RGS20 promotes non-small cell lung carcinoma proliferation via autophagy activation and inhibition of the PKA-Hippo signaling pathway
Xiaoyan Ding, Xiaoxia Li, Yanxia Jiang, Yujun Li, Hong Li, Lipeng Shang, Guilin Feng, Huhu Zhang, Ziyuan Xu, Lina Yang, Bing Li, Robert Chunhua Zhao

TL;DR
RGS20 promotes lung cancer growth by boosting cell proliferation through autophagy and disrupting a key signaling pathway.
Contribution
RGS20 is newly identified as a driver of NSCLC proliferation via autophagy and Hippo-PKA signaling inhibition.
Findings
High RGS20 expression correlates with poor survival in NSCLC patients.
RGS20 enhances cell proliferation by increasing autophagy and inhibiting the Hippo signaling pathway.
Inhibiting Hippo signaling mimics RGS20's effects, while GPCR activation reverses them.
Abstract
Novel therapeutic targets are urgently needed for treating drug-resistant non-small cell lung cancer (NSCLC) and overcoming drug resistance to molecular-targeted therapies. Regulator of G protein signaling 20 (RGS20) is identified as an upregulated factor in many cancers, yet its specific role and the mechanism through which RGS20 functions in NSCLC remain unclear. Our study aimed to identify the role of RGS20 in NSCLC prognosis and delineate associated cellular and molecular pathways. Immunohistochemistry and lung cancer tissue microarray were used to verify the expression of RGS20 between NSCLC patients. CCK8 and cell cloning were conducted to determine the proliferation ability of H1299 and Anip973 cells in vitro. Furthermore, Transcriptome sequencing was performed to show enrichment genes and pathways. Immunofluorescence was used to detect the translocation changes of YAP to…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
