Robust Immune Response and Protection against Lethal Pneumococcal Challenge with a Recombinant BCG-PspA-PdT Prime/Boost Scheme Administered to Neonatal Mice
Monalisa Martins Trentini, Dunia Rodriguez, Alex Issamu Kanno, Cibelly Goulart, Michelle Darrieux, Luciana Cezar de Cerqueira Leite

TL;DR
A new vaccine strategy using a recombinant BCG and protein boost protects neonatal mice from deadly pneumococcal infection.
Contribution
A prime/boost vaccine using rBCG-PspA-PdT and rPspA-PdT provides robust neonatal protection against pneumococcal infection.
Findings
The prime/boost strategy induced an IgG1 to IgG2c isotype shift and increased memory cells and cytokine production.
100% protection was observed in neonates against a lethal pneumococcal challenge with the WU2 strain.
Two doses of rPspA-PdT alone provided non-significant protection in neonates.
Abstract
Pneumococcal diseases are an important public health problem, with high mortality rates in young children. Although conjugated pneumococcal vaccines offer high protection against invasive pneumococcal diseases, this is restricted to vaccine serotypes, leading to serotype replacement. Furthermore, the current vaccines do not protect neonates. Therefore, several protein-based pneumococcal vaccines have been studied over the last few decades. Our group established a recombinant BCG expressing rPspA-PdT as a prime/rPspA-PdT boost strategy, which protected adult mice against lethal intranasal pneumococcal challenge. Here, we immunized groups of neonate C57/Bl6 mice (6–10) (at 5 days) with rBCG