Mutations upstream from sdaC and malT in Escherichia coli uncover a complex interplay between the cAMP receptor protein and different sigma factors
Pernille Ott Frendorf, Sophia A. H. Heyde, Morten H. H. Nørholm

TL;DR
The paper explores how mutations in Escherichia coli affect gene regulation, revealing new insights into how bacteria control gene expression during different growth phases.
Contribution
The study reveals new insights into the interplay between the cAMP receptor protein and sigma factors in bacterial promoters.
Findings
Mutations in Escherichia coli lead to cAMP-independent regulation of malT and sdaC genes.
The location of the cAMP receptor protein binding site affects sigma factor usage in gene regulation.
The research provides new understanding of bacterial promoter architecture and gene regulation during growth phases.
Abstract
In Escherichia coli, one of the best understood microorganisms, much can still be learned about the basic interactions between transcription factors and promoters. When a cAMP-deficient cya mutant is supplied with maltose as the main carbon source, mutations develop upstream from the two genes malT and sdaC. Here, we explore the regulation of the two promoters, using fluorescence-based genetic reporters in combination with both spontaneously evolved and systematically engineered cis-acting mutations. We show that in the cya mutant, regulation of malT and sdaC evolves toward cAMP-independence and increased expression in the stationary phase. Furthermore, we show that the location of the cAMP receptor protein (Crp) binding site upstream of malT is important for alternative sigma factor usage. This provides new insights into the architecture of bacterial promoters and the global interplay…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