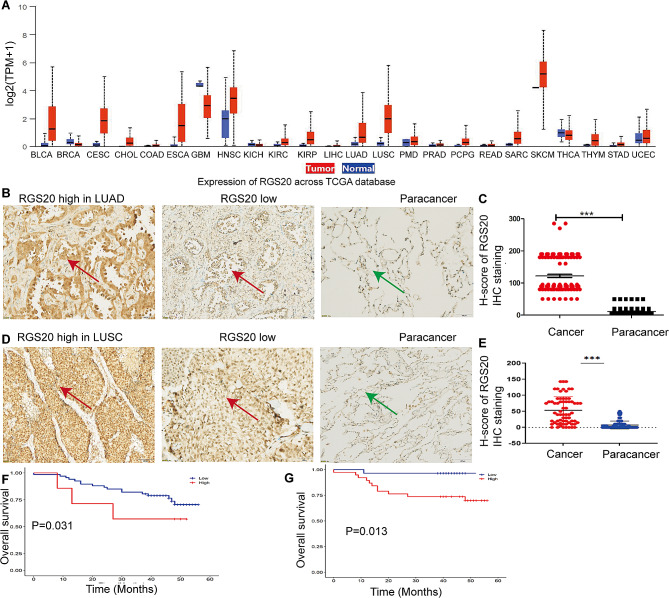 Figure 1
Figure 1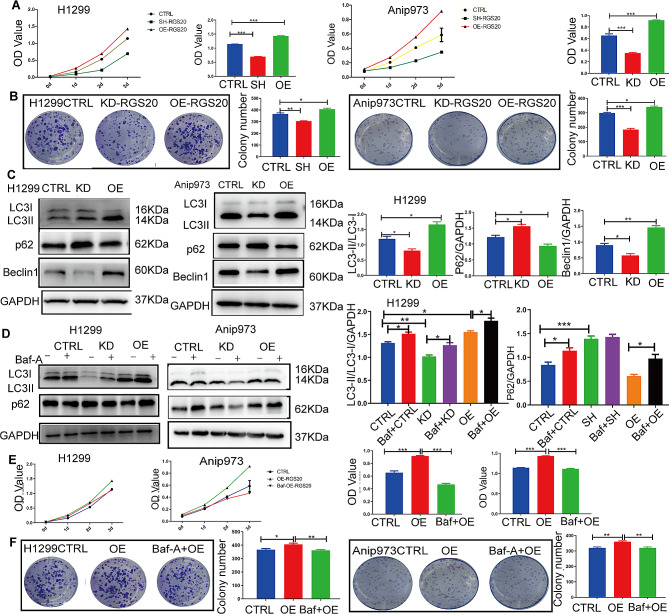 Figure 2
Figure 2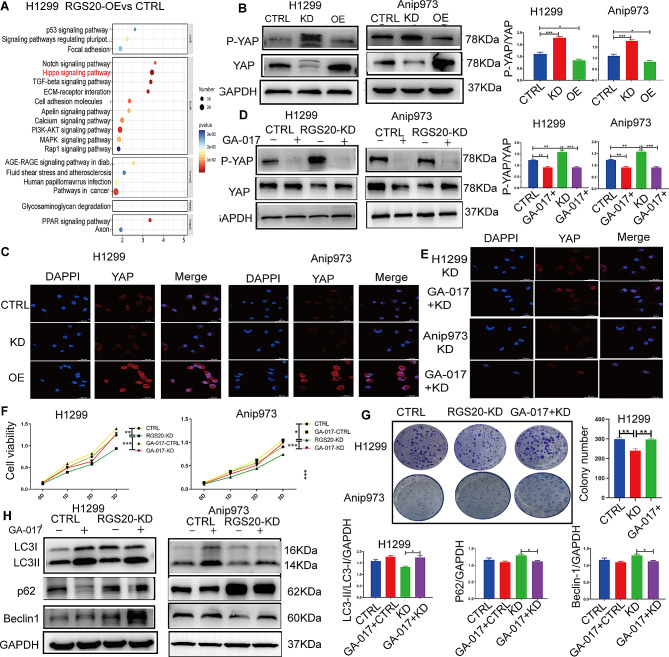 Figure 3
Figure 3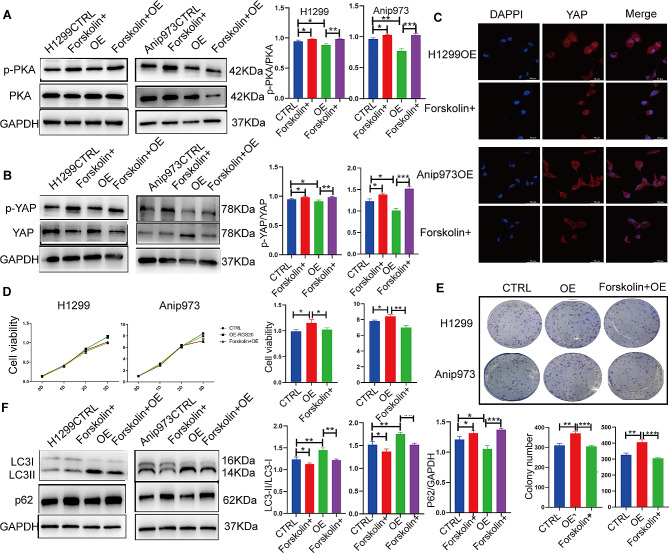 Figure 4
Figure 4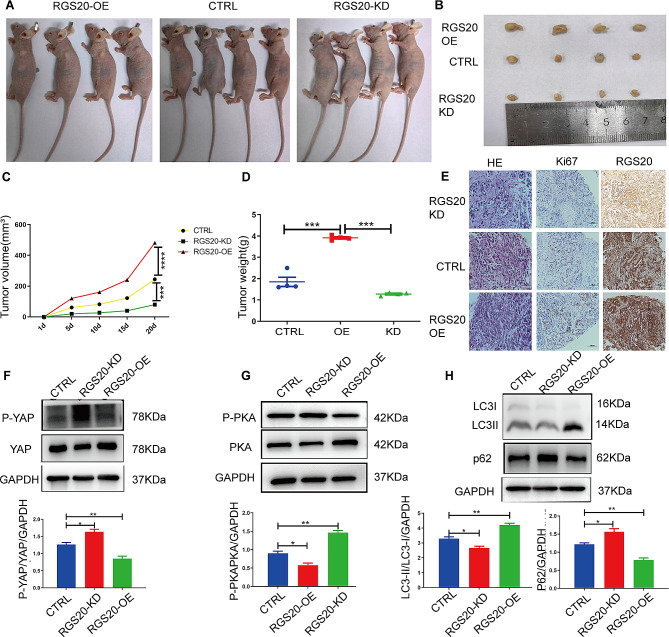 Figure 5
Figure 5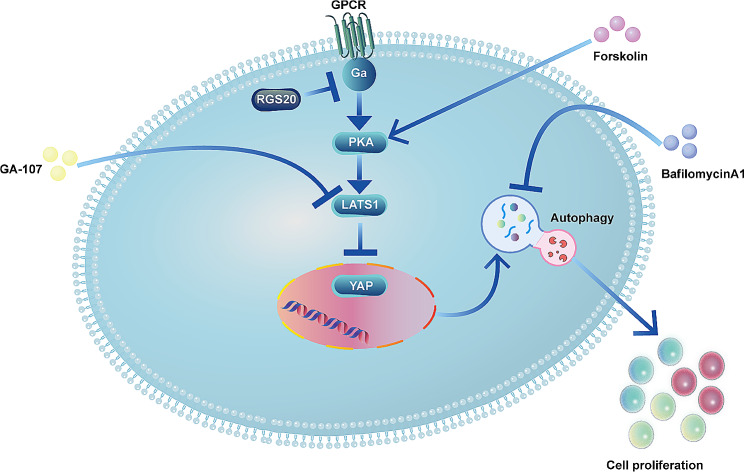 Figure 6
Figure 6- —http://dx.doi.org/10.13039/501100014761Natural Science Foundation of Qingdao Municipality
- —http://dx.doi.org/10.13039/501100001809National Natural Science Foundation of China
- —Chinese Academy of Medical Sciences Innovation Fund for Medical Sciences
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsRadioactive element chemistry and processing · Nuclear materials and radiation effects · Chemical Synthesis and Characterization
Introduction
Non-small cell lung cancer (NSCLC) is the leading cause of cancer-related deaths worldwide [1] and include adenocarcinoma, squamous cell carcinoma, and large cell carcinoma [2]. With the advancement of molecular-targeted therapies, patients with NSCLC have seen significant treatment improvements over the past decade [3–5]. Further, inhibitors of kinases, such as epidermal growth factor receptor (EGFR) signaling [6], are in routine clinical use. However, resistance to current treatments often emerges within 2–3 months from the start of therapy in the majority of patients with advanced NSCLC [7]. Therefore, the discovery of novel biomarkers associated with NSCLC development has emerged as a prominent area of research, especially for developing more targeted and personalized treatment approaches.