PspA-PdT and a boost with rPspA-PdT (at 12 days). Controls were saline or each antigen alone. The prime/boost strategy promoted an IgG1 to IgG2c isotype shift compared to protein alone. Furthermore,…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
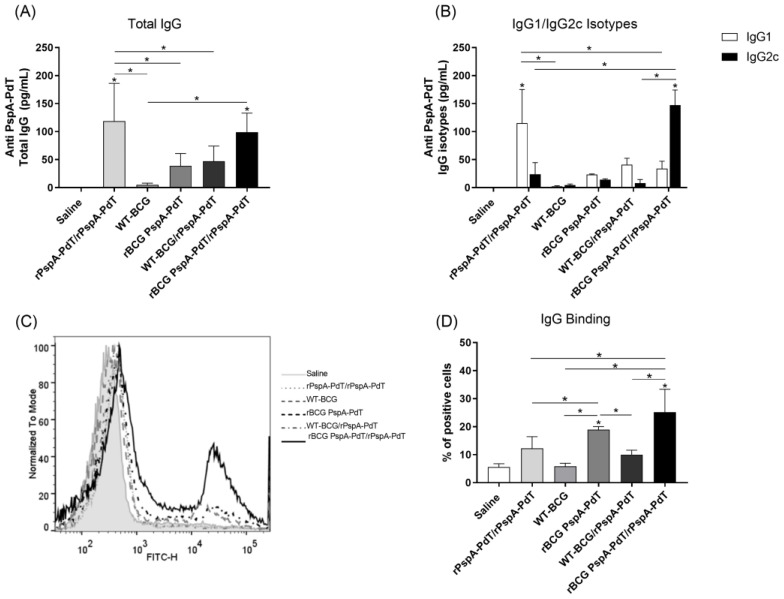 Figure 1
Figure 1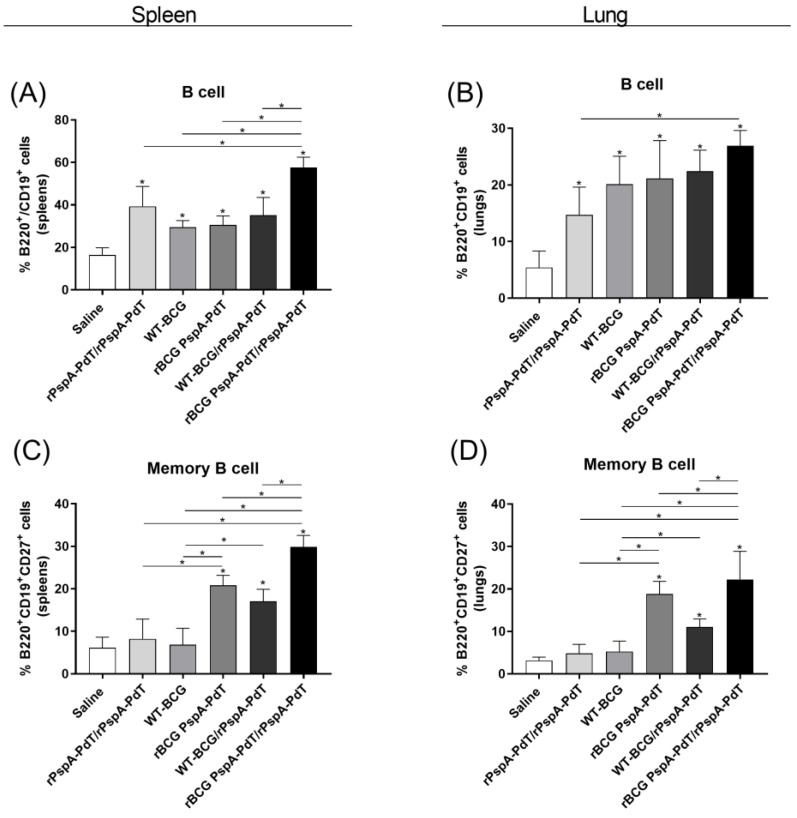 Figure 2
Figure 2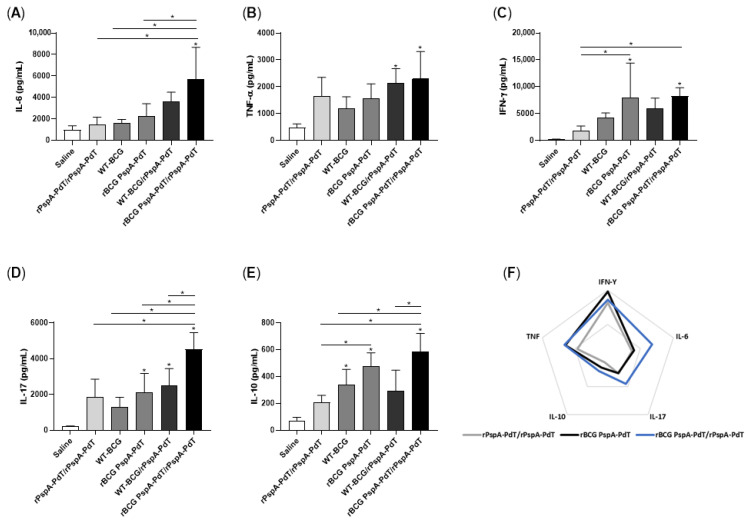 Figure 3
Figure 3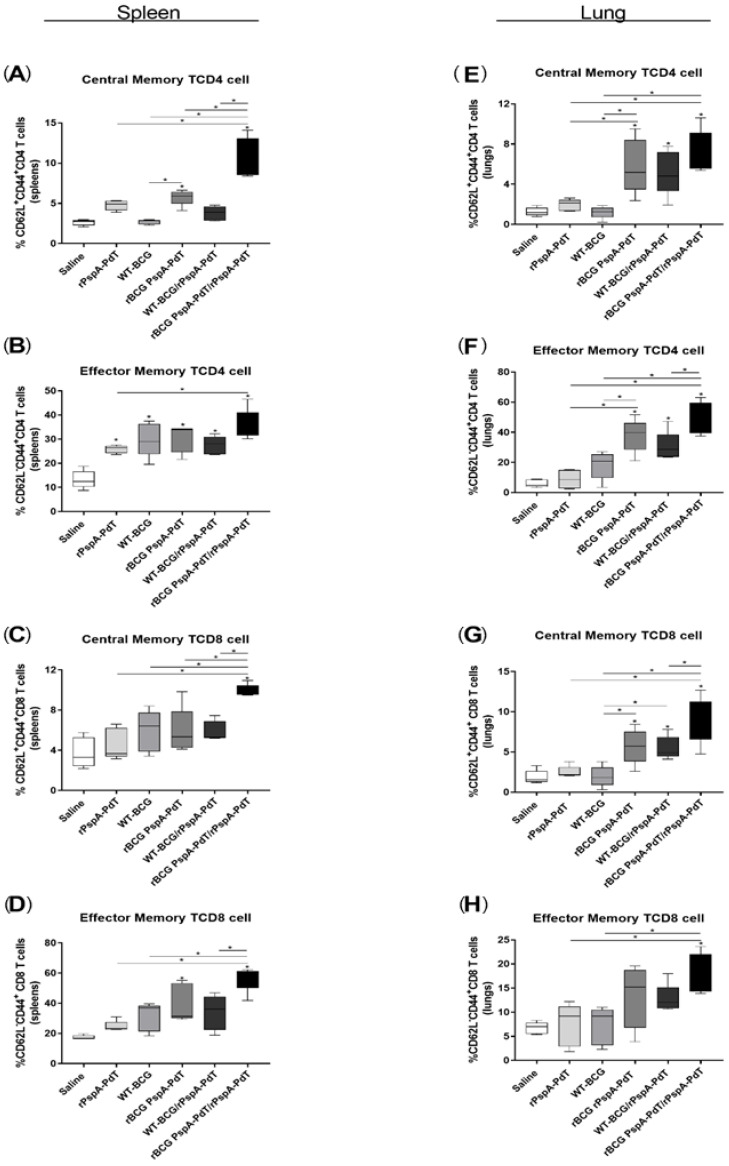 Figure 4
Figure 4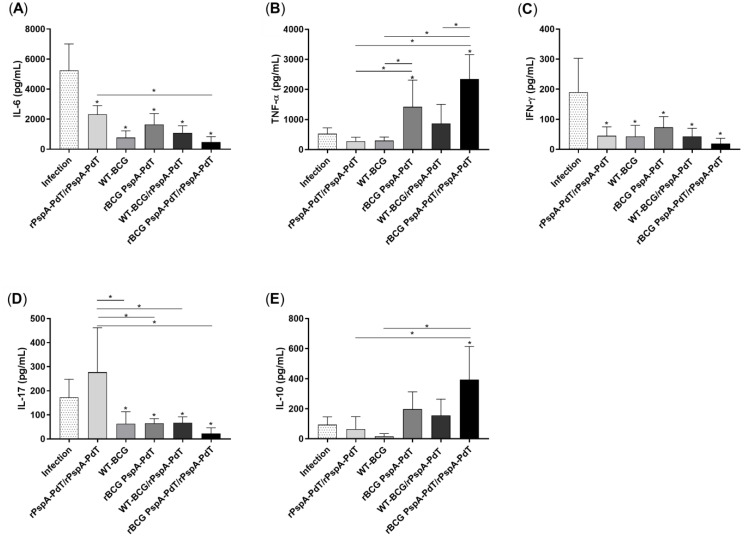 Figure 5
Figure 5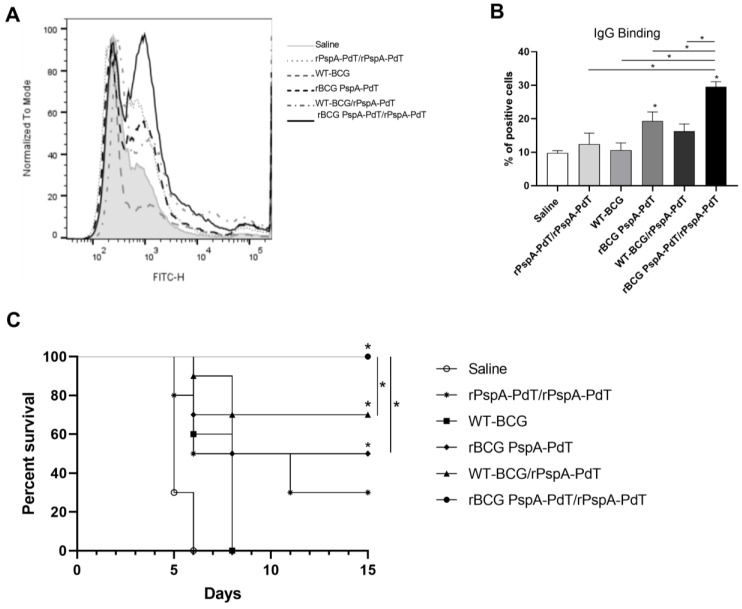 Figure 6
Figure 6- —FUNDAÇÃO DE AMPARO À PESQUISA DO ESTADO DE SÃO PAULO (FAPESP)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPneumonia and Respiratory Infections · Respiratory viral infections research · Influenza Virus Research Studies
1. Introduction
Pneumococcal infections remain a significant global public health problem worldwide, responsible for more than 740,000 deaths among children under 5 years of age in developing countries [1,2]. Pneumococcal conjugate vaccines are licensed for use in children, providing protection against up to 13 prevalent pneumococcal serotypes (PCV7, PCV10, and PCV13) [3,4,5,6]. While PCVs have a very low failure rate in children from industrialized countries with effective national immunization programs, the limited vaccination coverage in many parts of the world, replacement by non-vaccine serotypes, and increasing resistance to antibiotics greatly impact the efficacy of these formulations in low- and middle-income countries (LMIC), reinforcing the need to develop alternative vaccines [7,8]. The highly complex production process for PCVs leads to high production costs and limits their production in LMIC. Therefore, an ideal pneumococcal vaccine should induce broad cross reactivity (serotype-independent) and have low production costs [9,10,11].
Many research investigations have shown the promise of pneumococcal proteins as possible alternatives for developing serotype-independent vaccines against pneumococcal disease [12,13,14,15]. Pneumococcal surface protein A (PspA) has been extensively investigated and considered a promising vaccine candidate, displaying high immunogenicity and protective efficacy in different infection models [16,17,18,19]. PspA is an exposed virulence component that generates antibodies with the capacity to increase complement activation, C3 deposition, and killing via opsonophagocytosis [20,21,22,23]. PspA has also been shown to protect the bacterium from antimicrobial peptides derived from lactoferrin [24]. PspA has been shown to display some degree of variability, classified into three families comprising five clades [13,25,26]. We previously determined that most isolated strains in Brazil are distributed between families 1 (comprising clades 1 and 2) and 2 (clades 3 and 4) [27]; this is also true for other countries. Furthermore, we determined that the induction of serocross reactivity was more efficient within families [16]. Therefore, we selected PspA molecules inducing broad-range cross reactivity within family 1 [28] and family 2 [23].
