Diverse modes of chromosome terminal deletion in spontaneous canavanine-resistant Schizosaccharomyces pombe mutants
Xiao-Hui Lyu, Fang Suo, Wen Li, Guo-Song Jia, Yu-Sheng Yang, Li-Lin Du

TL;DR
This study explores how chromosome deletions in fission yeast lead to canavanine resistance and reveals different mechanisms behind these deletions.
Contribution
The paper identifies three distinct mechanisms of chromosome terminal deletion in canavanine-resistant fission yeast mutants.
Findings
Three mutants, including can2-1, had terminal deletions of the left arm of chromosome II.
Deletions occurred via homology-driven translocation, homology-independent fusion, and de novo telomere addition.
These deletions caused loss of amino acid transporter genes, contributing to canavanine resistance.
Abstract
Canavanine resistance has been used to analyze mutation rates in the fission yeast Schizosaccharomyces pombe . However, the genetic basis of canavanine resistance in this organism remains incompletely understood. Here, we performed whole genome sequencing on five spontaneously arising canavanine-resistant S. pombe mutants, including the can2-1 mutant isolated in the 1970s. This analysis revealed that three mutants, including can2-1 , experienced terminal deletions of the left arm of chromosome II, leading to the loss of multiple amino acid transporter genes. Interestingly, these three mutants underwent chromosome terminal deletion through distinct mechanisms, including homology-driven translocation, homology-independent chromosome fusion, and de novo telomere addition. Our findings shed new light on the genetic basis of canavanine resistance and mechanisms underlying chromosome terminal…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
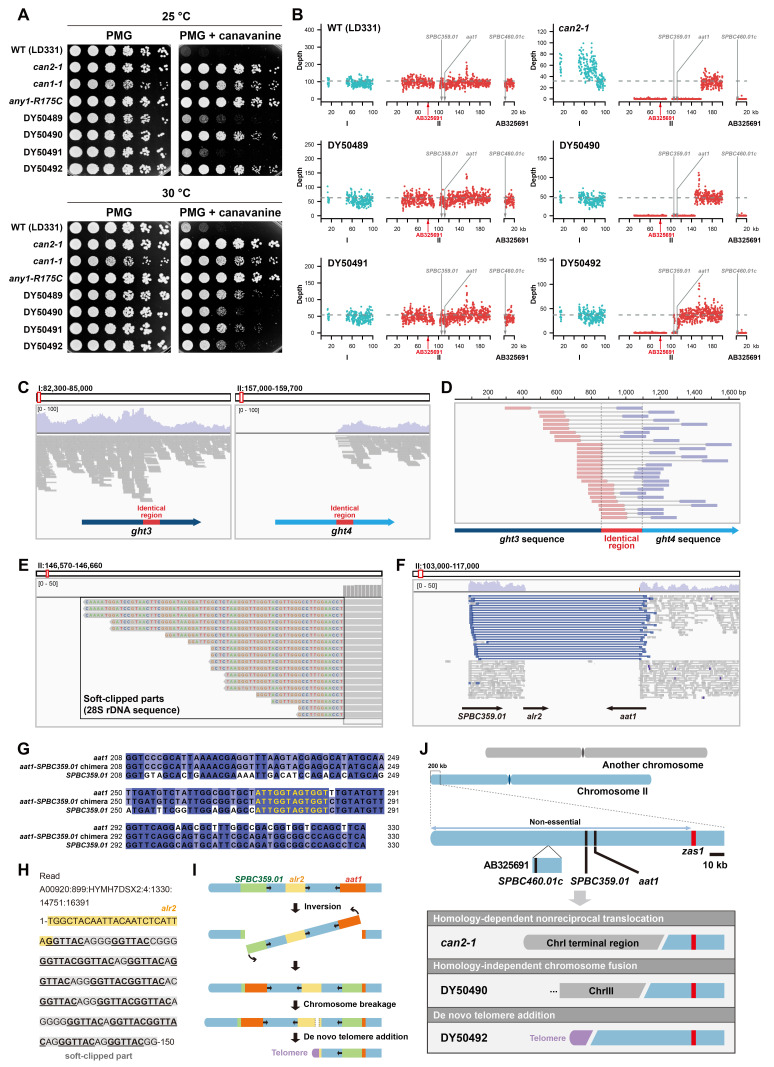 Figure 1
Figure 1|
Strains |
Genotype |
Original strain name |
Original source |
NBRP ID |
|
DY7144 |
|
Kohli 10-389 |
Jürg Kohli |
FY18665 |
|
DY49525 |
|
Kohli 10-391 |
Jürg Kohli |
FY18666 |
|
LD331 |
|
– |
Li-Lin Du |
– |
|
DY50489 |
|
– |
Xiao-Hui Lyu |
– |
|
DY50490 |
|
– |
Xiao-Hui Lyu |
– |
|
DY50491 |
|
– |
Xiao-Hui Lyu |
– |
|
DY50492 |
|
– |
Xiao-Hui Lyu |
– |
|
DY50493 |
|
– |
Xiao-Hui Lyu |
– |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsFungal and yeast genetics research · Polyamine Metabolism and Applications · Ubiquitin and proteasome pathways
Description