Regulators of G protein signaling (RGS) proteins exert significant influence on the functions of G protein-coupled receptors (GPCRs) [8]. Among mammalian RGS proteins, RGS17 binds to and activates the GTPase activity of G (i/o), G (z), and G (q), thereby inhibiting GPCR signaling [9]. Another member, RGS20, shares about 62% similarity with RGS17 [10] and has been implicated in the development of various malignant tumors. For example, it is linked to the proliferation and migration of penile cancer [9], bladder cancer [11] and oral squamous cell carcinoma [12]. Further, high RGS20 expression correlates significantly with the progression and prognosis of triple-negative breast cancer and renal cancer [13, 14]. Considering the involvement of RGS20 in various cancers, it is surprising that the role of RGS20 in NSCLC remains largely unexplored. A recent study reported that RGS20 promotes cell aggregation and migration [10]. However, functional and prognostic analyses, as well as the molecular mechanisms of RGS20 in NSCLC still remain poorly understood.
In this study, we first examined the expression of RGS20 in clinical tumor samples of lung adenocarcinoma and squamous cell carcinoma, and explored the relationship between its expression level and prognosis. Next, we investigated the effects of overexpression or knockdown of RGS20 on cell proliferation and autophagy in NSCLC cells. Furthermore, transcriptomic sequencing was performed to find the key pathways involved in the role of RGS20. At last, the xenograft nude mouse model was used to confirm the role of RGS20 in NSCLC in vivo.
Materials and methods
Patient specimens and tissue microarrays
A total of 126 pairs of paraffin sections of lung adenocarcinoma and corresponding normal lung tissues were obtained from the Affiliated Hospital of Qingdao University, China. All samples were sequenced by NGS to exclude egfr, alk, ros1, braf, her-2, kras, nras, pik3ca, ret or met gene mutations. A human lung squamous carcinoma tissue microarray consisting of 75 pairs of primary lesions with matched normal adjacent control tissues (HLug-Squ150Sur-02) was purchased from Xinchao Biotechnology Co., Ltd. (Shanghai, China). Patient consent and ethical approval were obtained from the Institutional Research Ethics Committee (Ethical approval number: QYFY WZLL 27,991, SHYJS-CP-1,904,009).
Cell lines and reagents
NSCLC cell lines (H1299, Anip973) were maintained in RPMI Medium 1640 (Gibco, Waltham, MA, USA) supplemented with 1% penicillin-streptomycin (NCM Biotech, Suzhou, China) and 10% heat-inactivated fetal bovine serum (Gibco, Waltham, MA, USA). All cells were kept at 37 °C in a humidified atmosphere of 5% CO_2_. Drug treatments were performed directly on cultured media. Bafilomycin A1 (BafA1; 50 nM), GA-017 (10 μM), and Forskolin (10 μM) were all purchased from GLPBIO (Montclair, America).
Lentivirus construction and transfection
The lentiviruses used in this study were constructed and supplied by Genomeditech Co., Ltd. (Shanghai, China). The shRNA sequences were specifically designed to target human rgs20. The three shRNA sequences used are listed in Supplementary Table 1. Following the manufacturer’s instructions, lentiviruses were transfected into the cells using the transfection reagent for 48 h. Transfection efficiency was assessed by western blot analysis.
Immunoblotting
Proteins were extracted and detected as previously described [16]. Briefly, SDS-PAGE (Epizyme, Shanghai, China) was used to separate proteins and then transferred onto PVDF membranes (Millipore, Germany). Membrane blocking was performed by using a 5% dry milk solution dissolved in Tris Buffer Saline with 1% Tween 20 (TBST) and incubated overnight at 4 °C with the primary antibody. A secondary antibody was used, and immunoblots were developed using an ECL western substrate (Epizyme, Shanghai, China). The bands were visualized using a gel imaging system (Tanon 5200, Shanghai, China), and ImageJ software was used to quantify the integrated optical density of the bands. The antibodies used in this study are listed in Supplementary Table 2.