Pneumolysin (Ply) is a cholesterol-dependent cytolysin that activates complement, induces apoptosis in different host cells, and promotes inflammatory responses and lung injury [29,30,31,32,33,34]. It is the main toxin produced by pneumococci and an important virulence factor. Ply is a TLR4 ligand that not only induces innate immune responses but also induces the TLR4-independent activation of inflammasomes [35,36], contributing to host protection. While Ply is naturally toxic, modified molecules, which are denominated pneumolysoids, display low toxicity while retaining the immunogenic properties of the original protein. PdT (a genetically detoxified form of Ply) is a well-characterized pneumolysoid that has been demonstrated to stimulate cells via Toll-like receptor 4 (TLR4) in a similar fashion to native pneumolysin [35,37].
The immunization of infants with PCVs includes 3–4 doses starting at 2 months, which renders neonates susceptible to infection in the first months of life, where mortality is higher. Partial protection is attained by reducing transmission from vaccinated older children and adults. Therefore, there are efforts to develop vaccines that would induce protection in newborns [38,39]; however, there are very few.
BCG is one of the only vaccines that induces protection in newborns. Our previous results showed that recombinant BCG (rBCG) expressing Bordetella pertussis antigens induced protection against challenge in neonate mice, contrary to the conventional vaccine [40]. We previously constructed a fusion protein comprising a PspA with PdT. This hybrid was able to induce antibodies that can adhere to the surface of pneumococci of different serotypes, promoting protection against lethal pneumococcal challenge and pneumonia [41]. Subsequently, we developed a rBCG strain expressing the rPspA-PdT fusion protein (rBCG PspA-PdT). When administrated in a prime/boost strategy, the rBCG PspA-PdT/rPspA-PdT vaccine induced elevated levels of antibody, with an IgG1/IgG2c antibody isotype shift, and effectively protected mice against a lethal pneumococcal challenge [42]. Furthermore, the immunized mice induced a significant increase in the binding of IgG2c and an improved deposition of complement on the pneumococcal surface in BALF samples, promoting an early clearance of pneumococci [43].
In this context, we investigated the immunogenicity and protective effect of the prime/boost strategy with rBCG PspA-PdT in neonatal mice.
2. Materials and Methods
2.1. Pneumococcal Strain and rBCG Strain
Streptococcus pneumoniae strain WU2 (serotype 3; PspA clade 2; family 1), generously provided by Dr. David Briles (University of Alabama at Birmingham, Birmingham, AL, USA), were cultivated using the methods previously described [41] and stored at −80 °C. Serotype 3 strains are a major cause of severe clinical manifestations of pneumococcal diseases in humans, exhibiting a wide geographical distribution [44,45]. Furthermore, the WU2 strain is also highly lethal in mice, providing a good model of disease [46].
We used an rBCG strain expressing a PspA clade 2, family 1 (previously selected for broad cross reactivity within family 1 and different than the WU2 PspA clade 2), in fusion with PdT, previously constructed in our lab [42]. Briefly, BCG Pasteur was transformed by electroporation with the pMIP12-pspA2-pdT mycobacterial expression vector and vaccines prepared [42].
2.2. Neonatal Mice Immunization
The Ethics Committee at Instituto Butantan, São Paulo, Brazil (CEUAIB) approved all animal experiments (Permit Number 9343010422). Groups of litters comprising 6–10 neonate C57BL/6 were immunized intraperitoneally with 50 μL containing 1 × 10^5^ CFU (1/10th adult mouse dose) of rBCG PspA-PdT or WT-BCG on the 5th day after birth. A booster containing 5 μg of rPspA-PdT (and 50 μg of Al(OH)3 (Alum) in saline, as adjuvant) was administered 7 days following the priming dose (on the 12th day after birth). Control groups received either saline (negative control) or two doses containing 5 μg of rPspA-PdT and 50 μg of Alum (positive control) (on days 5 and 12 after birth) (also i.p.).
2.3. Lethal Pneumococcal Challenge
All mice groups were under anesthetized 9 days after the protein boost (on day 21st after birth) before receiving 10^6^ CFU of the pneumococcal strain WU2 in 30 µL of saline using intranasal aspiration. Survival was observed for a total of 15 days. Mice that were in a moribund state or exhibited any indications of illness were euthanized.
2.4. Antibodies Measurement
Nine days after the protein boost (on day 21st after birth), sera were obtained from mice via retro-orbital collection, and anti-rPspA-PdT antibodies were evaluated using ELISA. The ELISA assay was conducted as previously described [26]. The absorbance was analyzed using an ELISA reader (Multiscan EX—Uniscience, Helsinki, Finland) at OD_450nm_. The standard curve for calculating the antibody concentration in each sample was established using purified mouse IgG, IgG1, or IgG2a (Southern Biotechnology, Birmingham, AL, USA).