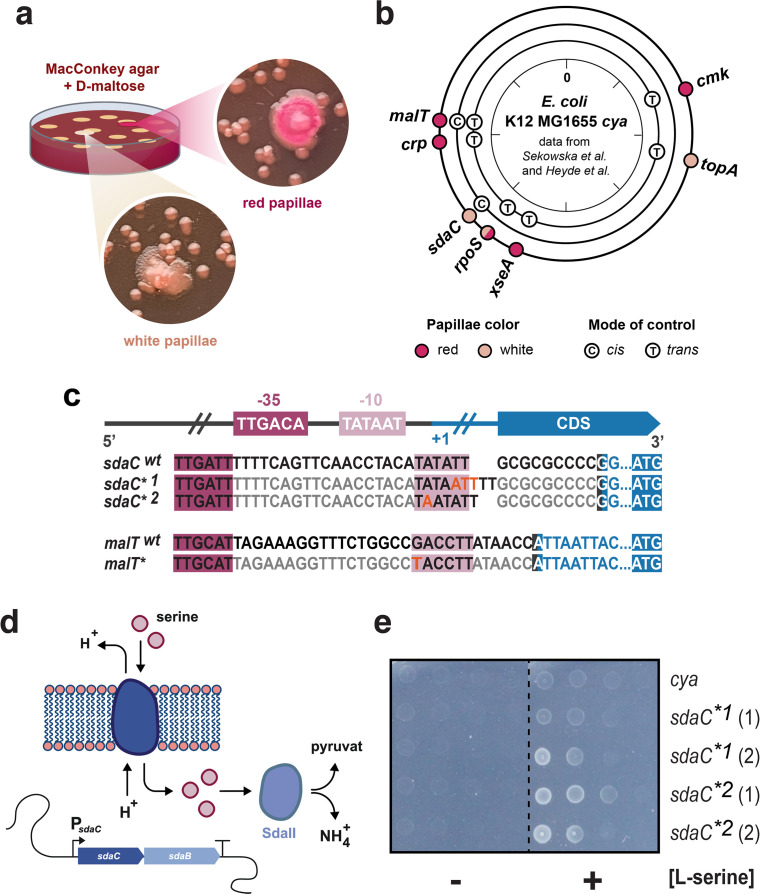 Fig 1
Fig 1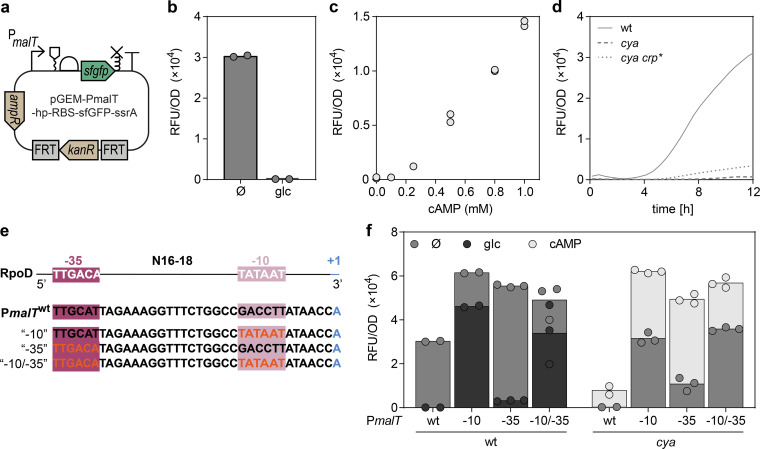 Fig 2
Fig 2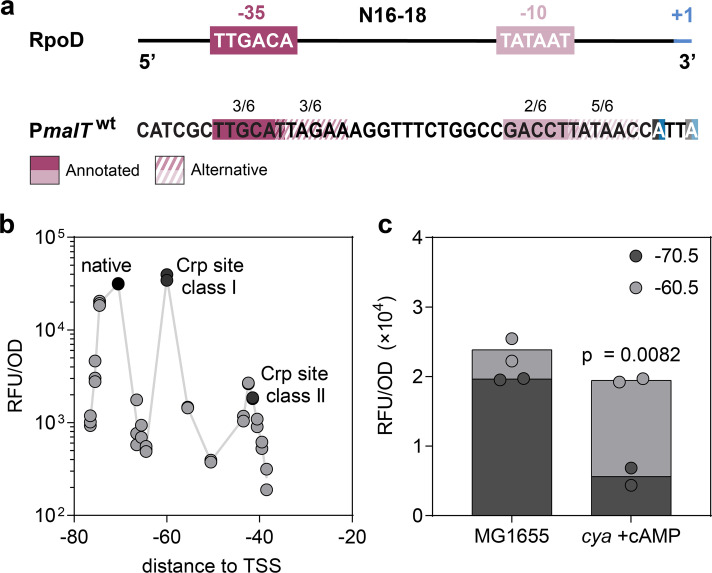 Fig 3
Fig 3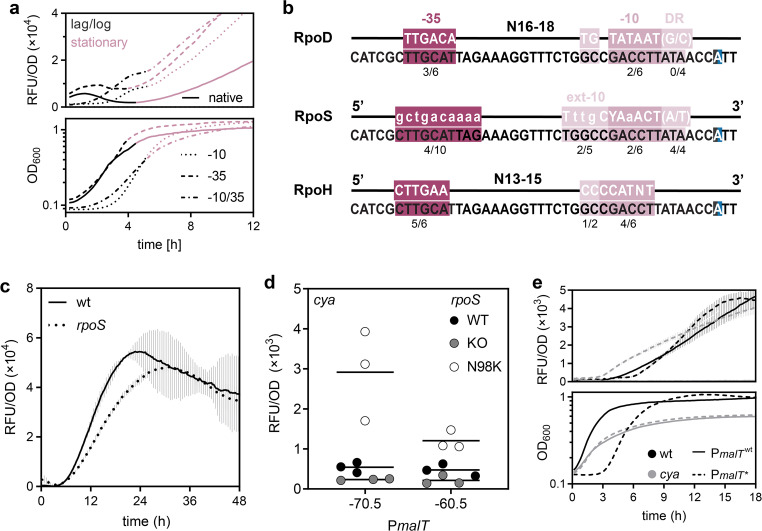 Fig 4
Fig 4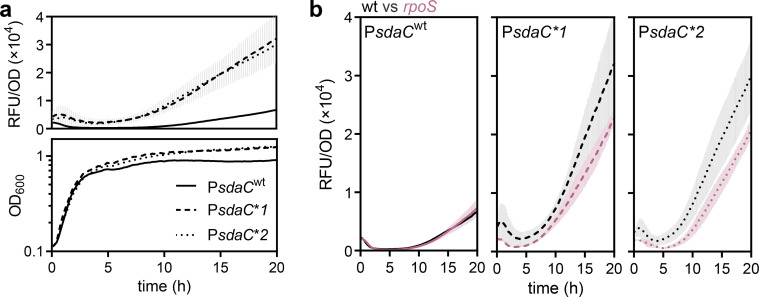 Fig 5
Fig 5- —Novo Nordisk Fonden (NNF)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsTuberculosis Research and Epidemiology · RNA and protein synthesis mechanisms · Antibiotic Resistance in Bacteria
INTRODUCTION
The genetics of the model bacterium Escherichia coli is made up of hierarchical regulatory systems functioning on both local and global scales (1, 2). On a local scale, transcription factors determine whether conditions for transcriptional initiation are met, while global systems control entire genome subsets for example via sigma factors directing RNA polymerase (RNAP) to specific promoter motifs. Some transcription factors function on a global scale, such as the cyclic AMP receptor protein, Crp, which regulates the expression of hundreds of genes and acts like a nucleoid-associated protein (1, 3–5). In the absence of glucose, adenylate cyclase (Cya) produces cAMP, and Crp-cAMP then activates genetic programs that enable the utilization of alternative carbon sources (6). In addition, global regulation by Crp responds to deficiencies in nitrogen metabolism and synthesis of nucleic acids, making Crp a ubiquitous regulator for stress adaptation (7, 8).