CCK-8 assay
Cells were seeded in 96-well plates at a density of 1,000 cells per well and assessed for proliferation using Cell Counting Kit-8 (CCK-8) (GK10001, Biosharp, Beijing, China). After incubation for 24, 48, 72, and 96 h, the culture medium was replaced with 100 μl of 10% CCK-8 serum-free medium. Following an additional hour in a cell incubator, the absorbance of cells was measured at 450 nm.
Tumor colony formation assay
A single-cell suspension containing 5 × 10^3^ cells was cultured in RPMI Medium 1640 (10% FBS) for 10 days in a six-well plate. Once most of the cell clones exceeded the 50-cell mark, the cells were washed twice with PBS, treated with 4% paraformaldehyde for 10 min, and stained with 0.05% crystal violet for 15 min at room temperature. After rinsing off the stain, the number of colonies (50 cells/colony) was counted under a microscope at 40× magnification (Olympus, Japan).
Transcriptome sequencing
TRIzol reagent was employed to extract total RNA from both the control group and the RGS20-OE group of lung adenocarcinoma H1299 cells, with three replicates per group. The transcriptome sequencing and subsequent analysis of the sequencing results were outsourced to OE Biotech Co. Ltd. in Shanghai, China. The libraries were subjected to sequencing on an Illumina HiSeq X Ten platform, generating 150 bp paired-end reads for subsequent analysis of differentially expressed genes.
Immunofluorescence
Cells were initially seeded on 12 mm coverslips in 24-well plates. After 24 h of culture, cells underwent fixation with 4% paraformaldehyde and were permeabilized in 0.1% Triton X-100/PBS. Subsequently, cells were blocked for 2 h in 5% bovine serum albumin/PBS, followed by overnight incubation at 4 °C with primary antibody at a dilution of 1:100. Afterward, fluorescence-labeled secondary antibodies were applied for 2 h. Nuclei were strained using DAPI. Imaging was performed using a confocal laser scanning microscope at 400× magnification (Leica STELLARIS 5, Germany). The antibodies used in this study are listed in Supplementary Table 2.
Xenograft nude mice model
Four weeks old female BALB/c nude mice were purchased from the Beijing Vital River Laboratory (Beijing, China), bred, and housed in a specific pathogen-free (SPF) environment. All animal experiments adhered to the ethical guidelines of the Institutional Ethics Committee of Qingdao University, China (Ethical approval number: 20221117BALB/cnude2420221230061). The antibodies used in this study are listed in Supplementary Table 2.
Statistical analysis
The data analysis was performed using Prism 5.0 (Graph Pad Software, Inc., La Jolla, CA, USA). A student’s T-test or one-way ANOVA, was employed to analyze differences between groups, including the expression of clinicopathological parameters and immunohistochemical indices. The results are presented as the mean ± SD of three or more observations in each experiment, and a significance level of p < 0.05 was considered statistically significant.
Results
Elevated expression of RGS20 is associated with a poor prognosis of NSCLC
We investigated rgs20 expression profiles across multiple tumors using the Cancer Genome Atlas (TCGA) database (https://ualcan.path.uab.edu/cgi-bin/Pan-cancer.pl?genenam=RGS20). Elevated rgs20 expression was noticed in lung adenocarcinoma (LUAD), lung squamous cell carcinoma (LUSC), and other tumors (Fig. 1A). Specifically, RGS20 was more highly expressed in adenocarcinomas than in adjacent noncancerous tissues (Fig. 1B and C) and exhibited markedly higher expression in squamous lung tissue than that of normal lung tissue (Fig. 1D and E). Furthermore, a positive correlation was observed between RGS20 expression levels and the T stage of the tumor as well as cervical lymph node metastasis (N stage) (Table 1). Kaplan-Meier analysis demonstrated that patients with high RGS20 expression levels exhibited significantly shorter five-year overall survival rates in both LUAD (Fig. 1F) and LUSC (Fig. 1G). Multivariate Cox regression analysis indicated that RGS20 expression was an independent prognostic factor for LUSC (Table 2). Taken together, these findings suggest RGS20 as a potential biomarker for NSCLC diagnosis, with elevated expression levels positively correlated with a poor prognosis.
