Genome-Wide Identification of the ABC Gene Family and Its Expression in Response to the Wood Degradation of Poplar in Trametes gibbosa
Jia Zhao, Achuan Wang, Qian Wang

TL;DR
This paper identifies and analyzes the ABC gene family in the wood-rotting fungus Trametes gibbosa, showing how these genes respond during wood degradation.
Contribution
The study provides the first genome-wide identification and expression analysis of the ABC gene family in T. gibbosa during wood degradation.
Findings
12 Tg-ABC genes were identified and classified into four subfamilies in Trametes gibbosa.
Tg-ABC genes show significant expression differences in woody versus non-woody environments.
Expression levels of Tg-ABC genes peak at 20 and 35 days during wood degradation.
Abstract
Wood-rotting fungi’s degradation of wood not only facilitates the eco-friendly treatment of organic materials, decreasing environmental pollution, but also supplies crucial components for producing biomass energy, thereby reducing dependence on fossil fuels. The ABC gene family, widely distributed in wood-rotting fungi, plays a crucial role in the metabolism of lignin, cellulose, and hemicellulose. Trametes gibbosa, as a representative species of wood-rotting fungi, exhibits robust capabilities in wood degradation. To investigate the function of the ABC gene family in wood degradation by T. gibbosa, we conducted a genome-wide analysis of T. gibbosa’s ABC gene family. We identified a total of 12 Tg-ABCs classified into four subfamilies (ABCA, ABCB, ABCC, and ABCG). These subfamilies likely play significant roles in wood degradation. Scaffold localization and collinearity analysis results…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