RpoS (σ^38^) is a main sigma factor in the stationary phase, playing a role when cells have exhausted resources where growth stalls and bacterial aging starts due to oxidative stress (9–11). Both Crp and RpoS are associated with starvation but increased activity of Crp causes a decrease in intracellular RpoS, and correspondingly, strains with no Crp activity show increased resistance to oxidative stress due to increased levels of RpoS (11, 12). Another sigma factor associated with stress is RpoH (σ^32^), classically termed a heat shock sigma factor, although knockouts are severely impaired already above room temperature (13, 14) and a broader physiological role in stationary phase has been indicated (15). Several heat shock proteins are also important during starvation stress, and, thus, the heat shock implication and quasi-essentiality of RpoH may be due to its more general role in protein folding, for example in growth phase transitions where the cellular machinery goes through extensive reorganization (15, 16).
Sigma factors of the RpoD family, which includes RpoS and RpoH, recognize specific promoter motifs termed the −35 and −10 boxes based on their positions in relation to the transcription start site (TSS, +1) (17). In the case of RpoD, the “house-keeping” sigma factor, the −35 box consensus sequence is 5′-T_35_T_34_G_33_A_32_C_31_A_30_-3′, and the −10 box is 5′-T_12_A_11_T_10_A_9_A_8_T_7_-3′. T_35_ and T_34_ are the most conserved, and A_11_ and T_7_ are the most important for open complex formation (17–19). Some promoters contain additional recognition sequences downstream (discriminator region, DR) and upstream (extended −10 boxes, ext-10) of the −10 box, depending on specific domains in the sigma factor (20–22). For example, the region 1.2 domain recognizes the DR.
The MalT transcriptional activator controls the synthesis of enzymes required for the utilization of maltose and maltodextrins (23). Crp binds the PmalT promoter at position −70.5 and is an example of a class I Crp promoter, where Crp binds upstream (centered at position -61.5,–71.5, −81.5, or −91.5) from the RNAP αCTD domain. This is opposed to class II Crp promoters, where Crp binding is centered at position −41.5 (24–26). The 10 bp-spacing regularity is due to the spatial alignment of Crp and the αCTD for every turn of the DNA helix (24, 27).
In previous studies, using a cAMP-deficient E. coli strain, regions upstream from malT and sdaC were found to be mutational hotspots (4, 28). Using fluorescence-based reporters of promoter activity, we use these evolved cis-acting mutations as evolutionary clues to investigate the malT and sdaC promoter architectures in relation to different sigma factors and Crp and how gene expression adapts during long-term stress.
RESULTS
Mutations develop upstream from malT and sdaC in starving bacterial colonies
Due to the absence of cAMP, the cya mutant of Escherichia coli K-12 MG1655 only forms small white colonies on maltose MacConkey agar but upon prolonged incubation, maltose fermenting (red) and non-fermenting (white) secondary colonies appear (Fig. 1a) (4, 28). Previously, in this evolutionary model system, mutations were identified upstream from malT (8, 28) and sdaC (4): a PmalT** mutation was isolated as a red secondary colony at 37°C, and two different PsdaC** mutants were isolated as white secondary colonies both at 37°C and 44°C (Fig. 1b and c). Looking at the DNA sequences, it appears that the three mutations result in putative −10 box sequences closer to matching the RpoD consensus (Fig. 1c). An identical P*malT** mutation was isolated in another study and was shown to increase transcription of malT in the absence of Crp (29). This suggests that transcription initiation at these promoters is suboptimal under the conditions where the mutations developed, but does it reveal more about the specific promoter architectures?
malt and sdaC mutations occurring in experimental evolution of Escherichia coli. (a) Illustration of the experiment. E. coli K-12 MG1655 cya was plated on MacConkey maltose agar, small white colonies formed, and over time maltose fermenting (red) and non-fermenting (white) secondary colonies appeared. (b) Mutational hotspots found previously (4, 8, 28) mapped by genomic locations, and marked by mutant phenotype and mutation type; red, fermenting; white, non-fermenting; (c), cis-acting mutation; (T), trans-acting mutation. (c) Consensus sequence for RpoD, along with alignment of the cis-acting malT (SNP) and sdaC (insertions) promoter mutations () to their native (wt) sequences in E. coli K-12 MG1655. (d) Illustration of the sdaCB operon and its function. L-serine import is mediated by the sdaC-encoded L-serine-H+-symporter, and degradation of L-serine to pyruvate and NH4+ is catalyzed by the L-serine deaminase SdaII. (e) Serial dilutions of E. coli K-12 MG1655 cya with the native or mutant PsdaC promoters on M9 minimal media at 37°C in the presence or absence of L-serine as the sole carbon source. Two independent isolates of each mutation type were found and are marked (1) and (2).*
Mutations upstream from sdaC enable growth on L-serine
The sdaC promoter was previously shown to be regulated by Crp (30). The sdaC gene encodes an L-serine importer that enables the use of extracellular L-serine as an energy source (Fig. 1d), and we speculated that the mutations in PsdaC** could improve the utilization of this amino acid. This was confirmed both at 37°C and 44°C when observing the growth of the four independently isolated PsdaC** strains on a minimal medium with L-serine as the sole carbon source (Fig. 1e; Fig. S1).
