Identification of Flo11-like Adhesin in Schizosaccharomyces pombe and the Mechanism of Small-Molecule Compounds Mediating Biofilm Formation in Yeasts
Yu-Gang Zhang, Tong Zhang, Lan Lin

TL;DR
This paper identifies a protein in a type of yeast that helps it form biofilms and shows how certain small molecules can either promote or inhibit this process.
Contribution
The study confirms a protein in Schizosaccharomyces pombe as a functional homolog of Flo11 and reveals how small molecules regulate biofilm formation.
Findings
Deletion of SPBPJ4664.02 in S. pombe reduces biofilm formation and invasive growth.
Indole-3-acetic acid and dodecanol inhibit biofilm formation at high concentrations but promote it at low concentrations.
IAA and dodecanol likely act downstream of SPBPJ4664.02 in the biofilm signaling pathway.
Abstract
Fungal infection is initiated by the adhesion of pathogens to biotic and abiotic surfaces, with various manifestations including biofilm formation and invasive growth, etc. A previous report, though devoid of functional data, speculated that the Schizosaccharomyces pombe glycoprotein SPBPJ4664.02 could be the homology of Saccharomyces cerevisiae Flo11. Here, our studies with S. pombe substantiated the previously proposed speculation by (1) the deletion of SPBPJ4664.02 attenuated biofilm formation and invasive growth in S. pombe; (2) the S. pombe’s lack of SPBPJ4664.02 could be complemented by expressing S. cerevisiae flo11. Furthermore, indole-3-acetic acid (IAA) and dodecanol were examined in S. pombe for their respective effects on biofilm formation. IAA and dodecanol at high concentrations could inhibit biofilm formation, whereas opposing effects were observed with low concentrations…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
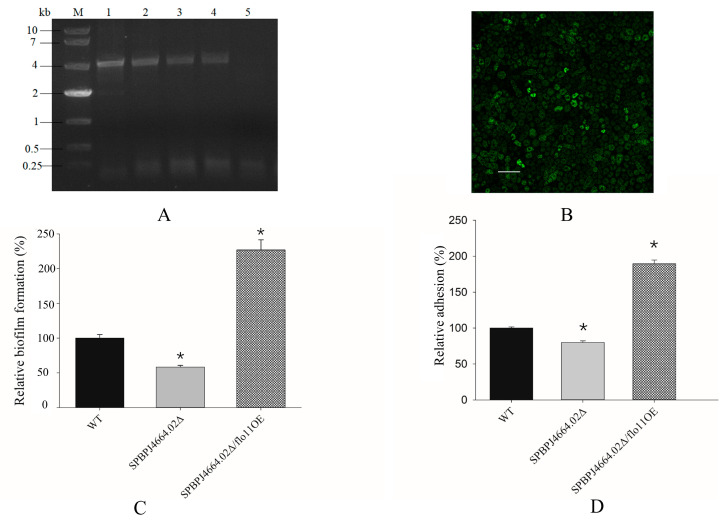 Figure 1
Figure 1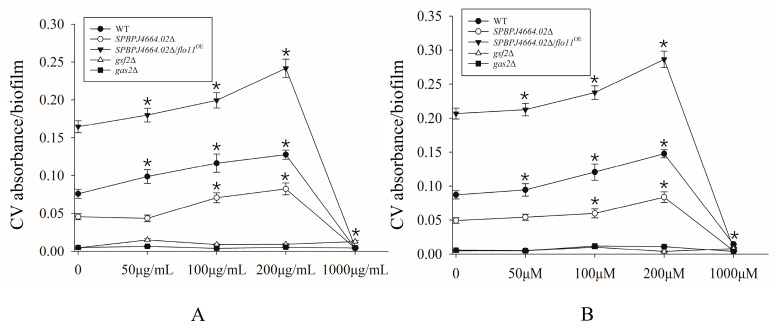 Figure 2