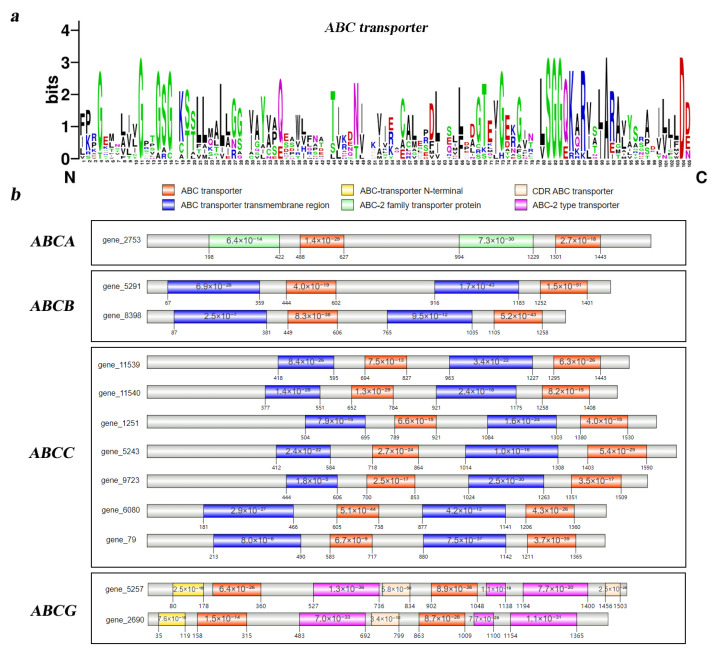 Figure 1
Figure 1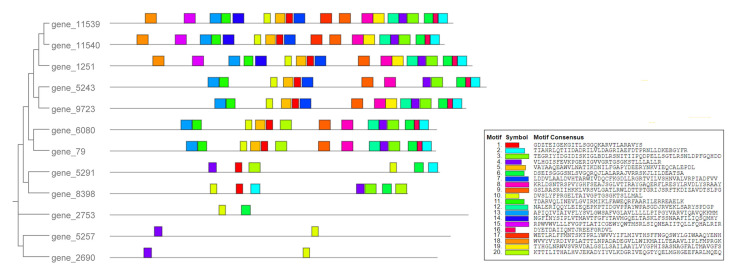 Figure 2
Figure 2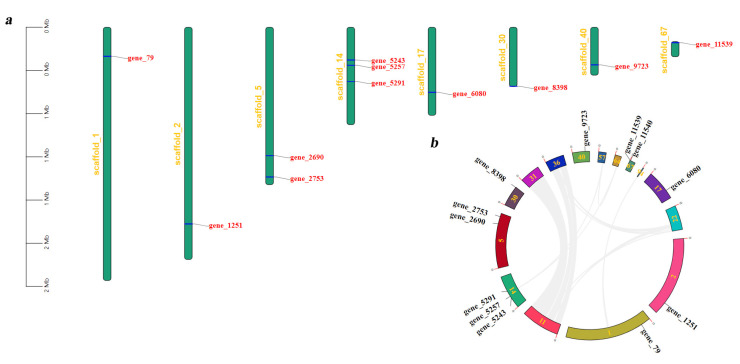 Figure 3
Figure 3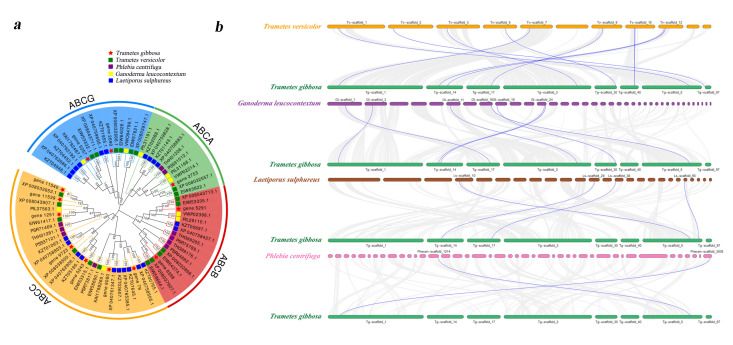 Figure 4
Figure 4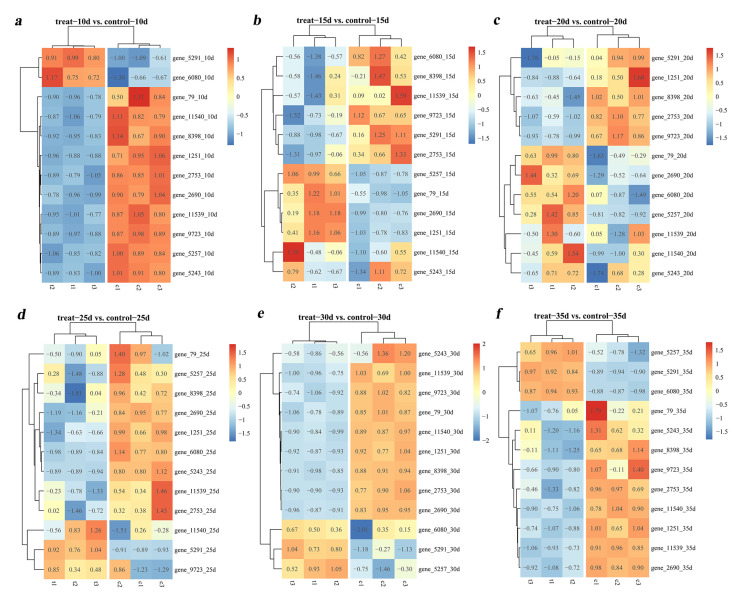 Figure 5
Figure 5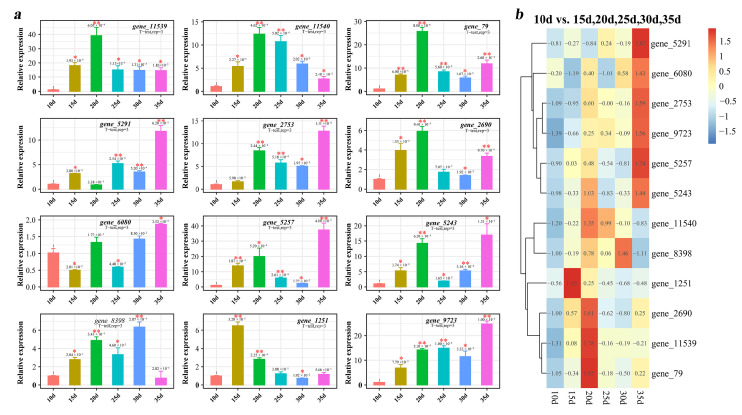 Figure 6
Figure 6Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsTannin, Tannase and Anticancer Activities · Enzyme-mediated dye degradation · Phytochemical compounds biological activities
1. Introduction
Poplars, known for their rapid growth, wide distribution, and short genome, among other advantages, have found extensive applications in various fields. Their fast growth makes their wood suitable for biomass energy production, and their lightweight, durable, and easy-to-process characteristics contribute to their widespread use in furniture manufacturing, construction, and the paper industry [1]. Populus simonii Carr × Populus nigra L., a hybrid variety of poplar, inherits favorable traits from both parent species, exhibiting excellent fast-growing properties and features such as drought resistance, cold resistance, and insect resistance. As a result, it has become a key afforestation species in northern regions [2,3,4]. Known for its rapid growth, it typically reaches commercial timber size in a few years. Its wood generally contains a moderate amount of cellulose, making it well-suited for paper and pulp production. Therefore, it has significant potential in areas such as timber production, the paper industry, and biomass energy production [5,6].