A fluorescent reporter predictably and sensitively reports on key regulatory features of PmalT
In the malT promoter, only 3/6 and 2/6 nucleotides match the RpoD −35 box and −10 box consensus sequences, respectively (Fig. 1c) (29). The A_11_ and T_7_ nucleotides important for open complex formation are conserved, and the PmalT** mutation adds the consensus T_12_, thereby potentially improving RpoD-mediated transcription, which could explain the selective advantage in a cAMP-deficient environment (29, 31). On the other hand, RpoD should play only a minor role under the long-term starvation conditions where we isolated PmalT**, motivating further studies of the molecular mechanisms at play.
To further investigate the regulation of malT, we took advantage of a reporter plasmid, which harbors the superfolder gfp gene under the control of PmalT (Fig. 2a) (8). The reporter is tightly repressed by glucose in E. coli K-12 MG1655 (Fig. 2b), highly inducible and titratable by cAMP in a cya knockout strain (Fig. 2c) (8), and sensitive to a cAMP-independent Crp* mutation (8) (Fig. 2d). It therefore shows promise for providing insights into the dynamics of PmalT regulation.
Characterization of the malT promoter and hybrid RpoD consensus mutants. (a) Illustration of the reporter plasmid according to SBOL standards, elements being; a truncated malT promoter, without the Mlc site, an RNA-stability element, a ribosomal binding site, the coding sequence of superfolder GFP, an ssrA tag for protein degradation, and a terminator. The reporter also contains an ampicillin resistance marker for plasmid maintenance and a kanamycin resistance marker with FRT sites for genomic integration. (b) Fluorescence from the PmalT reporter in E. coli K-12 MG1655 after 9 h of growth in the presence or absence (Ø) of glucose (glc, 1%). Data represent biological duplicates. (c)The activity of the reporter in E. coli K-12 MG1655 cya after 9 h of growth with different levels of cAMP. Data represent biological duplicates. (d) Fluorescence from the PmalT reporter in E. coli K-12 MG1655, cya, and cya crp through 12 h of growth. Data represent biological triplicates. (e) Promoter layout of (top to bottom) RpoD consensus, the native PmalT sequence from E. coli K-12 MG1655, and hybrid promoters mutants where the −10 box, −35 box, or both are changed to the RpoD consensus (red font). (f) Fluorescence from the native and consensus PmalT reporter variants in E. coli K-12 MG1655 wt and cya after 12 h of growth, with no supplement (Ø, grey), 1% glucose (wt only, blue), or 0.5 mM cAMP (cya only, red). Data represent two or three biological replicates.*
We first entertained a synthetic biology approach to better understand the regulation of PmalT. The poor similarity to the −10 box RpoD consensus sequence has been speculated to be the main cause of Crp-dependence (31), but the −35 box of PmalT also differs from the RpoD consensus. This prompted us to systematically investigate changes in PmalT toward the RpoD consensus. When converting either the −10 or −35 box to the exact RpoD consensus (Fig. 2e), fluorescence from the PmalT reporter increased as expected (Fig. 2f). However, while the consensus −35 mutants retained almost full glucose repression and fourfold cAMP induction, the −10 mutants only showed 20%–30% glucose repression and twofold stimulation by cAMP (Fig. 2f). This confirms that the low similarity of the −10 box to the RpoD consensus is the dominating cause of Crp-dependence (29). Nevertheless, even for the exact RpoD consensus mutants (“-10/–35”) in the wild-type background, fluorescence was still stimulated by cAMP (+50%, Fig. 2f).