Figure 2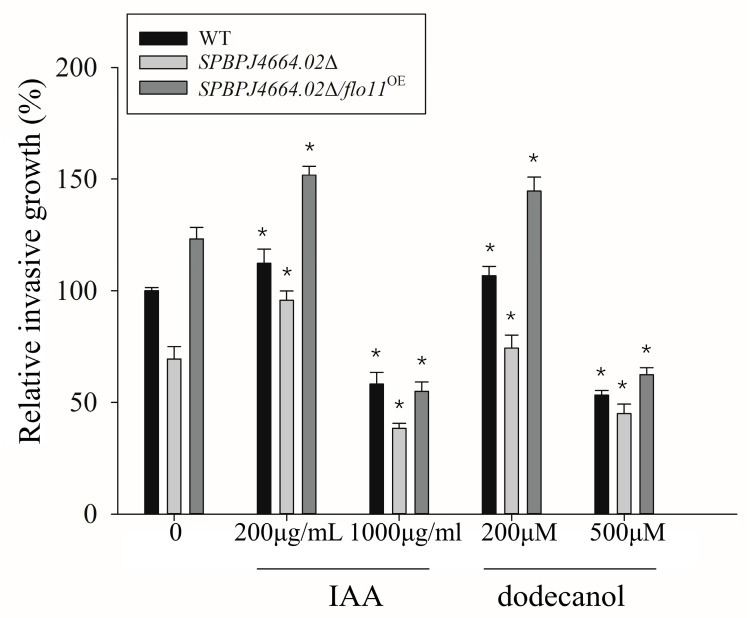 Figure 3
Figure 3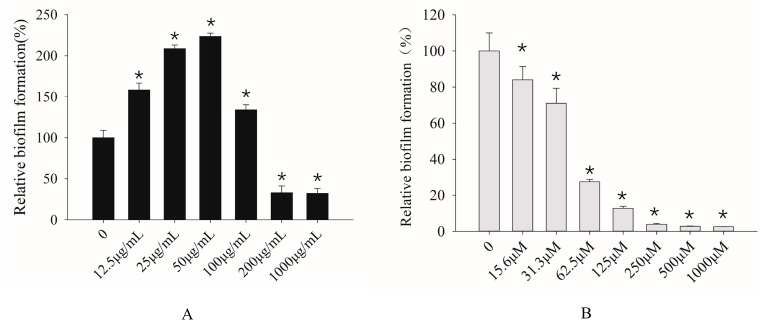 Figure 4
Figure 4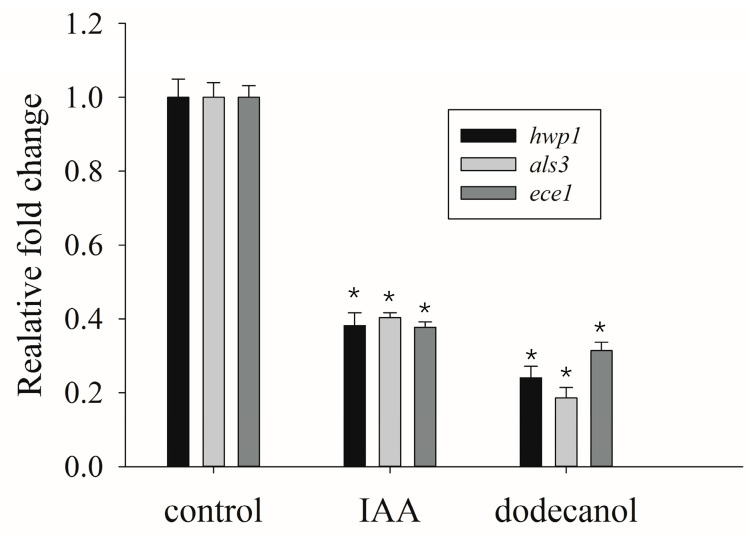 Figure 5
Figure 5- —National Natural Science Foundation of China
- —Natural Science Foundation of Jiangsu Province, China
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntifungal resistance and susceptibility · Fungal and yeast genetics research · Microbial Natural Products and Biosynthesis
1. Introduction
Human health is threatened by fungal infection. Beyond superficial fungal infections, invasive infections by fungi such as Candida spp. are known to impact people with compromised immunity due to medical interventions—for example, those undergoing chemotherapy therapy, those who underwent organ transplantation, and those with immunosuppressive diseases including AIDS [1]. More recently, a link has been reported between Candida albicans intestinal outgrowth and lasting immune activation during severe COVID-19, which may exacerbate the disease outcomes [2].
Yeast cells including non-pathogenic Saccharomyces cerevisiae, Schizosaccharomyces pombe, and pathogenic Candida albicans, in response to adverse environmental factors, may undergo a morphological transition from yeast to hyphal forms, the so-called filamentous growth, and, if necessary, develop into sophisticated biofilm to achieve adaptation fitness [3,4]. The morphological transition of yeasts is tightly associated with biofilm formation. The biofilm-forming ability of pathogenic yeast is recognized to be the virulence attribute as well as a major causal factor of antifungal resistance and refractory infection, thereby posing a great challenge in clinical scenarios [5,6]. Accordingly, there is in urgent need to explore and develop novel anti-biofilm strategies to fight against the increasingly emerging resistant fungal pathogens such as Candida albicans.