Wood degradation is the gradual breakdown of wood into smaller compounds and organic substances in natural surroundings or specific conditions. This process plays a crucial role in the organic material cycle, contributing to the preservation of ecological balance. Furthermore, it serves as a source of raw materials for biomass energy production, offering a means to reduce reliance on fossil fuels [7,8,9,10]. However, wood, consisting of intricate compounds like lignin, cellulose, and hemicellulose, encounters numerous challenges during degradation. Lignin, the most challenging component to degrade in wood, possesses a complex structure and resists degradation by most biological enzymes due to its multiple chemical bonds. Cellulose, which is relatively more susceptible to degradation than lignin, benefits from its linear structure, facilitating enzymes in breaking β-1,4-glucosidic bonds. Hemicellulose presents a degradation difficulty that falls between lignin and cellulose. These components intertwine, forming a robust and less easily destructible structure, thereby heightening the complexity of wood degradation [11,12,13].
Wood-rotting fungi excel at degrading wood, and wood decomposition constitutes their primary way of life. They efficiently break down wood and plant residues, converting organic matter into forms more readily absorbed by other organisms. This process contributes to the natural recycling of organic materials. Furthermore, these fungi can be employed to treat waste from the wood and pulp industry, reducing environmental pollution. They present a potential for low-energy, pollution-free wood degradation and sustainable energy production [14,15]. Trametes gibbosa, as one of numerous wood-rotting fungi, possesses significant wood-degrading capabilities. It can release extracellular enzymes such as manganese peroxidases, lignin peroxidase, and laccase, facilitating the degradation of lignin in wood. Additionally, it produces intracellular glucanases, extracellular glucanases, and xylanases, enabling the breakdown of cellulose and hemicellulose in wood. These enzymes assist T. gibbosa in breaking down wood into small molecular compounds, achieving wood degradation [16,17,18,19].
The ABC gene family is widely distributed among plants, insects, and microorganisms. Genes in this family encode ABC transporters, which play a pivotal role in the metabolism of lignin, cellulose, and hemicellulose [20,21]. ABC transporters facilitate the movement of the intermediates or degradation products of lignin and cellulose between intracellular and extracellular spaces. They can create channels on the cell membrane, transporting various degradation enzymes involved in breaking down lignin and cellulose from the intracellular space to the extracellular space or other cell structures, thereby enhancing degradation efficiency. Additionally, they can expel potentially toxic substances from the cell’s products into the external environment, maintaining the stability of the intracellular environment [22,23,24]. Researchers have conducted thorough investigations into the relationship between the ABC gene family and lignin and cellulose. For example, de Lima et al. found that the ABC gene (SvABCG17) is a potential transporter of lignin monomers [25]. Yu et al. suggested the potential role of the ABC gene (PgrABCG14) in promoting plant growth and lignin accumulation [26]. Xie et al. identified ABC family genes in Phanerochaete chryso-sporium and suggested that the transporter protein identified in the P. chrysosporium secretomes could play a role in cellobiose and/or cellodextrin uptake [27]. Based on these studies, it is speculated that the ABC gene family may have a significant impact on wood degradation. However, at present, no research has explored the specific functions of ABC family genes in wood degradation.