2.5. Antibody Binding to Pneumococcal Surface
The evaluation of the binding ability of antibodies from the sera or bronchoalveolar lavage fluid (BALF) of immunized mice to PspA exposed on the pneumococcal surface was assessed as previously described [26]. The WU2 strain was cultured using undiluted sera or BALF samples obtained from immunized mice. Subsequently, the strain was incubated using FITC-conjugated IgG antibody (MP Biomedical) at a dilution of 1:500 in PBS for 30 min. The bacteria were rinsed and placed back into a solution containing 1% paraformaldehyde (PFA). Unstained pneumococci were utilized as a negative control. The samples were then analyzed via flow cytometry using a FACS Canto II (BD, Bioscience, San Jose, CA, USA).
2.6. Spleen and Lungs Cell Culture
Nine days after the protein boost (on day 21 after birth), the animals were euthanized, and the spleens and lungs were collected aseptically. The cell preparations were conducted as previously described [26]. The cell suspensions were adjusted to 1 × 10^6^ cells/mL and incubated with anti-CD28 (1 µg/mL, clone: CD82.2, BD Pharmingen™), anti-CD3 (1 µg/mL, clone: OKT3, BD Pharmingen™), and rPspA-PdT (5 µg/mL) for 48 h at 37 °C and 5% CO_2_. The supernatant sample was collected, and the concentrations of cytokines (IFN-γ, IL-17, TNF-α, IL-10, and IL-6) were evaluated via Cytometric Bead Array (BD Pharmingen™) using the Mouse Th1/Th2/Th17 Cytokine Kit.
Additionally, to investigate the phenotype of memory B and T cells in the lungs and spleen, the remaining cells were labeled with specific antibodies: (i) Memory B cells: anti-B220-FITC antibody (clone: RA3-6B2, BD Pharmingen™), anti-CD19-BV421 antibody (clone: 1D3, BD Horizon™), and anti-CD27-PerCP.Cy5.5 antibody (clone: LG.3A10, BD Pharmingen™); (ii) Memory T cells: anti-CD4-APC.Cy7 antibody (clone: GK1.5, BD Pharmingen™), anti-CD8-PE.Cy7 antibody (clone:53-6.7, BD Pharmingen™), anti-CD62L-FITC antibody (clone: MEL-14, BD Pharmingen™), and anti-CD44-APC antibody (clone: IM7, BD Pharmingen™) for 30 min. Also, the cells were rinsed and resuspended in 1% PFA. Data were acquired using a FACSCanto II flow cytometer.
2.7. Bronchoalveolar Lavage Fluid (BALF) Collection and Cytokine Analysis
For BALF collection, a group of immunized mice was euthanized 3 days after the challenge with the WU2 strain (24 days after birth). The trachea was surgically opened, and a catheter was inserted for cannulation. The lungs were washed twice using cold PBS. The samples were stored on ice and centrifuged at 280× g for 10 min, and the supernatants sample were used to measure antibody binding and cytokine production.
The measurement of cytokines (IFN-γ, IL-17, TNF-α, IL-10, and IL-6) in the BALF supernatant samples was performed using Cytometric Bead Array (BD Pharmingen™) using the Mouse Th1/Th2/Th17 Cytokine Kit, according to the manufacturer’s recommendations.
2.8. Statistical Analysis
The collected samples were individually evaluated and presented as means ± SD. Statistical analyses were conducted using a one-way ANOVA and Bonferroni’s Multiple Comparison Test between the groups. Survival rates in each group were analyzed using the Kaplan–Meier test; p < 0.05 indicated statistical significance.
3. Results
3.1. Prime/Boost Immunization Using rBCG PspA-PdT and rPspA-PdT Promotes IgG1/IgG2c Antibody Isotype Class Shift and Memory B Cells in Neonatal Mouse Model
To evaluate the anti-PspA-PdT antibody production induced by the prime/boost scheme, neonatal mice were vaccinated with rBCG PspA-PdT on day 5 and received a booster with rPspA-PdT (and Al(OH)3 as adjuvant) at day 12). Sera from the immunized mice were evaluated to produce anti-rPspA-PdT antibodies. The groups of mice that received the rBCG PspA-PdT/rPspA-PdT (prime/boost scheme) showed increased total IgG production, comparable to those that received two doses of rPspA-PdT (Figure 1A). The other immunization schemes did not induce significant antibody levels. Mice immunized with rPspA-PdT showed a predominant production of the IgG1 isotype antibodies. Interestingly, mice immunized with rBCG PspA-PdT/rPspA-PdT promoted an IgG class shifting from IgG1 to IgG2c isotype, as seen in adult mice (Figure 1B).