Fig. 1. Elevated expression of RGS20 is associated with a poor prognosis for NSCLC A The mRNA expression of rgs20 in many cancers was assessed using TCGA database. B The representative IHC staining of the RGS20 protein in lung adenocarcinoma tumors (red arrow) and adjacent non-cancerous tissues (green arrow). C The quantitative analysis of IHC staining results for RGS20 in 126 LUAD tissues. D The representative IHC staining of RGS20 protein in lung squamous carcinoma tissue microarray (red arrow) and adjacent non-cancerous tissues (green arrow). E The quantitative analysis of IHC staining results for RGS20 in 75 LUSC tissues. Kaplan-Meier analysis of the association between RGS20 protein levels and overall survival in 126 LUAD (F) and 75 LUSC (G) patients. The results in C and E were quantified with CaseViewer (version 2.3) and presented as H-Scores. The samples from patients were divided into high and low subgroups based on the median H-score of IHC staining, which represents the expression level of RGS20. * p < 0.05, **p < 0.01, ***p < 0.001
Table 1. Correlation between RGS20 expression and clinicopathological characteristics in lung adenocarcinomavariablesRGS20 expressiontotalχ^2^p valuelowhighAge (year)0.1520.697≤ 60372560> 60362866Sex0.0210.886male282149Female453277T stage5.6900.029^^T1-T26339102T3/T4101424N stage10.2420.002^^N07434108Nx-N281018TNM stage2.7990.175I-II7145116III5611Tumor rize0.8330.361≤ 2 cm503787> 2 cm192039* Statistically significant (p < 0.05)
Table 2. Univariate and multivariate analyses of the factors correlated with Overall survival of LUSC patients Variables in the EquationvariablesUnivariate analysisMultivariate analysisp valueHR95%CIp valueHR95%CILowerlimitUpperlimitLowerlimitUpperlimitexpression0.0408.5731.10466.5490.018^^12.0171.52694.617Age0.0582.9870.9629.270sex0.4940.0430.000346.179Tumor size0.4960.6340.1702.357Grade stage0.1890.0290.0005.700TNM stage0.1732.1950.7086.808T stage0.0134.2141.35513.1000.002^^6.0611.92919.041N stage0.3441.7280.5575.357LUSC: lung squamous cell carcinoma, HR: Hazard Ratio, CI: Confidence Interval,* Statistically significant (p < 0.05)
RGS20 promotes NSCLC cell proliferation by activating autophagy in vitro
The role of RGS20 in NSCLC was further examined using H1299 and Anip973 cells (Supplementary Fig. 1A). Next, RGS20 overexpression (RGS20-OE) and knockdown (RGS20-KD) cell lines were generated using lentiviral vectors (Supplementary Fig. 1B, C, D). RGS20 enhanced cell proliferation and foci formation (Fig. 2A and B), consistent with previous reports [11]. Since autophagy plays a critical role in various stages of tumor progression [15], we investigated whether RGS20 promotes cell proliferation by increasing autophagy in NSCLC cells. RGS20 overexpression increased the ratio of LC3II/LC3I protein levels, elevated BECLIN1 expression, and reduced P62 protein levels, indicating increased autophagy (Fig. 2C). To further investigate the potential link between autophagy and proliferation in RGS20-OE cells, Bafilomycin A1 (Baf-A1) was used to block autophagic flux. The observed accumulation of LC3-II and P62 suggested that Baf-A1 effectively blocked the autophagic flux (Fig. 2D). Notably, suppression of autophagic flux resulted in a decrease in the proliferation of RGS20-OE cells (Fig. 2E and F). Taken together, these findings suggest that RGS20 induces NSCLC cell proliferation by upregulating autophagy in vitro.
Fig. 2RGS20 promotes NSCLC cell proliferation and activates autophagy in vitro A Cell proliferation of three different groups (CTRL, RGS20-KD, and RGS20-OE) in H1299 and Anip973 cells was detected by the Cell Counting Kit-8 (CCK-8) assay. B Colony formation assays show the colony-forming ability of H1299 and Anip973 cells in three groups (CTRL, RGS20-KD, and RGS20-OE). The number of colonies was counted and is displayed on the right side. C Representative western blot results show the protein levels of autophagy-related proteins and the corresponding quantification analysis. D Representative western blot results show the protein levels of autophagy related proteins after blockage of autophagy flux by Baf-A1 (50 nM, 2 h) in three groups (CTRL, RGS20-KD, and RGS20-OE) of H1299 and Anip973 cells. The corresponding quantification analysis results were listed on the right. E CCK-8 assay was performed to assess the proliferation of H1299 and Anip973 cells in the RGS20-OE group treated with Baf-A1 (50 nM, 2 h). F Colony formation assays were used to analyze the colony-forming ability of H1299 and Anip973 cells in the RGS20-OE group treated with Baf-A1 (50 nM, 2 h). The number of colonies was counted and is displayed on the right side. * p < 0.05, **p < 0.01, ***p < 0.001
RGS20 inhibites the Hippo signaling pathway in NSCLC
Transcriptome sequencing was performed to explore the specific molecular mechanisms of action of RGS20 in NSCLC. The Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analysis revealed 20 signaling pathways. Notably, differentially expressed genes were predominantly enriched in the Hippo signaling pathway (Fig. 3A). Therefore, we hypothesized that RGS20 promotes NSCLC cell proliferation and autophagy via the Hippo signaling pathway. Indeed, phosphorylated YAP (p-YAP) levels were decreased in RGS20-OE cells and increased in RGS20-KD cells (Fig. 3B). Furthermore, RGS20-KD cells exhibited reduced YAP nuclear translocation, whereas RGS20-OE cells demonstrated increased nuclear translocation of YAP (Fig. 3C). To examine the potential association between RGS20 and the Hippo signaling pathway in NSCLC, RGS20-KD cells were treated with GA-017, a specific inhibitor of the Hippo signaling pathway. GA-017 treatment led to a decrease in p-YAP protein levels (Fig. 3D) and an increase in nuclear translocation of YAP (Fig. 3E), confirming the inhibition of the Hippo signaling pathway in RGS20-KD cells. In addition, CCK8 and clone formation assays revealed accelerated cell proliferation and foci formation in RGS20-KD cells treated with GA-017 (Fig. 3F and G). These findings suggest that inhibition of the Hippo signaling pathway in RGS20-KD cells promotes cell growth and clonogenic potential. Moreover, the levels of autophagy-related proteins, such as LC3-II/LC3-I and BECLIN-1, were upregulated in RGS20-KD cells following inhibition of the Hippo signaling pathway (Fig. 3H). These results suggest that RGS20 exerts an inhibitory effect on the Hippo signaling pathway in NSCLC, potentially affecting autophagy regulation.
