kg of dexamethasone or vehicle control in combination with 150 mg/kg of ceftriaxone or vehicle control intraperitoneally 20 and 32 hours after inoculation. In some experiments, body temperature, body weight, and oxygen saturation by pulse oximetry (SpO_2_) were measured 12, 20, and 32 hours after inoculation. All mice were sacrificed 36 hours after infection or earlier if pre-specified criteria indicating unacceptably severe illness were met. Figure 2A depicts experimental procedures.
Lung Injury Endpoints
Mice underwent overdose of ketamine and xylazine, bilateral thoracotomy, and exsanguination by right ventricular puncture. In some mice, lungs were removed and homogenized to measure excess extravascular lung water (ELW) [21, 22]. In other mice, the lungs were lavaged, and bronchoalveolar lavage (BAL) protein was measured with the BCA Protein Assay (Thermo Fisher). BAL cell count was measured with a Coulter counter. Postmortem bacterial titers of BAL were measured by serial dilution and plaque counting on sheep blood agar plates. For histology, the lungs were fixed by 4% paraformaldehyde and stained with hematoxylin-eosin. Lung injury was assessed by an observer (XF) who was blinded to the experimental groups, based on the scoring system partly modified from the report of the American Thoracic Society [23]. BAL inflammatory cytokines and chemokines were measured using a FLEXMAP 3D^™^ (Luminex). Receptor for advanced glycation end products (RAGE) in BAL was measured using the Quantikine ELISA kit (R&D Systems). Alveolar fluid clearance was measured according to previously published methods [24, 25].
RNA Sequencing
In mice, the whole lung was placed in RNA Shield, frozen, homogenized, and then samples extracted using Quick-RNA mini prep plus kit (Zymo research). In humans, tracheal aspirate samples were collected on the day of enrollment and stored at −80C in RNAse-free conditions [26]. RNA was extracted using the Allprpep kit (Qiagen and then ribodepleted using FastSelect (Qiagen) before undergoing library preparation using the NEBNext Ultra II RNASeq Kit (New England Biolabs) and paired-end Illumina sequencing on an Novaseq 6000. Human and mouse samples were sequenced separately.
Bioinformatics analysis
Transcriptomic analysis was performed using established bioinformatics pipelines. Briefly, FASTQ files were aligned using STAR and count matrices were generated using tximport. Differential expression between groups was determined using limma-voom, and fgsea was used to identify pathways enriched in each experimental condition. CEMiTool was used to perform an unsupervised network analysis. A Benjamini-Hochberg adjusted p-value < 0.1 was considered significant for all analyses. Full details are provided in online supplement.
Statistical analysis
All data were tested for normality with Shapiro-Wilk tests. Comparisons of each group were made with one-way ANOVA followed by Tukey’s multiple comparisons tests for normally distributed parameters, or with Kruskal-Wallis test followed by Dunn’s multiple comparisons for skewed parameters. Two-way analysis of variance for repeated measures followed by Tukey’s multiple comparison test was used to evaluate the effect of time and group. All tests were two-tailed, and differences were considered to be statistically significant when p < 0.05.
RESULTS
Human subjects
Tracheal aspirate transcriptomes were available from 159 mechanically ventilated subjects, of whom 15 met inclusion criteria. Ten of the patients had clinically adjudicated S. pneumoniae pneumonia, in whom five the pathogen was detected by culture and five by metagenomic sequencing [20, 27] (Table S1). All patients were treated with antibiotics active against S. pneumoniae prior to sample collection, and five patients (three culture-positive) also received corticosteroids prior to sample collection. Samples were also available from five uninfected control subjects.