An alternative promoter may enable Crp-independent malT transcription in the stationary phase
Looking at the malT promoter region, a close match for another −10 box (T_7_A_6_T_5_A_4_A_3_C_2_) is present five bp downstream from the previously annotated PmalT −10 box (Fig. 3a). We decided to investigate whether transcription from this alternative −10 box could be stimulated by re-aligning the Crp binding site position. To this end, we constructed mutants of the malT promoter region with the Crp binding site re-positioned from the natural −70.5 position to −75.5 (±1),–65.5 (±1),–60.5, −55.5, and −50.5, and near class II positions 43.5 to −38.5 (Fig. S2). Fluorescence was monitored throughout growth and is in Fig. 3b plotted after 12 h of growth when the strain had entered the stationary phase. However, the observed trends were consistent throughout all growth phases: In accordance with previous studies, local optima in promoter activity at sites corresponding to class I (−60.5) and class II (−42.5) Crp-binding sites were observed (Fig. 3b) (24, 25). The lack of a local optimum at position −50.5 is expected due to the binding of the RNAP to the promoter at this position (32). Fluorescence as a result of a potential secondary transcript was significantly less intense than from the main transcript; we observed low signals from mutants −65.5 and −55.5, and thus, if a secondary promoter, as shown in Fig. 3a, is indeed active in vivo, it is not stimulated by Crp binding at class I binding sites (Fig. 3b).
Effects of varying the Crp binding site location within the malT promoter. (a) RpoD consensus sequences and the malT promoter with annotated and alternative RpoD recognition sequences and TSS—see main text for further description. The identity between the recognition sites and the consensus sequences is indicated by the numbers above. (b) Fluorescence from the PmalT reporter variants after 12 h of growth. Indicated are the native distances between the Crp-binding site and +1, and the distances classically associated with Crp promoter classes I and II. Data represent two or three biological replicates. (c) Fluorescence from the native PmalT reporter (−70.5) compared to the −60.5 variant in E. coli K-12 MG1655 WT or a cya strain supplemented with 0.5 mM cAMP after 12 h of growth. Significance was based on two-sided unpaired t-tests between the groups indicated and designated as significant if P < 0.05. Data represent biological duplicates.
The dynamic range of expression is higher for a non-native PmalT variant
An interesting observation, upon varying the distance between the annotated transcription start site and the Crp binding site, is that the fluorescence for PmalT with Crp in position −60.5 is close to, or even higher than that of the native promoter (Fig. 3b), as previously observed (24). When looking at other RpoD promoters, activating Crp sites are more often annotated around position −61.5 than at −71.5 (Fig. S3) (33); however, the evolutionary background for the selection of one class I site position over others remains unelucidated. In the data shown in Fig. 3c, the fluorescence for position −60.5 relative to position −70.5 was even higher when a ΔcyaA strain was supplemented with cAMP. Thus, in this assay, position −60.5 provides a larger dynamic range of expression than position −70.5. Nevertheless, the −70.5 position is preserved in K-12 MG1655. What might provide a selective advantage of binding Crp in this position? Do additional factors, besides those already known (29, 33–35), play a role?
Engineered RpoD consensus mutants show growth-phase-dependent effects
Given that different sigma factors dominate in different growth phases, we decided to look closer at the growth and time-resolved fluorescence of the RpoD consensus promoter mutants. As one would expect, all three PmalT RpoD consensus mutants exhibited increased fluorescence compared to the wild-type PmalT in the early stages of growth (Fig. 4a, top). However, the two −10 box mutants exhibited an approximately 2 h extended lag phase indicating early expression stress (Fig. 4a). In the transition from exponential to stationary phase, none of the constructs exhibited an increase in fluorescence, showing either a temporary stall (−10 box mutants) or a local minimum. This “pause” before the stationary phase could be due to changes in the transcriptional machinery involving sigma factors that take place around this point in growth (36, 37). This, however, poses a new question: If RpoD is exchanged from the transcriptional machinery upon entry into the stationary phase, exactly what sigma factor is responsible for the continued expression from PmalT throughout the stationary phase?
Expression from malT promoter reporters upon varying the central promoter motifs. (a) Fluorescence (top) and growth (bottom) of E. coli K-12 MG1655 with the native or consensus PmalT reporter variants during 12 h. Promoter variants: Solid line, native; dotted line, −10 consensus; stippled line, −35 consensus; dotted and stippled line, −10 and 35 consensus. Black, lag and log phase; red, stationary phase. (b) Alignment of PmalT to the reported consensus sequences of RpoD, RpoS, and RpoH, with potential recognition sites. RpoS and RpoH both recognize extended −10 boxes, although RpoH often is designated part of the −10 box (38). (c) Fluorescence with standard deviations from the PmalT reporter in E. coli K-12 MG1655 wt and rpoS during 48 h of growth. Solid line, wt; stippled line, rpoS. (d) Fluorescence from the native and −60.5 variant of PmalT in E. coli K-12 MG1655 cya crp pSEVA-crp with various rpoS genotypes after 12 h of growth. Gray, rpoS+; red, rpoS-; blue, rpoS N98K. (e) Fluorescence with standard deviations (top) and growth (bottom) of E. coli K-12 MG1655 with the PmalT reporter during 18 h. Solid line, native promoter; stippled line, evolved promoter; black lines, wild-type strain; gray lines, cya. Data represent biological triplicates.*
The malT promoter shows similarities to alternative sigma factor recognition sites
Both RpoS and RpoH are associated with stress responses and stationary phase (15, 37, 38) and could be involved in malT expression. The DNA sequence of PmalT is a potential good match for RpoS when taking into account the nucleotides in the RpoS ext-10 box (T_17_ and C_13_), and the discriminator region, which is A-T rich, as reported optimal for RpoS (Fig. 4b) (39). However, motifs can be found that match the RpoH consensus as well (Fig. 4b) (8, 40, 41). It is also worth noting that the PmalT mutants with an RpoD consensus −10 box retain similarity to the RpoS but not the RpoH consensus sequence. Thus, one hypothesis for the observed Crp-independence of these mutants could be that a RpoD-consensus −10 box causes a shift away from RpoH-driven transcription toward RpoS-driven transcription.