Fig. 3RGS20 inhibits the Hippo signaling pathway in NSCLC A KEGG pathway analysis was performed by transcriptome sequencing to identify the signaling pathway in which differentially expressed genes were enriched. B The representative Western blot result shows the protein levels of p-YAP and YAP in the control, RGS20-KD, and RGS20-OE groups of H1299 and Anip973 cells. The corresponding quantification analysis results are shown on the right. C Representative immunofluorescence images show the nuclear translocation of YAP in three different groups of H1299 and Anip973 cells. D The representative Western blot result shows the p-YAP and YAP protein levels after the blockage of the Hippo signaling pathway with GA-017 (10 μM, 24 h) in the RGS20-KD and control groups. The corresponding quantification analysis results are shown on the right. E Representative immunofluorescence images of YAP in the RGS20-KD group after treatment with GA-017. F CCK-8 assay to detect the proliferation of H1299 and Anip973 cells after treatment with GA-017 (10 μM, 72 h) in the control and RGS20-KD groups. G Colony formation assays were used to analyze the colony-forming ability of H1299 and Anip973 cells treated with GA-017 (10 μM, 72 h). H Representative Western blot result shows autophagy-related protein levels after the blockage of the Hippo signaling pathway with GA-017 (10 μM, 24 h) in the RGS20-KD and control groups. * p < 0.05, **p < 0.01, ***p < 0.001
RGS20 promotes cell proliferation and enhances autophagy by suppressing the PKA-Hippo signaling pathway in NSCLC
Because GPCRs represent a significant signaling branch upstream of the Hippo pathway [16], RGS proteins may modulate cell proliferation and autophagy by regulating the PKA-mediated GPCR signaling pathway. Consistently, the phosphorylation of PKA was reduced in RGS20-OE cells and increased in RGS20-KD cells (Fig. 4A). To investigate the relationship between PKA and the Hippo signaling pathway, RGS20-OE cells were treated with forskolin, a cyclic adenosine monophosphate (cAMP) activator. Subsequently, an elevated level of p-YAP compared to that of total YAP (Fig. 4B) and a decrease in the translocation of YAP to the nucleus (Fig. 4C) were observed. In addition, CCK-8 and clone formation assays revealed decreased cell proliferation and foci formation in RGS20-OE cells following forskolin administration (Fig. 4D and E). Furthermore, the LC3-II/LC3-I ratio decreased in RGS20-OE cells treated with forskolin, indicating a reduction in autophagy. Meanwhile, the protein level of P62 was increased, suggesting impaired autophagy (Fig. 4F). These findings demonstrate that RGS20 promotes cell proliferation and enhances autophagy by suppressing the PKA-Hippo signaling pathway in NSCLC.