Here, we focused on the degradation process of P. simonii Carr × P. nigra L. wood by T. gibbosa, aiming to investigate the functions and responses of the ABC gene family, conducting analyses on its structure, predicting functions, and identifying characteristics. Additionally, we conducted a quantitative analysis of T. gibbosa’s ABC genes at different time points in both woody and non-woody environments using the RT-qPCR analysis, to elucidate the response of ABC gene family in the process of degrading poplar wood of T. gibbosa. This study establishes a solid foundation for wood-rotting fungi to contribute to the production of low-energy, environmentally friendly organic matter and sustainable energy.
2. Materials and Methods
2.1. Sample Preparation
Trametes gibbosa (Pers.) Fr. was provided by the Forest Pathology Laboratory of the Forestry Protection subject at the College of Forestry, Northeast Forestry University. The fungus was cultured in 250 mL conical flasks with 70 mL of LNAS medium (Low-Nitrogen Asparagine Succinic Acid) [28] and 5 mL of 15% glucose, with a pH value set at 4.5. The cultures were maintained at 27 °C under static conditions for 10, 15, 20, 25, 30, and 35 days. For the treatment group, 2 g samples of wood chips (1 × 1 × 5 cm) from Populus simonii Carr × Populus nigra L. were added to the conical flasks, while the control group had no wood chips. Each treatment condition had three biological replicate samples, and each sample was obtained by combining the mycelia from 5 flasks. After the removal of wood chips, the mycelia were collected and stored in liquid nitrogen for subsequent experimental use.
2.2. Identification of ABC Genes in T. gibbosa
Using the Pfam database “http://pfam.xfam.org (accessed on 5 September 2023)” [29], we conducted a search for DNA-binding domain sequences in the ABC genes of T. gibbosa (Tg-ABCs) (Table S1). Following this, the Tg-ABCs were classified into subfamilies using the CDD database “https://www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrpsb.cgi (accessed on 5 September 2023)” [30]. The DNA binding domain was subsequently visually compared using WebLogo “http://weblogo.berkeley.edu/logo.cgi (accessed on 6 September 2023)”.
2.3. Phylogenetic Relationship and Motif Analyses of Tg-ABCs in T. gibbosa
The protein sequences of the Tg-ABC gene family were aligned using MEGA 11.0.13 software [31], and a phylogenetic tree was constructed using the neighbor-joining (NJ) method (Figure S1a). To further verify the evolutionary relationship of each protein, we constructed a phylogenetic tree of Tg-ABC proteins using the optimal model (LG+G+F) with the maximum likelihood (ML) method (Figure S1b). MEME “http://meme-suite.org (accessed on 26 September 2023)” was used to identify conserved motifs in the Tg-ABC protein sequences. TBtools 2.008 [32] was used to visualize the phylogenetic tree and gene structures of Tg-ABCs.
2.4. Scaffold Localization and Collinearity Analysis of Tg-ABCs in T. gibbosa
The annotation of Tg-ABCs was acquired from JGI databases “https://mycocosm.jgi.doe.gov/Tragib1/Tragib1.home.html (accessed on 28 September 2023)”. Tg-ABCs were mapped to the genome [33] of T. gibbosa, and the duplication events in Tg-ABCs were analyzed using MCScanX in TBtools 2.008. Both of these results were visualized using TBtools 2.008.
2.5. Phylogenetic Relationship and Collinearity Analyses of ABC Gene Family
Segmental duplication events and collinearity between Tg-ABCs and homologous genes from four species of wood-rotting fungi (T. versicolor, G. leucocontextum, L. sulphureus and P. centrifuga) were analyzed using Dual Synteny Plotter. The protein sequences of these four species were obtained from the NCBI databases “https://www.ncbi.nlm.nih.gov/protein (accessed on 8 September 2023)”. A phylogenetic tree was constructed using the neighbor-joining (NJ) method in MEGA. The results were visualized in TBtools.