The malT promoter is affected by different rpoS mutants
Potential effects of RpoS on PmalT were explored by monitoring the activity of the native malT promoter in mutant rpoS strains. Compared to the rpoS+ control, an rpoS knockout strain shows a significant decrease in fluorescence in the early stationary phase (Fig. 4c). This indicates the involvement of RpoS in the expression of malT.
Crp-independent effects were assayed in the cya mutant background where low levels of fluorescence were observed after 12 h, both in the presence and absence of rpoS. An RpoS_N98K mutant evolved on several independent occasions during long-term starvation in a previous study (28) and this mutant showed increased fluorescent levels from the malT promoter (Fig. 4d) but not the synthetic −60.5 PmalT variant. This provides independent evidence for RpoS-driven transcription and suggests that it is dependent on upstream elements that are compromised in the −60.5 mutant (Fig. 4d). Unfortunately, experiments to test potential in vivo direct interaction between RpoH and PmalT were inconclusive due to the severe physiological changes caused by rpoH deletion.
The PmalT* mutation enables malT expression in the absence of cAMP
The potential interplay between a stationary phase sigma factor and PmalT is not in conflict with the naturally evolved cis-acting mutation in PmalT (Fig. 1c), as the similarity to the consensus sequences is preserved for RpoH and improved for RpoS. To further explore the consequence of the PmalT** mutation, we introduced the point mutation into the gfp reporter construct. Similar to the −10 box mutants, when expressing gfp from PmalT** in a wild-type strain, growth exhibits a 3 h extended lag phase, indicating expression stress (Fig. 4e). However, the clearest effect was seen in the difference in fluorescence between PmalT and PmalT** in a cya strain, where fluorescence was completely absent from PmalT* but high with PmalT** (Fig. 4e). This clearly explains the selective advantage of the PmalT** mutation for growth on maltose in a cya strain.
sdaC is mainly expressed in stationary-phase and is increased in sdaC* mutants
Returning to sdaC, as interactions of alternative sigma factors with PmalT are indicated in the data presented here, it is not unreasonable to suspect that the cis-acting mutations that evolved in PsdaC under similar conditions similarly relate to sigma factor usage. Except for binding sites for Crp and Lrp (30, 42), only a few sdaC promoter features have been described. A TSS for an RpoD promoter has been annotated, and an RpoH-driven promoter has previously been implicated upstream from sdaC but no promoter has been experimentally verified (43, 44). Similar to malT, we monitored the growth and time-resolved fluorescence from PsdaC-GFP reporters with or without the PsdaC** mutations (Fig. 1c). The mutant promoters showed increased fluorescence from early stationary phase, with an additional increase occurring after additional ~3–4 h in stationary phase (Fig. 5a). At 44°C, the temperature where some of the PsdaC** mutants were isolated, the PsdaC** mutants show consistently higher levels of fluorescence compared to the native promoter in both exponential and stationary phase (Fig. S1). To test the involvement of RpoS in the regulation of PsdaC*, we assayed the wild-type and mutant promoters in an rpoS background. The two mutant promoters, especially PsdaC**2*, showed decreased fluorescence in the rpoS knockout background, while the wild-type promoter was unaffected (Fig. 5b). This implicates the involvement of RpoS in the regulation of the mutant sdaC promoters.
Growth-phase dependent regulation of PsdaC. (a) Fluorescence with standard deviation (top) during 20 h of growth (bottom) of E. coli K-12 MG1655 with the PsdaC reporter mutants. Promoter variants: Solid line, native (wt); stippled line, mutant 1; dotted line, mutant 2. (b) Fluorescence with standard deviations from the native PsdaC (left), PsdaC1 (middle), and PsdaC2 (right) in E. coli K-12 MG1655 wt (grey) and rpoS (red) through 20 h of growth. Solid line, native (wt); stippled line, mutant 1; dotted line, mutant 2.