RNA sequencing identifies reproducible effects of both pneumococcal pneumonia and steroid treatment in human patients and mice
S. pneumoniae infection was associated with markedly altered gene expression in the lower respiratory tracts of both humans and mice (Fig. 1B). There were 1,184 genes differentially expressed between patients with S. pneumoniae who were not treated with steroids versus uninfected controls, and 951 genes differentially expressed between infected mice treated with ceftriaxone and uninfected controls. No individual gene was differentially expressed between pneumonia patients who received steroids versus those who did not, while 162 genes were differentially expressed in mice treated with dexamethasone and ceftriaxone compared to mice treated with ceftriaxone alone (Fig. 1B). Gene set enrichment analysis (GSEA) identified notable overlaps between gene expression from clinical samples and gene expression in the mouse model (Fig. 1C), suggesting that the experimental model replicated biological features of clinical disease and steroid treatment. The top 100 genes upregulated in patients with S. pneumoniae pneumonia compared to controls were significantly enriched in the S. pneumoniae mouse model compared to control mice (p = 1.2×10^− 5^). The top 100 genes upregulated in patients with pneumococcal pneumonia who received steroids compared to patients with pneumococcal pneumonia who did not receive steroids were significantly enriched in mice treated with dexamethasone and ceftriaxone compared to mice treated with ceftriaxone alone (p = 1.9×10^− 12^).
GSEA identified enrichment of several pathways shared between humans and mice (Fig. 1D). For example, in both mice and humans, S. pneumoniae infection, as expected, increased expression of cellular stress responses and upregulated adaptive and innate immune signaling pathways, including interferon and interleukin 1 family signaling, neutrophil degranulation, and B cell receptor signaling compared to uninfected controls. The addition of steroids to antibiotics increased the expression of pathways associated with injury resolution, including TGF-beta signaling, anti-inflammatory signaling by PTEN, and cilium assembly pathways in humans and in mice. Interestingly, several pathways that were more highly expressed in S. pneumoniae infection compared to controls, including stress and hypoxia responses, non-canonical NF-kB signaling, and B cell receptor signaling were increased further by the addition of steroids. Weighted gene co-expression network analysis (WGCNA) results are described in the Supplementary Material.
Dexamethasone prevents the progression of hypoxemia and pulmonary edema in mice with pneumococcal pneumonia
Oxygen Saturation.
Mice inoculated intranasally with 10^8^ CFU of S. pneumoniae developed hypoxemia at 20 hours after infection. The average oxygen saturation (SpO_2_) declined to approximately 87–91% (Fig. 2B). At 20 hours, mice received either no treatment, ceftriaxone, or ceftriaxone plus dexamethasone. By 32 hours, dexamethasone-treated mice had significantly higher oxygen saturation than those who did not receive dexamethasone. Thus, dexamethasone treatment attenuated progression of arterial hypoxemia 12 hours after treatment, whereas ceftriaxone treatment did not prevent the progression of hypoxemia. Note that arterial hypoxemia in the mice treated with antibiotics alone reached a mean value of 76% at 32 hours whereas dexamethasone treatment increased oxygen saturation to a mean level above 90%.
Pulmonary Edema.
Mice inoculated with S. pneumoniae had approximately 150μL of excess lung water (ELW), demonstrating that 10^8^ of S. pneumoniae induced substantial pulmonary edema within 36 hours after infection. The results for hypoxemia and ELW by treatment group were similar, underscoring the relevance of ELW as a marker of physiologically important pulmonary edema (Fig. 2A, B). Treatment with ceftriaxone alone did not reduce ELW. However, the combination of ceftriaxone and dexamethasone significantly reduced ELW compared to the untreated mice and to ceftriaxone alone (Fig. 2B).
Temperature and Body Weight.
Infected mice became hypothermic (approximately 30°C) at 12 hours after infection and had lower body temperature than normal healthy control mice throughout the experiment. Mice in all infected groups had significant body weight loss compared with normal healthy controls. (Figure S3).
Dexamethasone reduces both alveolar protein permeability and accumulation of leukocytes in the airspaces
Mice with S. pneumoniae had higher protein concentrations in BAL compared with controls, indicating alveolar-capillary barrier disruption (Fig. 3A). The combination of ceftriaxone and dexamethasone significantly decreased BAL protein concentration by one way ANOVA; ceftriaxone alone also decreased BAL protein concentration, though not significantly compared to untreated mice. Total cell counts in BAL showed a substantial increase of white blood cells in the distal airpaces with S. pneumoniae (Fig. 3B). Similarly to the pulmonary edema results, treatment with ceftriaxone alone did not reduce total leukocyte accumulation. However, the combination of ceftriaxone and dexamethasone significantly reduced the accumulation of white blood cells compared to untreated mice and to ceftriaxone alone. Most of the leukocytes were neutrophils (Figure S4).