2.6. Other Characteristic Analyses in the Tg-ABCs
ProtParam “https://web.expasy.org/protparam (accessed on 16 September 2023)” was used to predict the physical and chemical properties of the Tg-ABC protein (Table 1). To examine the hydrophobicity of the Tg-ABC protein, we employed ExPASy ProtScale “https://web.expasy.org/protscale (accessed on 16 September 2023)” (Figure S2; Table S2). TMHMM “http://www.cbs.dtu.dk/services/TMHMM (accessed on 16 September 2023)" was employed to forecast transmembrane helices (Figure S3; Table S3). For the prediction of phosphorylation sites, Netphos “http://www.cbs.dtu.dk/services/NetPhos (accessed on 16 September 2023)” was used (Figure S4; Table S4). To anticipate the topological heterogeneity model of the Tg-ABC protein, Protter “http://wlab.ethz.ch/protter/start/ (accessed on 16 September 2023)” was employed (Figure S5; Table S5).
2.7. RT-qPCR Analysis of Tg-ABCs in Response to Wood Degradation by T. gibbosa
We separately extracted total RNA from T. gibbosa cultured under woody and non-woody environments for 10, 15, 20, 25, 30, and 35 days using the RNAprep Pure Plant Kit (DP441) from Tiangen Biotech (Beijing) Co., Ltd., Beijing, China and selected Gpd as the reference gene. Real-time quantitative reverse transcription PCR (RT-qPCR) was then employed to determine the expression levels of Tg-ABCs and the reference gene, and the RTq-PCR was performed following the protocol of a TaKaRa one-step RT-PCR kit from Baori Medical Technology (Beijing) Co., Ltd., Beijing, China. The primer sequences are provided in Table S6. The 2^−ΔΔCt^ [34] method was applied to calculate the relative expression levels of Tg-ABCs in woody environments compared to non-woody environments, as well as at different culture times relative to the 10-day culture.
3. Results
3.1. Identification of ABC Genes in T. gibbosa
We identified 12 members of the ABC gene family (Tg-ABCs) from the genome of T. gibbosa, all of which contain the conserved ABC transporter binding domain. Using WebLogo “https://weblogo.berkeley.edu/logo.cgi (accessed on 16 September 2023)”, we aligned the sequences of the conserved domains in the ABC transporter domains of the 12 Tg-ABCs (Supplementary Data S1), and the alignment results for 105 amino acids are depicted in Figure 1a. Each stack in the figure is composed of different amino acid symbols, and the overall height of the stack reflects the degree of sequence conservation at that position. The alignment results demonstrate a high conservation level of the ABC transporter domain sequences among the 12 Tg-ABCs. Specific amino acid positions, namely 11, 81, 82, 83, 91, and 104, exhibit complete amino acid identity, indicating excellent consistency in amino acid types at these positions.
The sequence alignment results for the DNA binding domain (Figure 1b) indicate that among the twelve Tg-ABCs, nine of them contain two conserved domains: the ABC transporter and the ABC transporter transmembrane region. Among these, seven belong to the ABCC subfamily, and two belong to the ABCB subfamily. However, in the twelve Tg-ABCs, three lack the ABC transporter transmembrane region. Of these genes, both gene5257 and gene2690 possess four conserved domains, placing them in the ABCG subfamily, while gene2753 features an ABC-2 family transporter distinct from other Tg-ABCs, categorizing it as part of the ABCA subfamily.
3.2. Phylogenetic Relationship and Motif Analyses of Tg-ABCs in T. gibbosa
To elucidate the relationships among members of the ABC gene family in T. gibbosa, we constructed an NJ-phylogenetic tree using their protein sequences (Figure S1) and identified 20 motifs within the 12 Tg-ABCs (Figure 2).
The results indicate that genes classified within the same ABC subfamily cluster on proximate branches, sharing similar motif types. Motif 10, found in all 12 Tg-ABCs, is characterized as the ABC transporter-like ATP-binding domain. Genes in the ABCC subfamily exhibit a higher number of motifs ranging from 13 to 19. Genes in the ABCB subfamily have six to seven motifs. The ABCA and ABCG subfamily genes contain the fewest motifs, with each subfamily having only two.
3.3. Scaffold Localization and Collinearity Analyses of Tg-ABCs in T. gibbosa
By analyzing the scaffold localization of the 12 Tg-ABCs, we discovered that only gene5243, gene5257, and gene5291 share the same location on scaffold14, and gene2690 and gene2753 are found together on scaffold5, while the rest of the genes are situated on distinct scaffolds individually. This finding indicates a dispersed distribution of Tg-ABCs within the scaffolds of T. gibbosa (Figure 3a).