Fungal adhesion to biotic and abiotic surfaces is a crucial prerequisite for pathogen colonization and the ensuing biofilm formation [7]. The biomaterial of medical implants is appropriate for fungal cell attachment and adherence, and the utilization of indwelling devices such as catheters and prosthesis may pose a high risk for fungal biofilm infections [6,8]. The fungal adhesion process depends upon a class of cell surface glycoproteins, namely, fungal adhesins or flocculins [1]. Flo adhesins have been first discovered in brewer’s yeast Saccharomyces cerevisiae, among which Flo11 is the main adhesin for controlling filamentous growth, mat, and biofilm formation [7,9].
Schizosaccharomyces pombe is an eminent model organism for investigating yeast adhesion and biofilm formation. It has been speculated by previous studies that SPBPJ4664.02 might be S. pombe Flo11-like adhesin [10], based on the following facts: (1) SPBPJ4664.02 shares 55.1% of its homology with Flo11 in the functional domain through a database search; and (2) the transcripts of SPBPJ4664.02, via microarray analysis, were found to be upregulated in two ribosomal protein-deficient mutants of S. pombe, namely, rpl32-1Δ and rpl32-2Δ, in which enhanced cell flocculation was observed [11]. However, it has not been substantiated whether SPBPJ4664.02 is the functional homology of Flo11 adhesin in S. pombe. Thus, in this study, a gene deletion mutant of SPBPJ4664.02 was constructed, followed by the rescue experimentation of the resulting SPBPJ4664.02Δ with the heterologous flo11 gene. One of our goals is to verify that the functional role of SPBPJ4664.02 is Flo11-like.
The morphological switch from yeast to hyphal forms as well as biofilm formation are governed by many environmental cues acting through several signaling molecules in a number of yeasts, including C. albicans [12,13,14]. Although indole-3-acetic acid (IAA) is a well-known phytohormone in planta, mounting evidence has revealed the presence of IAA and its effects on the morphological changes in some yeasts [13,15,16]. In S. cerevisiae, IAA could induce adhesion and hyphal growth at lower concentrations, while at higher concentrations, it could inhibit the growth [13]. Apart from the small-molecule compound IAA, dodecanol, a C_12_ fatty alcohol, was demonstrated to modulate hyphal development in C. albicans [17,18]. Notwithstanding, the mechanisms regarding such chemical signaling molecules as IAA and dodecanol influence the yeast morphologic transition, and the biofilm formation in S. pombe is poorly understood. Also, how the adhesins such as Flo11-like protein (SPBPJ4664.02) and Gsf2 (galactose-specific flocculin 2, encoded by gsf2) interplay with chemical signaling molecules, thereby giving rise to the biofilm formation, merits in-depth investigation.
In this study, the mechanism was elucidated regarding the effects of the small-molecule compounds IAA and dodecanol on invasive (hyphal) growth and biofilm formation using S. pombe. The impacts of the above-described compounds on the biofilm formation of C. albicans were further examined. This paper represents the first report dealing with the mechanistic studies in a well-defined model of yeast S. pombe and subsequent extrapolating to the pathogenic yeast C. albicans, which is of clinical significance. Our research provides novel insight that either IAA or dodecanol have potential as the synergistic anti-film agent in combination with available antifungals to eradicate the biofilm-forming pathogenic yeast C. albicans.
2. Materials and Methods
2.1. Strains and Media
Saccharomyces cerevisiae cells were routinely grown on YPD medium (Sangon Co., Ltd., Shanghai, China). Schizosaccharomyces pombe cells were routinely grown on YE5S medium—5 g/L yeast extract (Oxoid, UK), 30 g/L glucose supplemented with 0.225 g/L of adenine, uracil, histidine, leucine, and lysine (Sigma, St. Louis, MO, USA). For yeast adhesion and biofilm assays, S. pombe wild-type, the gene-deletion mutant, and gene overexpressed strains were cultivated on LNB medium—0.067 g/L yeast nitrogen base without amino acids (Solarbio, Beijing, China), 20 g/L glucose, and salts and vitamins, as for EMM [3] and 2% w/v agar. The above-described yeast strains originated from our laboratory collection or were created in this study. The PCR-tagging vector pFA6a-KanMX6 used for generating the specific gene deletion mutant of S. pombe [19] and the GFP-tagging shuttle vector pSGP572a are gifts kindly provided by Dr. P. T. Tran, University of Pennsylvania, USA.
Candida albicans SC5314, a known clinical isolate from CGMCC (China general microbiological culture collection center, Beijing, China), was routinely cultured on YPD agars and, if needed, grown in RPMI1640 broth (Acmec Biochemical, Shanghai, China).