As for the capacity of the antisera to adhere to PspA that is visible on the pneumococcal surface, it was observed that antisera obtained from mice immunized with rBCG PspA-PdT/rPspA-PdT prime/boost exhibited increased IgG binding to pneumococci (strain WU2) (Figure 1C). Surprisingly, the antisera obtained from animals that were administered only rBCG PspA-PdT also exhibited increased binding, although they presented much lower anti-rPspA-PdT antibody levels. However, sera obtained from animals that received two doses of rPspA-PdT (two doses) demonstrated lower binding on the pneumococcal surface, even though it induced high levels of IgG in the ELISA assay (Figure 1A).
Since significant levels of antibodies (IgG and IgG2c) were observed in mice immunized with the prime/boost scheme, the induction of B cells and memory B cells were evaluated in the spleen and lungs of immunized animals, 9 days after the rPspA-PdT booster (gating strategy in Figure S1). All groups of immunized mice displayed an induction of B cells in the spleen (Figure 2A) and lungs (Figure 2B) in comparison to the control group. Vaccination with rBCG PspA-PdT/rPspA-PdT was particularly effective and induced higher levels of B cells. Analysis of the lungs showed significantly higher B cell counts in the prime/boost scheme than in the group receiving two doses of protein. B cells showed a higher percentage in the spleens as compared to the lungs. This is expected since the spleen is a hemopoietic organ with the function of maturation and storage of immune cells.
When analyzing the induction of memory B cells in the spleens and lungs, a similar and more pronounced phenomenon was observed (Figure 2C,D); the rBCG PspA-PdT/PspA-PdT-immunized mice displayed a significant increase in memory B cells compared with all other control groups. Furthermore, the group immunized with only rBCG PspA-PdT also displayed increased memory B cells compared to the controls.
3.2. Prime/Boost Immunization Using rBCG PspA-PdT and rPspA-PdT Induces Inflammatory Cytokine and Memory T Cells in Neonatal Mouse
Cytokine secretion was assessed in the supernatant of spleen and lung cells cultured with rPspA-PdT. Mice receiving rBCG PspA-PdT/rPspA-PdT induced significantly higher levels of IL-6 (Figure 3A), IL-17 (Figure 3D), and IL-10 (Figure 3E) in the lungs compared to the other immunized mice. The levels of TNF-α were also elevated compared to most groups (Figure 3B). Additionally, the levels of IFN-γ were either higher or comparable in the prime (rBCG PspA-PdT) and rBCG PspA-PdT/rPspA-PdT (prime/boost) groups (Figure 3C). When analyzing the data in a radar chart, it becomes clear that the prime/boost rBCG PspA-PdT/rPspA-PdT group shows increased overall cellular immune responses (Figure 3F).
In contrast, the unstimulated cells obtained from mice that were immunized with prime/boost strategy (rBCG PspA-PdT/rPspA-PdT) exhibited elevated levels of IFN-γ, IL-17, TNF-α, IL-10, and IL-6 in the lungs (Figure S2) compared to the stimulated groups. Furthermore, the spleen cells obtained from groups that received rBCG PspA-PdT/rPspA-PdT, when stimulated with rPspA-PdT, showed significantly higher levels of IL-6, TNF-α, IL-17, and IL-10 (Figure S3A,C–E) when compared to the other immunized groups. The animals immunized only with the prime rBCG PspA-PdT also showed higher levels of IFN-γ, TNF-α, IL-10, and IL-6 (Figure S3).
However, vaccination with prime/boost rBCG PspA-PdT/rPspA-PdT showed high levels of inflammatory cytokines; we evaluated the induction of subsets of Memory T cells in the spleen and lungs of immunized mice. Those mice that received the rBCG PspA-PdT or rBCG PspA-PdT/rPspA-PdT had a greater increase in the production of Central Memory T cells (TCM) and Effector Memory T cells (TEM) of both CD4 and CD8 phenotypes (Figure 4A–D).
3.3. rBCG PspA-PdT/rPspA-PdT Protects Neonatal Mice against Lethal Pneumococcal Challenge
The efficacy of vaccination with rBCG PspA-PdT/rPspA-PdT in neonate mice was assessed using a lethal pulmonary challenge. Seventy-two hours after the challenge, the BALF was recovered and analyzed from cytokine production. All groups of immunized mice had decreased levels of IL-6, IFN-γ, and IL-17 in the lung cells after infection (Figure 5A,C,D). Interestingly, only mice that received rBCG PspA-PdT (prime or prime/boost) displayed elevated levels of TNF-α and IL-10 (Figure 5B,E).
At this time point, BALF samples were collected to evaluate the capacity of the bind to the pneumococcal surface. BALF samples obtained from animals that received rBCG PspA-PdT/rPspA-PdT displayed significantly greater levels of total IgG binding to pneumococci (~29% of positive cells). This was followed by rBCG PspA-PdT (~19% of positive cells) (Figure 6A,B). Mouse survival was monitored for 15 days, showing that only rBCG PspA-PdT/rPspA-PdT provides 100% protection against fatal challenges (Figure 6C). Interestingly, although the prime/boost with protein alone showed comparable levels of anti-PspA-PdT antibodies as those primed with rBCG, the protection observed was much lower. Furthermore, mice that received rBCG PspA-PdT (prime dose) or WT-BCG/rPspA-PdT presented partial protection in this model of neonatal mice.