We analyzed the segmental duplication events of Tg-ABCs in T. gibbosa and found no duplication of gene sequences. Additionally, genes located on the eight scaffolds (1, 2, 5, 14, 17, 30, 40, and 67) associated with Tg-ABCs displayed limited collinear gene pairs with genes on other scaffolds. In contrast, scaffolds without Tg-ABCs, specifically scaffold31 and scaffold36, exhibited higher numbers of collinear gene pairs with scaffold11 (Figure 3b). The integration of scaffold localization information and collinearity analysis results suggests that the functional roles of Tg-ABCs in T. gibbosa are diverse, potentially contributing differently to the process of wood degradation.
3.4. Physicochemical Analyses of Tg-ABCs in T. gibbosa
Physicochemical analyses (Table 1) revealed that the 12 Tg-ABC proteins in T. gibbosa share comparable lengths and molecular weights. The average amino acid length is 1521.5, and the average molecular weight is 167,619.4 Da, ranging from 145,732.30 to 185,461.04 Da. These proteins exhibit closely comparable lengths and molecular weights, displaying fluctuations around their respective averages.
Theoretical isoelectric points for Tg-ABC proteins range from 5.85 to 8.51, and the aliphatic index varies between 86.22 and 107.98. The average protein hydropathicity spans from −0.065 to 0.262, with the mean hydropathicity of five proteins being negative, classifying them as hydrophilic. In contrast, the remaining seven proteins exhibit positive predicted values, indicating their hydrophobic nature. These proteins are composed of five atoms, namely C, N, H, O, and S. Notably, three proteins (gene_5291, gene_11540, and gene_9723) have an instability index exceeding 40, suggesting structural instability, while the majority of proteins have an index below 40, indicating stable structures.
3.5. Phylogenetic Relationship and Collinearity Analyses of ABC Gene Family
To elucidate the similarity between Tg-ABCs and homologous genes in other species, we selected four wood-rotting fungi (T. versicolor, G. leucocontextum, L. sulphureus, and P. centrifuga) with comparable subfamily classifications of ABC genes to T. gibbosa. Subsequently, we screened the protein sequences of ABC subfamilies (ABCA, ABCB, ABCC, and ABCG) from each of these fungi (Supplementary Data S2) and constructed a collinearity relationship between T. gibbosa and each of the other species (Figure 4).
We conducted a phylogenetic analysis of T. gibbosa and four wood-rotting fungi (Figure 4a). We observed that in the phylogenetic trees of each ABC subfamily, the ABC genes of T. gibbosa and those of its same-genus fungus (T. versicolor) are positioned on relatively close branches. Additionally, the branches of ABC genes within the same subfamily are closer than those within the same species, indicating a higher degree of homology within the same ABC subfamily across different species. This suggests that ABC protein sequences exhibit a higher level of conservation during the evolutionary process.
We individually constructed collinearity relationships for T. gibbosa and four wood-rotting fungi (Figure 4b). In the collinearity analysis between T. gibbosa and its same-genus fungus (T. versicolor), we observed that among the twelve Tg-ABCs, nine exhibited collinearity, with seven, four, and two Tg-ABCs displaying collinearity with the other three species, respectively. Integrating the results from the phylogenetic and collinearity analyses, it becomes evident that T. gibbosa shares a closer phylogenetic relationship with T. versicolor, followed by G. leucocontextum, L. sulphureus, and P. centrifuga in descending order of phylogenetic proximity.
3.6. Quantitative Analysis of Tg-ABCs in T. gibbosa
To investigate the response of T. gibbosa’s Tg-ABCs to woody and non-woody environments, we conducted a comparative analysis, examining the relative expression levels of Tg-ABCs at different culture times in these distinct conditions (Figure 5). The results reveal that regardless of the cultivation duration, there are significant differences in the expression levels of Tg-ABCs in woody and non-woody environments. Specifically, at 10 days, the majority of genes exhibited noticeable downregulation, while the upward trends of other genes were also evident, indicating the initiation of an interaction between T. gibbosa and wood. This suggests the involvement of Tg-ABCs in the wood degradation process. Between days 15, 20, and 25, there were highly significant trends of both upregulation and downregulation for all 12 Tg-ABCs, with peak values observed during this period. This signifies that wood degradation is most strongly influenced by Tg-ABCs during this timeframe. At day 30, a relatively calm period ensues, with minimal changes in the expression levels of Tg-ABCs, suggesting a potentially minor impact of Tg-ABCs on wood degradation during this stage. However, at day 35, the previous calm period is disrupted, and new fluctuations in the expression levels of Tg-ABCs emerge, indicating a renewed and more significant impact of Tg-ABCs on wood degradation at this stage.