2.2. Construction of the SPBPJ4664.02Δ Deletion Mutant
The SPBPJ4664.02-targeting DNA fragments containing KanMX6, the so-called transformation module (Figure S1), were PCR-amplified from the genomic DNA of WT cells and plasmid pFA6a-KanMX6 using deletion primer sets (Table S1). The transformation module was introduced into a WT fission yeast cell targeting the chromosomal locus of SPBPJ4664.02 to generate the SPBPJ4664.02Δ mutant via the homologous recombination technique [20]. For details, refer to the Supplementary Methods.
2.3. Construction of the SPBPJ4664.02Δ/flo11OE
The flo11 gene from S. cerevisiae BY4742 genomic DNA (gDNA) was amplified via PCR using Hi-fidelity DNA polymerase (ToYoBo, Tokyo, Japan), followed by sub-cloning into the pSGP572a vector to ensure in-frame expression with GFP under the promoter nmt1 using the primers F_BglII-flo11 and R_Not I-flo11 (see Table 1 for nucleotide sequences) and the ClonExpress^®^ Ultra One Step Cloning Kit (Vazyme, Nanjing, China), generating the recombinant vector pSGP572a-flo11 (see Figure S4 in the SI for a schematic representation). Next, the SPBPJ4664.02Δ cells of S. pombe were transformed with pSGP572a-flo11 using lithium acetate transformation, as previously described in Siam et al. [21], yielding the SPBPJ4664.02Δ/flo11^OE^ strain of S. pombe. Ura4^+^ transformants (positive clones) were screened on solid EMM plates, and the orientation of flo11 inserted in the transformants was verified by colony PCR with the primers listed in Table 1. Since Flo11 is expressed in fusion with GFP, the cells of SPBPJ4664.02Δ/flo11^OE^ could be observed under confocal laser scanning microscopy (CLSM).
2.4. Molecular Characterization of SPBPJ4664.02Δ/flo11OE for Flo11-GFP Expression
2.4.1. Colony PCR of SPBPJ4664.02Δ/flo11OE
Transformants were grown on a solid EMM plate at 30 °C for 3 days. Four individual colonies were then selected, followed by overnight culture in YE5S for yeast proliferation. Yeast genomic DNA was obtained using the TIANamp Yeast DNA Kit (TIANGEN, Beijing, China). Colony PCR was conducted by using the primers F_nmt and R_GFP, as listed in Table 1, and the respective transformant genomic DNA as the template, where SPBPJ4664.02Δ was used as the negative control.
2.4.2. Confocal Laser Scanning Microscopy (CLSM)
Yeast cell colonies harboring flo11-gfp fusion expression (SPBPJ4664.02Δ/flo11^OE^) were selected for overnight culture on the EMM medium. Suspensions of cells at an exponential phase were examined on a glass slide under CLSM, where the GFP was excited at 488 nm. Images were collected using an Olympus FV3000 confocal microscope with a Plan-Apo 100X/1.4 Oil objective lens, analyzed by FV31S-SW Olympus.
2.5. Assays of Biofilm Formation in S. pombe
The biofilm formation of fission yeast on the abiotic surface was determined using a microtiter plate-based assay, as described by Kimani et al. [22]. S. pombe strains of WT, gene-deletion mutants (gsf2Δ, gas2Δ, and SPBPJ4664.02Δ), and gene-overexpressed strains (SPBPJ4664.02Δ/flo11^OE^) were examined. Briefly, overnight cultures of S. pombe strains (OD_600_ 0.5, c.a. 10^7^ cells/mL) were harvested. One hundred microliter aliquots of the cell suspensions were transferred into 96-well flat-bottomed polystyrene plates (Corning, USA). After 8 h of cell adhesion at 30 °C, the planktonic cells were removed from each well and the plates were rinsed twice with physiological saline. After drying for 10 min, 100 µL of the LNB containing indole acetic acid (IAA) or dodecanol at different concentrations was added to each treated well. IAA and dodecanol were dissolved in 50% ethanol, giving rise to the stock solutions of 100 mg/mL and 100 μM, respectively. The control well contained the solvent (50% ethanol) of an equal volume to that used in the respective treated well as well as the growth broth lacking small-molecule compounds to make the volume 100 µL. The microplates were incubated for 72 h at 30 °C, followed by the crystal violet (CV) staining protocol to examine the biofilm cells, as described in Kimani et al. [22], where absorbance at 590 nm (A_590_) was recorded. The assays were carried out in six replicates for each treatment. The inhibition extent of biofilm formation was calculated by the following formula:
2.6. The Plate-Washing Assay
This method was carried out as previously described in Cullen (2015) [23], with slight modifications. Briefly, strains of S. pombe of different genotypes were cultured in LNB liquid to an OD_600_ of 0.8–1.0. The cell suspension of 15 μL was patched on LNB agar with an underlying layer of YE5S agar. The plates were incubated at 30 °C for 14–21 d and photographed. Running tap water was used to wash noninvasive cells from the surface, and then the plate was photographed again (for adhesion examination). Finally, under running tap water, a gloved finger was used to rub the plate and more thoroughly remove noninvasive cells, and the plate was photographed a third time (for the filamentation test). The photographs of colonies were quantitated by densitometry using ImageJ.