4. Discussion
Despite the successful implementation of conjugate pneumococcal vaccines in many countries, there remain important limitations, including serotype replacement and geographic variations in serotype distribution, which compromise the long-term success of such formulations. Proteins are considered the most effective means of attaining broad cross-reacting immunity. However, proteins have not shown sufficient immunogenicity in humans, even using different adjuvant formulations. Therefore, new presentation systems should be investigated.
BCG has been used in newborn vaccination against tuberculosis for over a hundred years, displaying high safety; it is one of the few vaccines that can induce protective immunity in neonate children. BCG has shown the induction of long-lasting immunity in humans, and the recombinant BCG strategy could show interesting results. Its potent adjuvanticity reinforces its use as a platform to deliver heterologous proteins from different pathogens. Previously, we showed in a prime/boost strategy that the rBCG PspA-PdT/rPspA-PdT immunization scheme induced high levels of antibody, with an IgG1/IgG2c antibody isotype shift [42] and high protection in adult mice. The present study investigated the use of this prime/boost strategy as a vaccine targeting newborns. The prime/boost strategy in the neonate mouse model induced similar antibody levels in comparison with the recombinant protein alone (Figure 1A). The presence of rBCG in the priming dose promoted an antibody shift to IgG2c (Figure 1B). This isotype change has been previously observed in an adult model [42]. In both adults and neonate mice, immunization with rPspA (in Alum) primarily showed an IgG1 isotype profile, which is protective in adults but not neonates [42].
Furthermore, in the adult model, IgG2a/c and IgG2b antibodies present in the serum and the BALF from mice immunized with the prime/boost strategy also showed increased binding to the pneumococcal surface, indicating that the vaccine antibodies can recognize the native proteins expressed by the bacterium [42,43,47,48]. In the neonatal model, we observed comparable results, showing that the prime/boost strategy induced high levels of IgG2c antibodies, and the serum and BALF were able to bind to the pneumococcal surface, promoting an early clearance of pneumococci (Figure 1C,D; Figure 6A,B). Unfortunately, we did not have a sufficient amount of BALF sample to measure IgG isotypes.
Most interesting is that although the immunization with rBCG-PspA-PdT only induced lower levels of antibodies to the fusion protein, these antibodies showed a more balanced proportion of IgG1/IgG2c and high binding activity to the pneumococci surface. These results may partly explain the higher protection induced by rBCG-PspA-PdT.
Regarding cytokine production, vaccination with the prime/boost scheme in the neonatal model induced an increase in the secretion of IL-6, IL-17, TNF-α, and IFN-γ in lung cells (Figure 3). Interestingly, these cytokines have been described as protective against pneumonia. IL-17 is considered important in the recruitment of neutrophils to the site of the infection, correlating with the highest protection level, mainly in a mouse model of pneumococcal colonization [49,50,51]. The secretion of IFN-γ and TNF-α via lung cells has been correlated with protection against pneumonia [52,53,54]. In oral administration of non-recombinant Lactobacillus casei [55], the balance of the induction of cytokines TNF-α and IL-10 was related to the protection against lung injuries caused by S. pneumoniae infection due to the rapid increase in the infiltration of neutrophils.
Another interesting result is the higher protection observed via immunization with WT-BCG/rPspA-PdT. This has been previously observed in adult mice [43]. This effect has been attributed to the recently described innate immune memory (or trained immunity) properties of BCG, where increased protection against heterologous pathogens has been characterized. However, this protection is not expected to be long-lasting. In fact, this group should lower the production of memory B cells (Figure 2).
Besides the generation of specific antibodies in serum and cytokines in the lung, priming with rBCG PspA-PdT also resulted in enhanced production of memory B- and T-cells, including TCM and TEM cell populations, in the spleen and lungs (Figure 4). Previous studies have shown that regardless of parenteral or intranasal administration, the memory B and T cell populations residing in the lungs can mediate protection by enhancing the production of pneumococcus-reactive antibodies in the lungs and generating neutrophil-dependent protection against pneumococcal nasal challenge [56,57,58,59]
A significant reduction in cytokine production was observed in the lungs of prime/boost-immunized mice following intranasal challenge (Figure 5). This correlated with the 100% survival after the challenge (Figure 6), suggesting that a more controlled inflammation is important for protection against pneumococcal sepsis. Similar results have been described in previous studies investigating PspA-based vaccines [54]. The subcutaneous immunization of adult mice led to a reduction in IL-6 and IFN-γ and the induction of TNF-α post-challenge in the lungs [54]. Our group has previously shown that the prime/boost strategy provided 100% protection in adult mice after pneumococcal lethal challenge [42].