Canavanine, an arginine analog, can enter cells through amino acid transporters (Grenson et al., 1966; Fantes and Creanor, 1984) . Once inside cells, canavanine becomes incorporated into proteins, impairing their function and causing cytotoxicity (Rosenthal, 1991) . In the budding yeast Saccharomyces cerevisiae , spontaneously arising canavanine-resistant mutations are exclusively those disabling the arginine transporter gene CAN1 (Whelan et al., 1979) . Since CAN1 is located in a terminal, nonessential region of chromosome V, canavanine resistance can result from terminal deletion of this chromosome—a type of gross chromosomal rearrangement (GCR) (Chen et al., 1998; Chen and Kolodner, 1999) . The well-defined genetic basis of canavanine resistance in S. cerevisiae has led to the widespread use of canavanine resistance as an assay to analyze the rates and spectra of both small mutations and GCRs in this organism (Lang and Murray, 2008; Putnam and Kolodner, 2017; Jiang et al., 2021) .
Canavanine resistance has also been used to analyze mutation rates in the fission yeast Schizosaccharomyces *pombe * (Kaur et al., 1999; Tanaka et al., 2004; Sabatinos et al., 2013) . However, unlike in S. cerevisiae , the genetic basis of canavanine resistance in S. pombe is not fully understood. The first reported canavanine-resistant mutants in S. pombe are two spontaneous mutants can1-1 and can2-1 isolated in the 1970s (Kohli et al., 1977; Fantes and Creanor, 1984; Jürg Kohli, personal communication). Recently, the can1-1 mutation was identified as a gain-of-function mutation (R175C) in the * any1 * gene, which encodes a ubiquitin ligase that negatively regulates amino acid transporters (Yang et al., 2022; Saada et al., 2022; Pai et al., 2023) . Several other S. pombe genes, including * tsc1 * , * tsc2 * , * pas1 * , and * cat1 * , have been reported to cause canavanine resistance when individually deleted (van Slegtenhorst et al., 2004, 2005; Aspuria and Tamanoi, 2008) . However, it is unclear to what extent mutations in these genes are responsible for spontaneous canavanine resistance in S. pombe . The identity of the
- can2-1* mutation has remained unknown.
During our investigation of the can1-1 mutation (Yang et al., 2022) , we isolated four spontaneous canavanine-resistant mutants. They were derived from a wild-type S. pombe strain LD331. Among these four mutants, DY50489 and DY50491 showed stronger canavanine resistance at 30 °C than at 25 °C, while DY50490 and DY50492 exhibited stronger resistance at 25 °C than at 30 °C ( Figure 1A ). As a control, an *any1-R175C * knock-in mutant constructed in the LD331 background displayed greater canavanine resistance at both temperatures compared to the four mutants. A can1-1 strain and a *can2-1 * strain deposited by Jürg Kohli at the Yeast Genetic Resource Center of Japan (YGRC/NBRP) also showed canavanine resistance as expected.