Bacterial load in the bronchoalveolar lavage (BAL).
Since dexamethasone may impair host defense, we measured the bacterial load in BAL (Fig. 3C). Ceftriaxone substantially reduced the bacterial loads 36 hours after inoculation, as expected. The addition of dexamethasone to ceftriaxone treatment did not increase the bacterial burden in the BAL.
Dexamethasone improves lung histopathology in mice with pneumococcal pneumonia
As quantified by an investigator blinded to the experimental condition, when compared to healthy mice (Fig. 4A), the lungs of mice inoculated with S. pneumoniae had severe inflammation including the infiltration of neutrophils into the airspaces and alveolar septal thickening and alveolar edema fluid at 36 hours after infection (Fig. 4B). Treatment with ceftriaxone alone had no effect on histological markers of lung injury despite its capacity to eliminate bacteria (Fig. 4C). In contrast, the combination of ceftriaxone and dexamethasone significantly and markedly reduced histological markers of lung injury and inflammation and significantly improved the histopathological score compared to untreated controls and to ceftriaxone alone (Fig. 4D, E), findings that are consistent with the results of pulmonary edema, total protein concentration and white blood cells in the BAL.
Alveolar Epithelium – Functional and Biochemical Studies.
To test the effect of dexamethasone on the vectorial fluid clearance capacity of alveolar epithelium [28, 29], we measured alveolar fluid clearance 36 hours after infection. Alveolar fluid clearance was substantially impaired with pneumococcal pneumonia, but there was no significant difference between the untreated and other treatment groups (Figure S5A), indicating that the favorable effects of dexamethasone combined with antibiotics on pulmonary edema, hypoxemia and histology were explained by a reduction in the formation of pulmonary edema not by an increase in the rate of alveolar fluid clearance.
In addition, we measured BAL levels of RAGE, a type I epithelial cell injury marker [30, 31]. There was a significant increase in RAGE in BAL of pneumococcal infected mice (Figure S5B), indicating significant alveolar epithelial cell injury. However, the level of RAGE in BAL was not reduced by ceftriaxone or dexamethasone, a result that supports no significant effect of either treatment on the vectorial fluid transport capacity of the alveolar epithelium.
Dexamethasone attenuates the inflammatory responses in the air spaces of mice with pneumococcal pneumonia
We measured cytokines and chemokines in BAL fluid using a multiplex assay to quantify the inflammatory responses with dexamethasone treatment for pneumococcal pneumonia. Selected cytokines and chemokines with the high relevance to lung injury are displayed in Fig. 5. IL-1β, TNF-α, IL-6, and IFN-γ and chemokines including KC, MCP-1, and MCP-3 were significantly higher in infected mice compared with normal healthy control mice (Fig. 5). Ceftriaxone reduced IL-1β, TNF-α, IL-6, and KC, but did not impact the levels of IFN-γ, MCP-1, and MCP-3. Importantly, the combination of ceftriaxone and dexamethasone significantly attenuated all of these inflammatory cytokine and chemokine accumulation in the air spaces compared with untreated controls and significantly reduced IL-1β, TNF-α, IFN-γ, MCP-1, and MCP-3 as compared to ceftriaxone alone. Additional biomarkers were measured, several of which showed a decrease in BAL concentration with dexamethasone and antibiotic treatment (Figure S6). These results indicate that dexamethasone in combination with appropriate antibiotics reduces the release of pro-inflammatory cytokines and chemokines in pneumococcal pneumonia.
DISCUSSION
This study addresses several important clinically relevant gaps in our knowledge regarding bacterial pneumonia and the effects of corticosteroid treatment. First, prior to this work, limited data were available on the biological overlap of experimental pneumonia in mice compared to patients with pneumococcal pneumonia. Second, few studies have directly evaluated the mechanistic effects of corticosteroids in an experimental model of pneumococcal pneumonia, a particularly important issue in view of the recent clinical trials reporting improved outcomes in patients with severe community acquired pneumonia when treated with corticosteroids [11]. Finally, although patients recognized to have bacterial pneumonia are uniformly treated with antibiotics, animal models using bacterial pathogens do not routinely include the use of antibiotics. This study identifies several mechanisms by which dexamethasone decreases lung injury in a S. pneumoniae model and supports the biological relevance of this model to humans with community acquired pneumococcal pneumonia.