One of the limitations of this study is the reduced number of studies with neonates in the literature, especially in the BALF, which provided few options for proper sampling design. We adopted alternative designs, more appropriate for our objectives, which, unfortunately, reduces the possibility of comparison with previous data. Another important factor was the fact that obtaining a reasonable amount of neonate mice at the same time requires very careful coordination for their birth to be able to obtain several litters at the same time. Furthermore, the amount of blood and BALF obtained is much more limited than in adult mice, restricting the number of samples available for analysis. Another important limitation is the coverage provided by the PspA component of the antigen. Our studies are based on a PspA fragment from family 1, previously demonstrated to induce broad protection within family 1 strains [28]. Although PdT is highly conserved in all pneumococcal strains, it is not expected to provide high protection, working mostly as an adjuvant. In order to attain wide coverage, our strategy would be to include another rBCG strain expressing a family 2 PspA. We previously selected a family 2 PspA inducing broad cross-reactive protection [23] and a fusion protein of this family 2 PspA with PotD, induced protection against invasive challenge and colonization with strains containing PspA family 1 and 2 [60]. Our plan is to investigate the prime/boost strategy with a mixture of rBCG-PspA1-PdT and rBCG-PspA2-PotD, which has the potential to induce protection against invasive challenge and colonization in neonate mice.
5. Conclusions
The results of the present study demonstrate that rBCG PspA-PdT is an effective platform for the delivery of pneumococcal antigens, and the prime/boost immunization promotes strong antibody and cell-mediated responses, generating memory T and B cells and conferring protection against invasive infection in neonate mice.
BCG vaccine has been employed for more than a century with the goal of protecting newborns against tuberculosis, demonstrating the induction of long-lasting immunity and a commendable safety profile. Recombinant BCG vaccines are expected to carry the main properties of BCG and therefore have the potential to induce long-term protection. Our results reinforce the potential of the rBCG-expressing pneumococcal proteins in a prime/boost strategy to be further investigated in humans.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1WHO Pneumonia in Children Available online: https://www.who.int/news-room/fact-sheets/detail/pneumonia(accessed on 1 September 2023)
- 2O’Brien K.L. Wolfson L.J. Watt J.P. Henkle E. Deloria-Knoll M. Mc Call N. Lee E. Mulholland K. Levine O.S. Cherian T. Burden of Disease Caused by Streptococcus Pneumoniae in Children Younger than 5 Years: Global Estimates Lancet 200937489390210.1016/S 0140-6736(09)61204-619748398 · doi ↗ · pubmed ↗
- 3Oligbu G. Hsia Y. Folgori L. Collins S. Ladhani S. Pneumococcal Conjugate Vaccine Failure in Children: A Systematic Review of the Literature Vaccine 2016346126613210.1016/j.vaccine.2016.10.05027838066 · doi ↗ · pubmed ↗
- 4Feikin D.R. Kagucia E.W. Loo J.D. Link-Gelles R. Puhan M.A. Cherian T. Levine O.S. Whitney C.G. O’Brien K.L. Moore M.R. Serotype-Specific Changes in Invasive Pneumococcal Disease after Pneumococcal Conjugate Vaccine Introduction: A Pooled Analysis of Multiple Surveillance Sites P Lo S Med.201310 e 100151710.1371/journal.pmed.100151724086113 PMC 3782411 · doi ↗ · pubmed ↗
- 5Lehmann D. Willis J. Moore H.C. Giele C. Murphy D. Keil A.D. Harrison C. Bayley K. Watson M. Richmond P. The Changing Epidemiology of Invasive Pneumococcal Disease in Aboriginal and Non-Aboriginal Western Australians from 1997 through 2007 and Emergence of Nonvaccine Serotypes Clin. Infect. Dis.2010501477148610.1086/65244020420501 · doi ↗ · pubmed ↗
- 6Richter S.S. Diekema D.J. Heilmann K.P. Dohrn C.L. Riahi F. Doern G.V. Changes in Pneumococcal Serotypes and Antimicrobial Resistance after Introduction of the 13-Valent Conjugate Vaccine in the United States Antimicrob. Agents Chemother.2014586484648910.1128/AAC.03344-1425136018 PMC 4249410 · doi ↗ · pubmed ↗
- 7Keller L.E. Robinson D.A. Mc Daniel L.S. Nonencapsulated Streptococcus Pneumoniae: Emergence and Pathogenesis M Bio 20167 e 01792-1510.1128/m Bio.01792-1527006456 PMC 4807366 · doi ↗ · pubmed ↗
- 8Hsu H.E. Shutt K.A. Moore M.R. Beall B.W. Bennett N.M. Craig A.S. Farley M.M. Jorgensen J.H. Lexau C.A. Petit S. Effect of Pneumococcal Conjugate Vaccine on Pneumococcal Meningitis N. Engl. J. Med.200936024425610.1056/NEJ Moa 080083619144940 PMC 4663990 · doi ↗ · pubmed ↗