We performed next-generation sequencing on the can2-1 strain and the four LD331-derived canavanine-resistant mutants. No mutations were found in * any1 * , * tsc1 * , * tsc2 * , * pas1 * , and * cat1 * . Remarkably, the can2-1 strain, DY50490, and DY50492 each harbored a >100-kb terminal deletion on the left arm of chromosome II ( Figure 1B ). Even though the deletion junctions fall at different positions in these three strains, the deletions all result in the loss of more than one amino acid transporter gene. In the case of the can2-1 strain and DY50490, three amino acid transporter genes
- SPBC460.01c* ,
SPBC359.01 * , * aat1 * are lost ( Figure 1B ). In the case of DY50492, due to a recombination between * SPBC359.01 * and * aat1 * , no intact SPBC460.01c , * SPBC359.01 * , or * aat1 * remains and an * aat1 *
SPBC359.01 * chimera is present near a newly formed telomere and thus may be silenced (see more details below). The deletion of * aat1 * is known to cause a reduction of arginine uptake by more than 50% (Nakase et al., 2012) . SPBC460.01c and * SPBC359.01 * encode amino acid transporters closely related to Aat1 (Yang et al., 2022) . It is likely that the simultaneous loss of function of these amino acid transporter genes is the underlying cause of the canavanine resistance phenotype of these three strains. This conclusion is consistent with previous assignment of the *can2 * locus to chromosome II by genetic mapping (Kohli et al., 1977) . The only mutation found in DY50489 was a missense mutation (A535T) in * rpc1 * that encodes a subunit of RNA polymerase III, while the only mutation found in DY50491 was a missense mutation (P308S) in * drs1 * that encodes aspartyl-tRNA synthetase. It remains to be determined whether these two mutations are related to the canavanine resistance phenotype.
We next investigated the mechanisms of chromosome II terminal deletions in the can2-1 strain, DY50490, and DY50492 by inspecting the next-generation sequencing reads. The deletion junction in the can2-1 strain is located in the middle of the * ght4 * gene on chromosome II ( Figure 1C ). Interestingly, read depth analysis revealed a duplication of the left arm terminal sequence of chromosome I in this strain ( Figure 1B ). The duplication ends in the middle of the * ght3 * gene ( Figure 1C ). * ght3 * and * ght4 * encode closely related hexose transporters (84% overall amino acid identity) (Heiland et al., 2000) . Moreover, a 240-bp sequence in these two genes share 100% nucleotide identity (codons 286–365, denoted as the “identical region” in Figures 1C and 1D). Because these two genes are both oriented in a telomere-to-centromere direction ( Figure 1C ), a nonreciprocal translocation occurring through homologous recombination between their identical regions could explain the terminal deletion on chromosome II and the terminal duplication on chromosome I. In support of this scenario, we identified read pairs that map to a ght3
ght4 chimera, but not to * ght3 * or * ght4 * ( Figure 1D ). Collectively, these findings indicate that the chromosome II terminal deletion in the can2-1 strain results from a homology-driven unbalanced translocation.
The deletion junction of DY50490 is located in the intergenic region between SPBC1683.04 and * thi7 * . Soft-clipped reads at the deletion junction indicate that the first undeleted nucleotide is at coordinate 146651 of chromosome II ( Figure 1E ). Strikingly, the soft-clipped parts of these reads match the sequence of the 28S rDNA gene, which normally resides in the two rDNA arrays located at the two ends of chromosome III ( Figure 1E ). The rDNA nucleotide immediately next to the first undeleted nucleotide of chromosome II is at position 2028 of the annotated 28S rDNA gene (GenBank accession Z19578.1). In the soft-clipped reads, the rDNA sequence is oriented with its telomere-proximal side facing the deletion junction, suggesting that chromosome II and chromosome III have fused at this junction. No microhomology was observed between the chromosome II sequence and the rDNA sequence flanking the fusion breakpoint. Thus, the chromosome II terminal deletion in DY50490 is likely due to a homology-independent chromosome fusion event. Because the dicentric chromosome resulting from such a fusion event is incompatible with cell viability, the fusion event presumably has been followed by either breakage of the fused chromosome or epigenetic inactivation of one of the two centromeres in the fused chromosome (Sato et al., 2012; Gu et al., 2022) .