To further investigate the changes in the relative expression levels of Tg-ABCs in response to gradients of T. gibbosa’s wood degradation time, we analyzed the relative expression levels of Tg-ABCs cultivated in woody environments for 10, 15, 20, 25, 30, and 35 days (Figure 6). The results reveal that during a cultivation period of T. gibbosa in a woody environment lasting from 10 to 35 days, the relative expression levels of most Tg-ABCs followed a trend of increasing, decreasing, and increasing, with peak values observed at 20 and 35 days. At these time points, Tg-ABCs may have the most significant impact on T. gibbosa’s wood degradation. The relative expression levels of gene11539 and gene11540 showed a trend of increasing and then decreasing, with one fewer increase compared to most genes. Gene8398 demonstrated a trend of increasing, decreasing, increasing, and then decreasing again, featuring one additional decrease compared to most genes. On the other hand, gene5291 exhibited one additional increase compared to gene8398. As for gene6080, its expression had an extra decrease in the early stage compared to the majority of genes. These variations in expression trends may stem from differences in the functions of distinct genes, and different ABC subfamily genes may also play specific roles in wood degradation.
4. Discussion
The investigation of wood degradation holds substantial ecological and biological importance. Efficient wood degradation is vital for carbon cycling in ecosystems and provides essential technical support for sustainable biofuel production [35,36]. The ABC gene family plays a crucial role in lignin, cellulose, and hemicellulose metabolism, regulating vital substrate transport during this process [37]. T. gibbosa, as one of the numerous wood-rotting fungi, demonstrates efficient wood degradation capabilities [38], presenting a promising opportunity to investigate the role of the ABC gene family in wood degradation.
We identified a total of 12 Tg-ABCs by analyzing the genomic sequence of the ABC gene family in T. gibbosa. The results of the DNA binding domain alignment reveal that all of these genes contain conserved ABC transporter domains, exhibiting a high degree of sequence conservation. Tg-ABCs are classified into four subfamilies, ABCA, ABCB, ABCC, and ABCG. These subfamilies likely play significant roles in wood degradation, while the absence of five subfamilies [39] may have a relatively minor impact on this process. Scaffold localization results indicate that Tg-ABCs are dispersed on scaffolds. A collinearity analysis reveals that there is no duplication of gene sequences within the Tg-ABCs in the genome sequence of T. gibbosa. These findings collectively suggest that the functions of T. gibbosa’s ABC gene family in the wood degradation process may exhibit diversity. Phylogenetic and collinearity analyses of T. gibbosa with four other wood-rotting fungi show that T. gibbosa shares a closer phylogenetic relationship with its same-genus fungus (T. versicolor) [40], followed by G. leucocontextum, L. sulphureus, and P. centrifuga in descending order of phylogenetic proximity. Additionally, the branches of ABC genes within the same subfamily are closer than those within the same species, indicating a higher degree of homology within the same ABC subfamily across different species. This suggests that ABC protein sequences exhibit a higher level of conservation during the evolutionary process, and their wood degradation characteristics may also show a higher degree of similarity [41].