Fig. 4RGS20 promotes cell proliferation and enhances autophagy by inhibiting the PKA-Hippo signaling pathway in vitro A Representative western blot result shows the protein level of p-PKA and PKA in RGS20-OE cells after treatment with forskolin (10 μM, 72 h). The corresponding results of the quantification analysis are displayed on the right. B Representative western blot result shows the protein level of p-YAP and YAP after the activation of the PKA signaling pathway with forskolin (10 μM, 72 h) in RGS20-OE and control cells. C Representative immunofluorescence images of YAP in the RGS20-OE group after treatment with forskolin. D The CCK-8 assay detects the proliferation of H1299 and Anip973 cells after treatment with forskolin (10 μM, 72 h) in the RGS20-OE and control groups. E Colony formation assays were used to analyze the colony-forming ability of H1299 and Anip973 cells after treatment with forskolin (10 μM, 72 h). F Representative western blot result shows the protein levels of autophagy-related proteins after the activation of the PKA signaling pathway with forskolin (10 μM, 72 h) in the RGS20-OE and control groups. * p < 0.05, **p < 0.01, ***p < 0.001
RGS20 enhances the tumorigenesis of NSCLC cells in vivo
To further assess the tumorigenic effects of RGS20 on NSCLC cells in vivo, we established a xenograft nude mouse model. The growth curve of xenograft tumors showed that RGS20 promoted tumorigenesis in vivo (Fig. 5A and B). Xenograft tumors in the RGS20-OE group were larger and heavier than those in the control group. We observed significant inhibition of tumor growth in the RGS20-KD group (Fig. 5C and D). Hematoxylin and eosin (HE) staining showed that all tumors were solid, with a higher Ki-67 index in tumors of the RGS20-OE group than in those of the control group (Fig. 5E). To validate the mechanisms by which RGS20 promotes the proliferation of NSCLC cells in vivo, we measured protein levels linked to the PKA-Hippo signaling pathway and autophagy in solid tumors. We noticed a significant decrease in the ratio of p-PKA/PKA and p-YAP/YAP protein levels in the tumor tissues of the RGS20-OE group compared to that of the control group (Fig. 5F and G). Further, the ratio of LC3-II/LC3-I protein levels significantly rose in the tumors of the RGS20-OE group compared to that in the control group, whereas P62 protein levels showed the opposite trend (Fig. 5H). Therefore, RGS20 enhanced the tumorigenesis of NSCLC cells in vivo by increasing autophagy via the suppression of the PKA-Hippo signaling pathway.
Fig. 5RGS20 enhances the tumorigenesis of NSCLC cells in vivo A Tumor-promoting effects of RGS20 in a xenograft mouse model. The representative images of tumor-bearing mice that were photographed at the end of the third week of experimentation. B Representative images of the dissected tumors. A ruler was used to indicate the size of the tumors. C The tumor growth curve was plotted using xenograft tumor volume data. D Tumor weight was measured after tumor excision. E Representative images of HE staining and IHC staining of Ki-67 and RGS20. Representative western blot results show the protein level of the PKA-Hippo (F, G) signaling pathway and autophagy-related proteins (H). The corresponding quantification analysis results are listed below. Protein levels were normalized to GAPDH. Results are presented as the mean ± SD (n = 3). * p < 0.05, **p < 0.01, ***p < 0.001
Discussion
In this study, we observed elevated RGS20 expression in NSCLC tumors, correlating with a poor prognosis. Additionally, high RGS20 levels were significantly related to advanced T stage, lymphatic metastasis, and poorer 5-year overall survival rates. Multivariate Cox regression analysis confirmed RGS20 expression as an independent prognostic factor. These findings suggest that RGS20 is likely an oncogene and can serve as a novel diagnostic and prognostic marker for NSCLC.
RGS proteins negatively regulate G-protein signaling by binding to G-protein α subunits and facilitating GTP hydrolysis, thereby dampening signal transduction [17]. A previous study assessing the global map of GPCR by RGS proteins reported that RGS A/RZ subfamily members (RGS17, RGS19, and RGS20) uniquely regulate Gαz, Gαi/o, and Gαq proteins [18]. RGS17 has been reported to induce lung and prostate cancer proliferation via the cAMP-PKA-CREB pathway [19]. Though a few other studies have explored the relationship between the cAMP-PKA pathway and pathogenic lung inflammation [20] or injury [21], limited research exists on the role of this pathway in NSCLC. Though a few exceptions exist, such as a study that revealed that endogenous glutamate regulates ferroptosis sensitivity in LUAD through PKA signaling [22], a second study demonstrated the involvement of MHY4571 in the regulation of cAMP-PKA signaling in squamous cell lung cancer treatment [23]. Our study highlighted RGS20’s inhibition of cAMP-PKA signaling, which was reversed by forskolin in RGS20-OE NSCLC cells.
As a downstream branch of GPCR signaling, the Hippo pathway plays a crucial role in regulating cell survival [16], proliferation [24], differentiation and organ size [25, 26]. The core Hippo signaling pathway in mammals includes mammalian sterile 20-like kinase 1/2 (MST1/2), large tumor suppressor kinase 1/2 (LATS1/2), and YAP/transcriptional coactivator with a PDZ-binding motif (TAZ) [27]. Phosphorylation of YAP/TAZ leads to its cytoplasmic retention and ubiquitination-dependent proteasomal degradation. When upstream kinases are inactivated, dephosphorylated YAP/TAZ translocates to the nucleus and induces the expression of target genes [16, 27]. Elevated expression and nuclear localization of YAP have been observed in many cancers, including cervical squamous cell carcinoma, colorectal cancer, and esophageal carcinoma [28–30] and have also been associated with poor prognosis in esophageal carcinoma and ovarian cancer [30, 31]. Elevated YAP levels have also been detected in lung cancer patients with acquired EGFR inhibitor resistance [32] and TAZ level was a prognostic factor in NSCLC progression [33, 34]. Our study demonstrated that overexpression of RGS20 decreased the expression of p-YAP and increased YAP translocation to the nucleus, which further verified the important correlation between GPCR and the Hippo pathway.