RNA sequencing identified several pathways that were dysregulated by S. pneumoniae infection in both humans and mice. We identified signatures of inflammasome activation, including IL1 signaling and NF-kB activation, and neutrophil degranulation in infected mice and humans. Transcriptomic analysis also identified several pathways modified by the addition of dexamethasone to ceftriaxone. While steroids decreased the expression of some pathways upregulated in S. pneumoniae-infected mice compared to controls, they also increased the expression of other pathways, suggesting the effects of steroids are more complex than simply reversing expression of dysregulated pathways. Notably, tracheal aspirate RNA sequencing from mechanically ventilated patients with S. pneumoniae infection identified similar gene expression changes to those observed in the mouse model, suggesting this experimental system replicates several biological features of clinical disease. In addition, some pathways that were modified by dexamethasone treatment in mice also were also modified in humans treated with steroids compared to those who were not.
The mouse studies of pneumococcal pneumonia demonstrated therapeutic benefits of dexamethasone on oxygenation, pulmonary edema, and lung histology when added to antibiotics. These results are consistent with recent clinical studies, in which early administration of dexamethasone reduced the duration of mechanical ventilation in ARDS patients and reduced the progression to mechanical ventilation in non-intubated patients with severe pneumonia [11, 32, 33]. Assessment of BAL fluid identified that the combination of antibiotics and dexamethasone reduced protein and leukocyte permeability into the distal air spaces. In contrast to the favorable effect on protein permeability, dexamethasone did not enhance alveolar fluid clearance, suggesting that the favorable effects of dexamethasone combined with antibiotics were explained by a reduction in the formation of pulmonary edema, not by an increase in the rate of alveolar fluid clearance.
Because of their immunosuppressive effects, steroids can impair host bacterial clearance [34]. In one mouse study using a very early administration of glucocorticoids (one hour after infection) [14], high dose dexamethasone increased bacterial burden in the lung. In contrast, dexamethasone did not impair host bacterial clearance in the current study, possibly because dexamethasone was given 20 hours after infection and at the same time antibiotic therapy was initiated. We selected this schedule in part to enhance the clinical relevance of the model, as patients with pneumonia usually come to medical attention when their infection has progressed enough to cause symptoms. These findings are consistent with the results of some recent clinical trials of steroids in COVID-19 ARDS [35, 36], non-COVID ARDS [33], and meta-analyses of steroid use in community-acquired pneumonia [37], in which steroid administration provided clinical benefit in patients with pneumonia of differing etiologies but was not associated with significantly higher rates of secondary infection.
Since dexamethasone has pleiotropic effects on the immune system [34], we investigated the impact of dexamethasone on the release of inflammatory cytokines and chemokines. The combination of dexamethasone and ceftriaxone attenuated the release of inflammatory cytokines in BAL fluid, including IL-1β, IL-6, TNF-α, and IFN-γ, and chemokines including KC, MCP-1, and MCP-3. These results are consistent with a prior study of corticosteroid treatment in ARDS patients [38]. Also, the combination of dexamethasone and ceftriaxone significantly reduced the release of IL-1β, TNF-α, IFN-γ, MCP-1, and MCP-3 compared with ceftriaxone alone. An experimental study using a mouse model of pneumococcal pneumonia reported that neutralization of IFN-γ accelerates recovery from lung injury [39]. Growing evidence suggest that MCP-1 is involved in lung inflammatory disorders [40]. The beneficial effect of dexamethasone on lung injury might be attributed in part through suppressing the release of these proinflammatory cytokines and chemokines. Transcriptomic analyses also identified additional pathways, including increased non-canonical NF-kB signaling and shifts in metabolic pathways, that may be important targets for future research.