Based on the read mapping result, DY50492 has undergone sequence loss in two regions: a chromosome II left arm terminal region telomeric to the amino acid transporter gene * SPBC359.01 * and an approximately 5-kb region centromeric to * SPBC359.01 * ( Figure 1F ). Discordant read pairs revealed that this dual loss pattern probably results from an inversion event followed by a chromosome terminal deletion event ( Figure 1F ). The two inversion junctions fall within * SPBC359.01 * and * aat1 * , which are homologous amino acid transporter genes with opposite orientations. Alignment of the coding sequences of * aat1 * , * SPBC359.01 * and the aat1
SPBC359.01 chimera in DY50492 narrowed the inversion breakpoint to a 12-bp sequence identical between * SPBC359.01 * and * aat1 * ( Figure 1G ). The chromosome terminal deletion junction in DY50492 is located in the middle of * alr2 * , the gene situated between * SPBC359.01 * and * aat1 * . The first undeleted nucleotide is at the 62nd position of the coding sequence of * alr2 * . In the soft-clipped reads mapped to the deletion junction, the soft-clipped parts contain tandem copies of the core telomeric repeat sequence GGTTAC ( Figure 1H ) (Ares and Chakrabarti, 2008) , indicating that the deletion has been healed by de novo telomere addition. A schematic depicting the inversion and deletion events is shown in Figure 1I . We suspect that the aat1
SPBC359.01 chimera in DY50492 may not be functional, because its close proximity to the telomeric repeats may result in its silencing.
Our findings indicate that the loss of function of three amino acid transporter genes ( SPBC460.01c , * SPBC359.01 * , and * aat1 * ) through chromosome II left arm terminal deletion may account for a significant fraction of spontaneous canavanine-resistant mutants in S. pombe . It is remarkable that the three chromosome terminal deletion mutants that we analyzed ( can2-1 , DY50490, and DY50492) involve three distinct deletion mechanisms ( Figure 1J ). On the left arm of chromosome II, the most telomeric essential gene is * zas1 * (Sasaki et al., 2013) , which is located about 71 kb centromeric to * SPBC359.01 * ( Figure 1J ). Assuming that canavanine resistance requires the deletion junction to be on the centromeric side of * SPBC359.01 * so that SPBC460.01c and * SPBC359.01 * are lost and * aat1 * is either lost or inactivated by telomeric silencing, but not too close (< 5 kb) to * zas1 * to avoid its silencing, the deletion junction can fall anywhere within an approximately 66 kb region. The large size of this region may explain the abundance of chromosome II left arm terminal deletion mutants among spontaneous canavanine-resistant mutants and the diverse mechanisms underlying the deletions. Further study of spontaneous canavanine-resistant S. pombe mutants may provide additional insights into the mechanisms of chromosomal rearrangement.
Methods
**Isolation of spontaneously arising canavanine-resistant mutants: ** In our previous study (Yang et al., 2022) , we conducted any1-R175C knock-in by transforming a wild-type strain (LD331) with an any1-R175C PCR product and selecting canavanine-resistant clones. A no-DNA control transformation also yielded a small number of canavanine-resistant clones on PMG plates supplemented with 5 μg/mL canavanine. Four canavanine-resistant clones from this no-DNA control transformation were saved as DY50489–DY50492.