DISCUSSION
Promoter-activating mutations are frequent outcomes of evolution (45) and have led to important insights into the mechanisms of regulation (46). The cis-acting mutations characterized here explain well the growth benefits gained under the specific selective conditions: As PmalT* was isolated at 37°C on maltose, the main selection regime was carbon starvation, which would be abolished, should the cell gain the ability to utilize maltose; a likely outcome of increased malT expression. Similarly, PsdaC controls the sdaCB operon, and overexpression may result in an increased rate of L-serine import into the cell and the use of L-serine in an energy-providing role (47). Selection of P*sdaC** at 37°C and 44°C thus implies that L-serine can be used as a carbon source from MacConkey agar during both starvation and heat stress. Alternatively, there is strong selection pressure to express sdaC to prevent cell lysis during starvation (48).
In both cases mentioned above, it can be reasoned that the relief of stress is achieved via adapting the promoter in question to the transcriptional machinery active at the time of the mutational event. The mutations were isolated following days or weeks of starvation stress (and for some PsdaC mutations, additional heat stress), causing growth to stall. Since RpoS is stabilized during conditions such as long-term starvation and heat stress (11, 16, 36), it seems likely that the mutants were rescued from stress based on cis-acting mutations improving RpoS interactions.
The sdaC promoter, as annotated in this study, shows a great match to the RpoD consensus, especially when considering the high GC content in the discriminator region and the lack of an extended −10 sequence. RpoD and RpoS consensus sequences show significant overlap, and the difference in the transcription of some RpoD/RpoS promoters may simply be growth phase dependent, as RpoS shows a higher affinity for relaxed DNA in stationary phase (49). Insertions of +ATT (1) or +A(2) in the −10 box of PsdaC elongates the AT-rich sequence of this promoter, resulting in a discriminator region with diminished GC content, which may improve RpoS recognition of the promoter. Indeed, PsdaC1/2* show increased promoter activity through both exponential and stationary phase, and a knockout of rpoS diminishes this effect somewhat, indicating dual transcription by RpoD and the stationary phase sigma factor. Similarly, the mutation in P*malT, while changing the consensus of the −10 box towards the RpoD consensus, simultaneously improves the similarity to the RpoS consensus. When considering the already AT-rich discriminator region, and a match for a potential extended −10 region, the native malT promoter, while annotated as RpoD-transcribed, may very well be transcribed by RpoS as well; a hypothesis supported by the in vivo data presented here. The native malT promoter also contains motifs that may indicate RpoH recognition, and the PmalT** mutations were previously isolated during long-term starvation (29, 31). Thus, it seems that there is a connection between PmalT** and the expression context that is long-term stationary phase, suggesting that P*malT** gains the ability to be transcribed by RpoS or RpoH, while native RpoD-dependent and possibly RpoH-dependent transcriptional programs are preserved.
As RpoS partially controls the transcription of rpoH in the stationary phase, the RpoS-dependent increased transcription from PmalT** and PsdaC**1/2 could implicate either sigma factor. The high frequency of rpoS knockout mutations isolated during long-term starvation at both 37°C and 44°C (4, 28), and a mutation found in ftsH (4), a protease that degrades RpoH, could suggest that although RpoS may recognize the promoters, expression using RpoH is more important for long-term survival. The interplay between RpoS and RpoH should also be considered in the context of the frequently observed rpoS knockout mutations that are associated with a growth advantage in the stationary phase (50–53).
Besides its status as a heat shock sigma factor, RpoH is implicated in general stress responses as a key regulator of chaperones and proteases, ensuring homeostasis of protein function in E. coli. RpoH is also, directly or indirectly, implicated in starvation responses (15, 54), DNA relaxation (independently from heat-shock-induced DNA relaxation) (55), and oxidative stress resistance in the exponential phase (56). RpoH has previously been suspected to be involved in the regulation of the malT promoter; in previous work, the supplementation of cytidine to a strain carrying the PmalT reporter caused a reduction in fluorescence output while PmalT** abolished this effect (8). Cytidine binds to CytR, which represses rpoH transcription, releasing repression and potentially resulting in increased RpoH levels. PmalT* activity could thus be diminished in the presence of cytidine due to decreased availability of RNAP-RpoD relative to RNAP-RpoH; however, the lack of a cytidine response for PmalT** rather suggests that the repressive effect of cytidine on PmalT* is mediated by the sigma factor recognition of PmalT, most likely via RpoH. Due to the difficulties in assaying RpoH effects on promoter activity without perturbing the entire genetic system by low growth temperatures, RpoH interactions with the malT promoter need further investigation.