To gain a deeper understanding of how Tg-ABCs response to wood degradation by T. gibbosa, we conducted quantitative analyses of the expression of Tg-ABCs from T. gibbosa cultivated in both woody and non-woody environments for 10, 15, 20, 25, 30, and 35 days using an RT-qPCR analysis. The results reveal a significant difference in the expression levels of Tg-ABCs between woody and non-woody environments, suggesting an active involvement of the ABC gene family in wood degradation [21,42,43]. During the wood degradation period of T. gibbosa, spanning from 10 to 35 days, the relative expression levels of most Tg-ABCs exhibited a trend of increasing, decreasing, and then increasing again. It is speculated that this trend is associated with the different products generated during various stages of wood degradation by T. gibbosa. In the early stages of wood degradation, T. gibbosa produces a significant amount of extracellular enzymes, and ABC transporters facilitate the transport of these enzymes from the extracellular space into the cell, leading to an increase in their expression levels. In the mid-phase of wood degradation, these enzymes catalyze the oxidation and decomposition of large molecules such as lignin, cellulose, and hemicellulose. At this stage, ABC transporters are not required, resulting in a decrease in their expression levels. In the later stages of wood degradation, the generation of small molecular substances like acids, alcohols, and esters prompts ABC genes to transport these substances, causing a resurgence in their expression levels [16,44,45]. The fluctuation in the relative expression levels of Tg-ABCs corresponds to the metabolic trajectory of wood degradation, indicating the pivotal role of Tg-ABCs in the process. Additionally, at 20 and 35 days of wood degradation by T. gibbosa, the relative expression levels of Tg-ABCs peak, suggesting that at these time points, Tg-ABCs exert the most significant impact on wood degradation by T. gibbosa.
5. Conclusions
In this study, we systematically analyzed the ABC gene family in T. gibbosa, examining its structure, predicting its functions, and identifying the characteristics of Tg-ABCs. Additionally, we also explored how Tg-ABCs respond to woody environments. Our findings revealed that the ABC gene family plays a crucial role in the degradation of poplar wood by T. gibbosa. This study lays the foundation for a deeper understanding of the wood degradation mechanism of the ABC gene family and provides strong support for the development of more efficient strategies for the biological treatment of wood and other organic materials.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Thakur A.K. Kumar P. Parmar N. Shandil R.K. Aggarwal G. Gaur A. Srivastava D.K. Achievements and prospects of genetic engineering in poplar: A review New For.20215288992010.1007/s 11056-021-09836-3 · doi ↗
- 2Gao S. Li C. Chen X. Li S. Liang N. Wang H. Zhan Y. Zeng F. Basic helix-loop-helix transcription factor Pxb HLH 02 enhances drought tolerance in Populus (Populus simonii × P. nigra)Tree Physiol.20234318520210.1093/treephys/tpac 10736054366 · doi ↗ · pubmed ↗
- 3Tao J. Dong F. Wang Y. Chen H. Tang M. Arbuscular mycorrhizal fungi enhance photosynthesis and drought tolerance by regulating MAPK genes expressions of Populus simonii × P. nigra Physiol. Plant.2022174 e 1382910.1111/ppl.1382936437546 · doi ↗ · pubmed ↗
- 4Zhou B. Kang Y. Leng J. Xu Q. Genome-Wide Analysis of the mi RNA-m RN As Network Involved in Cold Tolerance in Populus simonii × P. nigra Genes 20191043010.3390/genes 1006043031195761 PMC 6627750 · doi ↗ · pubmed ↗
- 5Ahmed A.K.M. Fu Z. Ding C. Jiang L. Han X. Yang A. Ma Y. Zhao X. Growth and wood properties of a 38-year-old Populus simonii × P. nigra plantation established with different densities in semi-arid areas of northeastern China J. For. Res.20203149750610.1007/s 11676-019-00887-z · doi ↗
- 6Liu D. Liu M. Li Z. Wang G. Li Y. Zheng M. Liu G. Zhao X. Variation Analysis of Growth Traits of Transgenic Populus simonii x P.nigra Clones Carrying Ta LEA Gene Bull. Bot. Res.201535540546
- 7de Vries L. Guevara-Rozo S. Cho M. Liu L.Y. Renneckar S. Mansfield S.D. Tailoring renewable materials via plant biotechnology Biotechnol. Biofuels 20211416710.1186/s 13068-021-02010-z 34353358 PMC 8344217 · doi ↗ · pubmed ↗
- 8Vanholme R. De Meester B. Ralph J. Boerjan W. Lignin biosynthesis and its integration into metabolism Curr. Opin. Biotechnol.20195623023910.1016/j.copbio.2019.02.01830913460 · doi ↗ · pubmed ↗