Spot assay: Cells were cultured to log phase in the YES medium and harvested by centrifugation. Cell pellets were washed three times with sterile water and resuspended to an OD 600 of 0.4. 200 μL of each cell suspension was added to a 96-well plate and serial five-fold dilutions of each strain were prepared. Cell suspensions were spotted onto PMG plates with or without 5 μg/ml of canavanine using a pin tool and cultured at 30 °C for 108 hours. Images of plates were obtained using an Epson Perfection V800 Photo scanner.
Whole-genome sequencing: Illumina sequencing libraries were constructed using home-made Tn5 transposase as described previously (Tao et al., 2019) . Paired-end Illumina sequencing (2 x 150 bp) was performed on the Illumina NovaSeq 6000 system at Novogene (Beijing, China). The sequencing data have been deposited in the NCBI Short Read Archive (SRA) under BioProject accession PRJNA1062442. The sequencing data of LD331 was deposited under the strain name DY49197.
Read mapping: Illumina sequencing reads were cleaned using fastp (version 0.20.0) (Chen et al., 2018) and then mapped onto the reference genome of S. pombe using BWA-MEM (v0.7.17-r1188) (Li and Durbin, 2009) . Mapped reads were visualized using the Integrative Genomics Viewer (IGV) (v2.16.1) (Robinson et al., 2011) .
Read-depth analysis : Mappability was calculated by running GenMap version 1.3.0 using 100 bp k-mers with up to 2 mismatches (Pockrandt et al., 2020) . Genomic regions larger than 1 kb, where every position has a mappability value of one, were considered unique regions. For these regions, we plotted the average read depth in 200-bp sliding window calculated using samtools depth (version1.14) (Li et al., 2009) .
Reagents
**: **
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ares M Jr Chakrabarti K 200811 Stuttering against marginotomy.Nat Struct Mol Biol 1511545-9993181910.1038/nsmb 0108-1818176550 PMC 2577027 · doi ↗ · pubmed ↗
- 2Aspuria PJ Tamanoi F 200851 The Tsc/Rheb signaling pathway controls basic amino acid uptake via the Cat 1 permease in fission yeast.Mol Genet Genomics 27951617-461544145010.1007/s 00438-008-0320-y 18219492 PMC 2670428 · doi ↗ · pubmed ↗
- 3Chen C Kolodner RD 199991 Gross chromosomal rearrangements in Saccharomyces cerevisiae replication and recombination defective mutants.Nat Genet 2311061-4036818510.1038/1268710471504 · doi ↗ · pubmed ↗
- 4Chen C Umezu K Kolodner RD 199871 Chromosomal rearrangements occur in S. cerevisiae rfa 1 mutator mutants due to mutagenic lesions processed by double-strand-break repair.Mol Cell 211097-276592210.1016/s 1097-2765(00)80109-49702187 · doi ↗ · pubmed ↗
- 5Chen S Zhou Y Chen Y Gu J 201891 fastp: an ultra-fast all-in-one FASTQ preprocessor.Bioinformatics 34171367-4803 i 884i 89010.1093/bioinformatics/bty 56030423086 PMC 6129281 · doi ↗ · pubmed ↗
- 6Fantes PA Creanor J 1984121 Canavanine resistance and the mechanism of arginine uptake in the fission yeast Schizosaccharomyces pombe.J Gen Microbiol 130120022-12873265327310.1099/00221287-130-12-326518357653 · doi ↗ · pubmed ↗
- 7Grenson M Mousset M Wiame JM Bechet J 19661031 Multiplicity of the amino acid permeases in Saccharomyces cerevisiae. I. Evidence for a specific arginine-transporting system.Biochim Biophys Acta 12720006-300232533810.1016/0304-4165(66)90387-45964977 · doi ↗ · pubmed ↗
- 8Gu X Ye T Zhang XR Nie L Wang H Li W Lu R Fu C Du LL Zhou JQ 2022823 Single-chromosome fission yeast models reveal the configuration robustness of a functional genome.Cell Rep 40811123711123710.1016/j.celrep.2022.11123736001961 · doi ↗ · pubmed ↗