The Hippo signaling pathway also acts as an upstream regulator of autophagy [35, 36]. Autophagy plays a dual role in tumors, acting as a double-edged sword by both promoting tumor growth and inhibiting tumor progression. For example, loss of autophagy results in the incidence of hepatocellular carcinoma [37]. Further, autophagy has been reported to suppress breast cancer metastasis by degrading the neighbor of BRCA1 gene 1 (NBR1) [38, 39]. However, autophagy can also promote pancreatic cancer [40]. The potential ability of autophagy to modulate cell death and regulate autophagy has made it an effective interventional strategy for cancer therapy [41–43]. Lung cancer has high basal autophagic activity, which is associated with autophagy during tumorigenesis [44]. A recent study revealed that sex-determining region Y-box 2 (SOX2) promotes LC3A expression and enhances the proliferation of lung cancer cells. Autophagy is activated in serine/threonine kinase 11 (STK11) mutant lung cancer [45]. Our study found that RGS20 overexpression enhanced autophagy by increasing the expression of the proteins LC3 II and BECLIN1 while decreasing the protein level of P62. Further, our in vitro experiments demonstrated that the viability of RGS20-OE cells was decreased by blocking autophagy. Furthermore, we noticed that the tumor-promoting activity of RGS20 relies on autophagy. Our findings, along with previous research, point to the crucial role played by autophagy in various cancer types. However, the specific molecular mechanisms by which RGS20 influences autophagy remain unclear and require further exploration by future studies.
In conclusion, our study suggests that elevated RGS20 expression in NSCLC is associated with a poor five-year survival rate. RGS20 promotes NSCLC cell proliferation by enhancing autophagy via suppression of the PKA-Hippo signaling pathway (Fig. 6). Further research on RGS20 may offer novel and promising therapeutic targets, particularly for the treatment of patients with drug-resistant NSCLC.
Fig. 6. Graphical Abstract RGS20 promotes NSCLC proliferation and activates autophagy by inhibiting the PKA-Hippo signaling pathway
However, there are still gaps in our understanding of the role of RGS20 in NSCLC. For example, the specific mechanisms by which RGS20 activates autophagy, its impact on other tumor behaviors such as invasion, metastasis, and metabolism, and the existence of small molecule inhibitors targeting RGS20, all require further experimental investigations to be addressed in future.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Supplementary Material 1
Supplementary Material 2
Supplementary Material 3
Supplementary Material 4
Supplementary Material 5
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Chen Z Fillmore CM Hammerman PS Kim CF Wong KK Non-small-cell lung cancers: a heterogeneous set of diseases Nat Rev Cancer 20141485354610.1038/nrc 377525056707 PMC 5712844 · doi ↗ · pubmed ↗
- 2Siegel RL Miller KD Jemal A Cancer statistics, 2016 CA Cancer J Clin 201666173010.3322/caac.2133226742998 · doi ↗ · pubmed ↗
- 3Guo H, Zhang J, Qin C, Yan H, Liu T, Hu H et al. Biomarker-targeted therapies in non-small cell lung cancer: current status and perspectives. Cells. 2022;11(20).10.3390/cells 11203200 PMC 960044736291069 · doi ↗ · pubmed ↗
- 4Chen R Manochakian R James L Azzouqa AG Shi H Zhang Y Emerging therapeutic agents for advanced non-small cell lung cancer J Hematol Oncol 20201315810.1186/s 13045-020-00881-732448366 PMC 7245927 · doi ↗ · pubmed ↗
- 5Wang M Herbst RS Boshoff C Toward personalized treatment approaches for non-small-cell lung cancer Nat Med 202127813455610.1038/s 41591-021-01450-234385702 · doi ↗ · pubmed ↗
- 6Hsu WH Yang JC Mok TS Loong HH Overview of current systemic management of EGFR-mutant NSCLC Annals Oncology: Official J Eur Soc Med Oncol 201829 suppl 1i 3i 910.1093/annonc/mdx 70229462253 · doi ↗ · pubmed ↗
- 7Rotow J Bivona TG Understanding and targeting resistance mechanisms in NSCLC Nat Rev Cancer 201717116375810.1038/nrc.2017.8429068003 · doi ↗ · pubmed ↗
- 8Alqinyah M Hooks SB Regulating the regulators: epigenetic, transcriptional, and post-translational regulation of RGS proteins Cell Signal 201842778710.1016/j.cellsig.2017.10.00729042285 PMC 5732043 · doi ↗ · pubmed ↗