2.7. Effects of IAA and Dodecanol on C. albicans Growth
To examine the effects of small-molecule compounds on the growth of planktonic C. albicans, a suspension of 2 mL at the appropriate cell density in YPD was cultivated, supplemented with IAA and dodecanol, respectively, followed by colorimetric measurement at 600 nm. Please refer to the Supplementary Methods for details.
2.8. Assays for the Biofilm Formation of C. albicans
The biofilm formation of C. albicans on polystyrene was detected by a microtiter plate-based protocol, as previously described for fission yeast S. pombe (refer to Section 2.5), with a slight modification. Overnight cultures of C. albicans strains in YPD medium were harvested. The cells were washed twice in sterile physiological saline and diluted in fresh RPMI1640 medium to a concentration of 10^6^ CFU/mL. The aliquots of cell suspensions of 100 μL were transferred into flat-bottomed 96-well polystyrene microplates (Corning, Corning, NY, USA). After incubation for 2 h at 37 ℃, the planktonic cells were removed from each well and the plates were rinsed twice with physiological saline. After drying for 10 min, 100 µL of the RPMI1640 medium containing IAA or dodecanol at different concentrations was added to each treated well. The control well contained the solvent of an equal volume to that used in the respective treated well as well as growth broth lacking small-molecule compounds to make the volume 100 µL. The plates were then incubated for 48 h at 37 ℃. For biofilm formation detection, the previously described crystal violet staining was used, in which A_590_ values served as the endpoints of biofilm formed on 96-well microplates [22,24] The assays were carried out in six replicates for each treatment.
2.9. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
Overnight cultures of fresh C. albicans SC5314 in YPD were collected, centrifuged at 1000× g for 3 min, and washed with sterile physiological saline and then resuspended in RPMI 1640 medium to a final concentration of 10^6^ CFU/mL. The cell suspensions of 2 mL were inoculated to each polystyrene flat-bottomed six-well plate and incubated at 37 °C for 2 h. The suspensions were pipetted out, followed by gentle washing of the wells with sterile physiological saline twice to remove non-adhesive cells. Subsequently, the respective well was supplemented with 1000 μg/mL IAA or 200 μM dodecanol in 2 mL of RPMI 1640, and RPMI 1640 medium was only used as the negative control, followed by incubation at 37 °C for 48 h. After the incubation, the upper medium of each well was discarded, the wells were washed twice with physiological saline. The biofilm of each well was scraped off, suspended in 1 mL of physiological saline, and transferred to a 1.5 mL Eppendorf (Ep) tube. The samples were centrifuged and rinsed with sterile water twice. The RNA was extracted from the biofilm samples in each Ep tube using the Spin Column Yeast Total RNA Purification Kit (Sangon Biotech, Shanghai, China), and cDNA synthesis was conducted using PrimeScript™ RT Master Mix (TaKaRa, Kusatsu City, Japan).
qPCR analyses of the C. albicans biofilm-related genes hwp1, ece1, and als3 were performed using TB Green^®^ Premix Ex Taq™ II (TaKaRa, Japan), according to the manufacturer’s manual [25], with the primers listed in Table S2, where the β-actin gene act1 was used as an internal control to standardize the transcripts of the genes. All analyses were conducted in the Applied Biosystems StepOnePlus Real-Time PCR System in triplicate for each sample.
2.10. Statistical Analysis
In this study, the significant differences between treated and untreated samples were determined by Student’s t-test, where p < 0.05 was considered as statistical significance and indicated by asterisks.
3. Results and Discussion
SPBPJ4664.02 is a member of fission yeast S. pombe glycosylphosphatidylinositol-modified (GPI) glycoproteins in association with the adhesion of fission yeast cells [26]. To test the hypothesis that SPBPJ4664.02 may be the fission yeast homology of Flo11, a known adhesin of the brewer’s yeast S. cerevisiae [7], our study examined the biofilm-forming abilities and filamentation assays following the genetic construction of the SPBPJ4664.02Δ mutant strain (see Supplemental Information) and the subsequent complementation experiment by introducing the heterologous (S. cerevisiae) flo11 gene into the SPBPJ4664.02Δ of fission yeast, in comparison to wild-type (WT) fission yeast.