This study has some limitations. First, the sample size from our clinical cohort is modest. Steroid treatment was non-randomly assigned and likely more commonly administered in sicker patients. In addition, our clinical respiratory tract samples were from tracheal aspirates, while our mouse experimental samples were from whole lung homogenate, which likely decreased our ability to detect differentially expressed genes overlapping between humans and mice. We were unable to identify individual genes that were significantly differentially expressed with steroids in humans. Despite this, we were able to detect changes at the pathway level that were shared between our experimental mouse model and the clinical samples. Second, our experimental studies in mice focused on the early phase of lung injury from pneumococcal pneumonia but did not address survival. This design made it possible to understand the earliest effects of steroid therapy on critical physiologic end points (oxygenation, lung fluid and protein balance), inflammatory responses, lung pathology, and bacterial burden. A recent clinical trial [11] reported a mortality benefit for corticosteroids in severe CAP, so we did not think that survival studies were as critical compared to in depth mechanistic analyses. Future studies will need to test the effects of steroids in other bacterial and viral pathogens in addition to the results from this study with S. pneumoniae.
CONCLUSIONS
In summary, adjunctive therapy with both antibiotics and dexamethasone reduces the severity of hypoxemia, pulmonary edema, and histologic evidence of lung injury in experimental pneumococcal pneumonia compared to antibiotics alone, in part through attenuating the release of inflammatory cytokines and chemokines. Importantly, steroid therapy in combination with antibiotics does not increase bacterial burden in the early phase of pneumococcal pneumonia in mice. Transcriptomic profiling supports the clinical relevance of this mouse model, which replicates several features of pneumococcal pneumonia and steroid therapy in humans. In conclusion, steroid therapy has a sound mechanistic basis as an adjunctive treatment for severe community acquired bacterial pneumonia from S. Pneumoniae with the use of appropriate antibiotics.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Matthay MA, Zemans RL, Zimmerman GA, Arabi YM, Beitler JR, Mercat A, Herridge M, Randolph AG, Calfee CS: Acute respiratory distress syndrome. Nat Rev Dis Primers 2019, 5(1):18.30872586 10.1038/s 41572-019-0069-0PMC 6709677 · doi ↗ · pubmed ↗
- 2Troeger C, Blacker B, Khalil IA, Rao PC, Cao J, Zimsen SRM, Albertson SB, Deshpande A, Farag T, Abebe Z : Estimates of the global, regional, and national morbidity, mortality, and aetiologies of lower respiratory infections in 195 countries, 1990–2016: a systematic analysis for the Global Burden of Disease Study 2016. The Lancet Infectious Diseases 2018, 18(11):1191–1210.30243584 10.1016/S 1473-3099(18)30310-4PMC 6202443 · doi ↗ · pubmed ↗
- 3Ortqvist A, Hedlund J, Kalin M: Streptococcus pneumoniae: epidemiology, risk factors, and clinical features. Semin Respir Crit Care Med 2005, 26(6):563–574.16388428 10.1055/s-2005-925523 · doi ↗ · pubmed ↗
- 4Weiser JN, Ferreira DM, Paton JC: Streptococcus pneumoniae: transmission, colonization and invasion. Nat Rev Microbiol 2018, 16(6):355–367.29599457 10.1038/s 41579-018-0001-8PMC 5949087 · doi ↗ · pubmed ↗
- 5File TM, Ramirez JA: Community-Acquired Pneumonia. New England Journal of Medicine 2023, 389(7):632–641.37585629 10.1056/NEJ Mcp 2303286 · doi ↗ · pubmed ↗
- 6Brouwer MC, Mc Intyre P, de Gans J, Prasad K, van de Beek D: Corticosteroids for acute bacterial meningitis. Cochrane Database of Systematic Reviews 2010(9).10.1002/14651858.CD 004405.pub 320824838 · doi ↗ · pubmed ↗
- 7Recovery Collaborative Group, Horby P, Lim WS, Emberson JR, Mafham M, Bell JL, Linsell L, Staplin N, Brightling C, Ustianowski A : Dexamethasone in Hospitalized Patients with Covid-19. N Engl J Med 2021, 384(8):693–704.32678530 10.1056/NEJ Moa 2021436 PMC 7383595 · doi ↗ · pubmed ↗
- 8Villar J, Ferrando C, Martinez D, Ambros A, Munoz T, Soler JA, Aguilar G, Alba F, Gonzalez-Higueras E, Conesa LA : Dexamethasone treatment for the acute respiratory distress syndrome: a multicentre, randomised controlled trial. Lancet Respir Med 2020, 8(3):267–276.32043986 10.1016/S 2213-2600(19)30417-5 · doi ↗ · pubmed ↗