3.1. Rescue of the SPBPJ4664.02Δ Mutant by flo11 Overexpression
By introducing the flo11 gene of S. cerevisiae origin into SPBPJ4664.02Δ fission yeast cells with the aid of pSGP572a-flo11, the transformants of SPBPJ4664.02Δ/flo11^OE^ were obtained on EMM plates, followed by colony PCR identification (Figure 1). Four transformants were selected, and SPBPJ4664.02Δ was used as the negative control. Colony PCR showed the successful transformation of SPBPJ4664.02Δ cells with the recombinant vector pSGP572a-flo11 (Figure 1A).
Since flo11 was co-expressed with gfp under the Pnmt1, the expressed GFP was examined under CLSM. Notably, the Flo11-GFP fusion protein was expressed in the interior and periphery of SPBPJ4664.02Δ cells under CLSM (Figure 1B).
In the biofilm assay (Figure 1C), the biofilm-forming ability of different genotypes of S. pombe was determined. The extent of biofilm formed in SPBPJ4664.02Δ decreased by 40% as compared to WT (p < 0.05), indicating that SPBPJ4664.02 is required for biofilm formation. In addition, as shown in Figure 1C, SPBPJ4664.02Δ/flo11^OE^ tremendously restored the biofilm formation ability manifested by SPBPJ4664.02Δ in comparison to WT (p < 0.05), which clearly demonstrated that flo11 is involved in biofilm formation as well as that the expression of flo11 might compensate for the deficiency of SPBPJ4664.02.
The agar adhesion abilities of different fission yeast genotypes were also tested. As shown in Figure 1D, SPBPJ4664.02Δ was found to form less adhesion than WT (p < 0.05), whereas SPBPJ4664.02Δ/flo11^OE^ restored the adhesion (p < 0.05) that was compromised in SPBPJ4664.02Δ. The results not only indicated the involvement of SPBPJ4664.02 and flo11 in the adhesion but also implied that flo11 overexpression could make up for the impairment of SPBPJ4664.02.
Altogether, the gene deficiency of SPBPJ4664.02 in the SPBPJ4664.02Δ mutant of fission yeast appeared to be complemented by expressing the S. cerevisiae-originated flo11, suggesting the functional similarity of SPBPJ4664.02 and flo11, which is reminiscent of the previous studies of Watson and Davey [27] and Younes and Khalaf [28]. Taking into account of the consensus sequence of SPBPJ4664.02 sharing with Flo11 via BLAST searches and the upregulated transcripts of SPBPJ4664.02 in rpl32-1Δ and rpl32-2Δ, the two ribosomal protein-deficient mutants of S. pombe concomitant with enhanced cell flocculation [11], it is strongly indicated that SPBPJ4664.02 is the fission yeast homology of flo11.
3.2. Impacts of SPBPJ4664.02 (Flo11-like) in the Small-Molecule Signaling Pathway for Biofilm Formation and Filamentation
3.2.1. Effects of IAA and Dodecanol on S. pombe Biofilm Formation and Cell Adhesion
Previous studies have shown that the small-molecule compound IAA at low concentrations could induce the adhesion and filamentation of the budding yeast S. cerevisiae, whereas at high concentrations, the inhibitory effects of IAA were observed [13]. Dodecanol is a structural analogue of farnesol, a known quorum sense (QS) signaling molecule that inhibits S. cerevisiae filamentation (viz. morphologic transition) and likewise in the case of C. albicans [29,30]. Since morphologic transition is a prerequisite for the yeast biofilm formation [4], it is hypothesized that the above-mentioned compounds, IAA and dodecanol, could be involved in the formation of yeast biofilm, likely via the dose-dependent mode. In this study, the effects of different levels of IAA and dodecanol on the biofilm formation and cell adhesion in fission yeast were investigated.
It is widely recognized that the genes gsf2 (coding the galactose-specific flocculin) and gas2 (coding the cell wall remodeling enzyme) are both involved in the flocculation of S. pombe [31,32]. When IAA or dodecanol was added, the results indicated that the effect of small-molecule compounds on biofilm formation was dose-dependent within a certain concentration range (Figure 2). The formation of biofilms was observed to elevate in SPBPJ4664.02Δ, SPBPJ4664.02Δ/flo11^OE^, as well as WT fission yeasts upon the supplementation of IAA at concentrations less than 200 μg/mL. Similarly, promotive effects of dodecanol (<200 μM) on the biofilm formation were found in the SPBPJ4664.02Δ, SPBPJ4664.02Δ/flo11^OE^, as well as WT fission yeasts. In contrast, at high concentrations (for example, 1 mg/L IAA or 1 mM dodecanol), c.a. 90% of the tested yeasts displayed suppressive biofilm formation (p < 0.05). Neither gsf2Δ nor gas2Δ exhibited biofilm formation at the tested concentrations of IAA and dodecanol. The fact that gsf2Δ or gas2Δ biofilm formation was unaffected by either of IAA and dodecanol indicated that IAA or dodecanol molecules might act independently of Gsf2 (another flocculin) and Gas2 (the cell wall remodeling protein) in the signaling pathways for biofilm formation. Even in the untreated controls (no IAA or dodecanol addition), biofilm formation was barely found, suggesting that gsf2 and gas2 are indispensable to the biofilm formation process. Taking into account that fission yeast gsf2 encodes a kind of flocculins, the knock-out of gsf2 would abolish the cell flocculation, thereby blocking biofilm formation. Given that gas2 encodes 1,3-β-glucanosyltransferase, the deletion of gas2 would annihilate the cell wall architecture and thus cell flocculation and adhesion, thereby retarding biofilm formation.
3.2.2. Effects of IAA and Dodecanol on S. pombe Invasive Growth
As hyphal growth could have implications for certain important pathogenic yeasts, studying morphological transitions would provide considerable insights into pathogenicity and drug resistance. On this basis, the effects of different concentrations of IAA and dodecanol on the invasive growth (filamentation) of WT, SPBPJ4664.02Δ, and SPBPJ4664.02Δ/flo11^OE^ S. pombe were investigated. As shown in Figure 3, IAA and dodecanol at low concentrations, at 200 μg/mL and 200 μM, respectively, could promote the invasive growth to some extent, while at high concentrations, the invasive growth of S. pombe was remarkably inhibited. No relative invasive growth was obtained for gsf2Δ and gas2Δ because none exhibited filamentation, as previously explained in Section 3.2.1.
The biofilm formation and filamentation of SPBPJ4664.02Δ were influenced by IAA or dodecanol and the two traits changed with the concentration of small molecules, but they never reached the manifestation degrees of WT in terms of biofilm formation and filamentation. Consequently, we postulate that the small molecules and SPBPJ4664.02 operate in the same signaling pathway, in which small molecules likely act in the downstream of SPBPJ4664.02. Our mechanism studies with S. pombe herein revealed that small molecules, namely, IAA and dodecanol, might exert actions at the site downstream of Flo11-like adhesin SPBPJ4664.02 in the signaling pathway towards filamentation and biofilm formation, which represents one of the virulence traits as well as resistance in pathogenic fungi such as Candida albicans.
3.3. Effects of IAA and Dodecanol on C. albicans Biofilm Formation
Given the influence of IAA and dodecanol on the biofilm formation of fission yeast, the effects of small molecules were further investigated on the biofilm formation in C. albicans SC5314, which is of clinical significance [33]. For IAA, it promotes biofilm formation in C. albicans at low concentrations while inhibiting biofilm formation at high concentrations (such as 200 μg/mL and 1000 μg/mL) by over 60% as compared to untreated counterparts (p < 0.05) (Figure 4A). Unlike IAA, dodecanol suppressed the biofilm formation of C. albicans by more than 70% in comparison to untreated counterparts (p < 0.05), with the minimal inhibition concentration as 62.5 μM (Figure 4B). The inhibitory impacts of dodecanol on biofilm formation showed a dose-dependent manner.
3.4. Transcriptional Analysis of C. albicans Biofilm-Related Genes
In C. albicans, the hwp1 (hyphal wall protein 1) and ece1 (extent of cell elongation 1) are hyphal-specific genes, while als3 (agglutinin-like sequence 3) is an adhesion-related gene, all of which are involved in filamentation and biofilm formation [25]. In this experiment, act1 was used as the house-keeping gene, and untreated samples served as the control group. Accordingly, the transcriptional levels of hwp1, ece1, and als3 were analyzed in C. albicans using qRT-PCR upon the treatment of small-molecule chemicals. The results revealed that 1000 μg/mL IAA significantly decreased the mRNA levels of hwp1, als3, and ece1 by 62%, 60%, and 63%, respectively (Figure 5). Similarly, 200 μM of dodecanol was found to cause the downregulation of hwp1, ece1, and als3 transcripts by 76%, 82%, and 70%, respectively (Figure 5).
4. Conclusions
In sum, the results demonstrated that the S. pombe protein SPBPJ4664.02 is the homology of Flo11, a known adhesin in brewer’s yeast S. cerevisiae governing the cell adhesion to the abiotic surface of substrates, based on the SPBPJ4664.02 gene knock-out and complementation experimentation in combination with functional tests. S. pombe SPBPJ4664.02 is thereafter designated as a Flo11-like protein. Adhesion is the prerequisite for such morphological changes as filamentous growth, biofilm formation, or cell flocculation in yeasts. Analogous to Flo11 in S. cerevisiae, Flo11-like protein SPBPJ4664.02 was found to be required for biofilm formation as well as filamentation (evidenced by invasive growth into agar). Furthermore, our studies uncovered that the supplementation of small-molecule compounds at high concentrations such as 1000 μg/mL (IAA) or 1 mM (dodecanol) inhibited the biofilm formation in all three genotypes of S. pombe, namely, SPBPJ4664.02∆, SPBPJ4664.02∆/flo11^OE^ (SPBPJ4664.02 deficiency rescued by flo11 expression), as well as isogenic WT, though opposing effects were observed with IAA and dodecanol at low concentrations. IAA or dodecanol, as signaling molecules, exert regulatory actions probably at the downstream of Flo11-like adhesin SPBPJ4664.02 in the chemical signaling pathway for biofilm formation; however, they might act independently of Gsf2 (galactose-specific flocculin), which is indispensable for cell flocculation and thus biofilm formation. The regulatory roles of IAA or dodecanol are deduced from our investigation of SPBPJ4664.02Δ and SPBPJ4664.02Δ/flo11^OE^ versus isogenic WT, along with two other mutants, gsf2∆ and gas2∆ (deficient in the cell wall-remodeling enzyme), in both of which the biofilm-forming abilities were unaffected by the supplementation of physiologically relevant levels of IAA or dodecanol. Moreover, the impacts of IAA and dodecanol were further extrapolated to the medically important yeast C. albicans, which indicates that: (1) dodecanol had suppressive effects on biofilm formation at all concentrations tested herein; (2) IAA, administrated at high levels such as 200 and 1000 μg/mL, might inhibit biofilm formation, whereas the opposite effects were observed with IAA at low levels (12.5~100 μg/mL). Taken together, high concentrations of IAA or dodecanol could inhibit the biofilm formation in two different yeasts, S. pombe and C. albicans. Therefore, IAA and dodecanol have great potential to be developed as new anti-biofilm agents in synergy with antifungal drugs in the prevention and treatment of C. albicans infection, including relapsed and refractory candidiasis.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Willaert R.G. Kayacan Y. Devreese B. The Flo Adhesin Family Pathogens 202110139710.3390/pathogens 1011139734832553 PMC 8621652 · doi ↗ · pubmed ↗
- 2Kusakabe T. Lin W.Y. Cheong J.G. Singh G. Ravishankar A. Yeung S.T. Mesko M. De Celie M.B. Carriche G. Zhao Z. Fungal microbiota sustains lasting immune activation of neutrophils and their progenitors in severe COVID-19Nat. Immunol.2023241879188910.1038/s 41590-023-01637-437872315 PMC 10805066 · doi ↗ · pubmed ↗
- 3Amoah-Buahin E. Bone N. Armstrong J. Hyphal growth in the fission yeast Schizosaccharomyces pombe Eukaryot. Cell 200541287129710.1128/EC.4.7.1287-1297.200516002654 PMC 1168962 · doi ↗ · pubmed ↗
- 4Baillie G.S. Douglas L.J. Role of dimorphism in the development of Candida albicans biofilms J. Med. Microbiol.19994867167910.1099/00222615-48-7-67110403418 · doi ↗ · pubmed ↗
- 5Talapko J. JuzbašićM. MatijevićT. Pustijanac E. Bekic S. Kotris I. Skrlec I. Candida albicans—The virulence factors and clinical manifestations of infection J. Fungi 202177910.3390/jof 702007933499276 PMC 7912069 · doi ↗ · pubmed ↗
- 6Alnuaimi A.D. O’Brien-Simpson N.M. Reynolds E.C. Mc Cullough M.J. Clinical isolates and laboratory reference Candida species and strains have varying abilities to form biofilms FEMS Yeast Res.20131368969910.1111/1567-1364.1206823927631 · doi ↗ · pubmed ↗
- 7Bouyx C. Schiavone M. François J.M. FLO 11, a developmental gene conferring impressive adaptive plasticity to the yeast Saccharomyces cerevisiae Pathogens 202110150910.3390/pathogens 1011150934832664 PMC 8617999 · doi ↗ · pubmed ↗
- 8Brown G.D. Denning D.W. Gow N.A.R. Levitz S.M. Netea M.G. White T.C. Hidden killers: Human fungal infections Sci. Transl. Med.20124165 rv 1310.1126/scitranslmed.300440423253612 · doi ↗ · pubmed ↗