Considering how the consensus sequences for RpoD and RpoH partially overlap and can coexist in one promoter, it is possible that during certain environmental conditions or metabolic states, RpoH can transcribe promoters otherwise designated RpoD, for example, a hypothetical scenario during carbon starvation when RpoD is sequestered and RpoS levels are decreased due to Crp activity. RpoH is known to undergo structural changes upon induction of the heat shock response, so promoter recognition may even change depending on the temperature. One can imagine how functional overlap of the transcription machinery may be beneficial for the fitness and adaptability of evolved microbes; it allows the cell to rapidly respond to changes in environmental conditions and survive until the transcription machinery is exchanged in favor of long-term stress responses. The observations reported here indicate that PmalT and P*sdaC** are recognizable by RpoS, and potentially RpoH, although RpoD-dependent transcription may be favored in the presence of Crp-cAMP, and hints that this interplay is affected by the location of the Crp-binding site in a class I Crp promoter. Thus, the regulation of the malT operon, and other metabolic genes, is based on an intricate interplay between Crp and sigma factors depending on the metabolic state.
MATERIALS AND METHODS
Bacterial strains
All strains applied were E. coli K-12 MG1655. The cya deletion (cya::cat) was introduced into a Δfnr background by λRed recombineering in previous work (28). Experimental evolution of cis-acting mutations, the crp* (crpA144T), as well as rpoSN98K, in the cya background were described previously (4, 28). The rpoS gene was deleted in wild-type and cya backgrounds using λRed recombineering with pSIM19 as previously described (57, 58) and integration of a tetA PCR product with homology ends to the rpoS locus as previously described (59). To remove and avoid secondary mutations in crp, the crp gene was further deleted in cya rpoSN98K, cya rpoS, and cya, and supplied on a low-copy plasmid described previously (8).
Growth conditions
Strains were grown in LB broth or on LB agar at 37°C unless otherwise mentioned and supplemented with ampicillin (100 µg/mL) or kanamycin (50 µg/mL) for plasmid maintenance when necessary. For characterization of serine utilization in sdaC* mutants, strains were drop-tested on M9 minimal medium supplemented with 2% L-serine. Supplements used for growth experiments were additionally cAMP (0.5 mM) or glucose (1%) as denoted.
Plasmid construction
Plasmids were constructed by USER cloning of relevant fragments (60) and sequenced before application. All plasmids are shown in Table S1 and oligonucleotides used for plasmid constructions as listed below are shown in Table S2. The PmalT reporter variants were constructed as one-fragment clonings based on the native PmalT reporter as follows: Variants −76.5 to −64.5 were constructed using oligo #5733 in combination with oligos #5727-#5732, variants −60.5 to −38.5 were constructed using oligo #5590 in combination with oligos #5591-#5600, and variants -10,–35, and -10/–35 were constructed using oligo #5726 in combination with oligos #5734-#5736. The PsdaC reporters were constructed based on the native PmalT reporter as a vector backbone, using oligos #4256 and #4258, and the genomic sdaC promoter amplified from the native or mutant strains using oligos #4577 and #4578.
Growth experiments
Fresh colonies from cryostock were inoculated into a 5 mL preculture and grown over the day (7 h). From the preculture, 1 µL was used for inoculation into 150 µL medium in a 96-well microtiter plate (Costar 96 flat bottom), which was sealed using a breath-seal film and incubated in a Synergy H1 microplate reader (BioTek) (20–48 h, 37*°C or 44°*C, orbital shaking continuously at 425 cpm, reading depth of 3 mm). Optical density at 600 nm and fluorescence (excitation 485 nm, emission 528 nm, gain 50–100) was measured every 10–15 minutes. Data output in Microsoft Excel were subjected to preprocessing and visualized and analyzed using GraphPad Prism 9.1.0.
Prediction of Crp binding sites
The PredCRP program was downloaded from github and executed in the Unix terminal using Python. The intergenic region between ppnN and sdaC was used as input (reference sequence NC_000913.3). The sequence was evaluated using a 42 bp moving window and plotted using the position of the base in the center right of the moving window. The output of PredCRP was exported as .csv and is included in Microsoft Excel format as Supplementary Data 2.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Martínez-Antonio A, Collado-Vides J. 2003. Identifying global regulators in transcriptional regulatory networks in bacteria. Curr Opin Microbiol 6:482–489. doi:10.1016/j.mib.2003.09.00214572541 · doi ↗ · pubmed ↗
- 2Browning DF, Busby SJW. 2016. Local and global regulation of transcription initiation in bacteria. Nat Rev Microbiol 14:638–650. doi:10.1038/nrmicro.2016.10327498839 · doi ↗ · pubmed ↗
- 3Zheng D, Constantinidou C, Hobman JL, Minchin SD. 2004. Identification of the CRP regulon using in vitro and in vivo transcriptional profiling. Nucleic Acids Res 32:5874–5893. doi:10.1093/nar/gkh 90815520470 PMC 528793 · doi ↗ · pubmed ↗
- 4Heyde SAH, Frendorf PO, Lauritsen I, Nørholm MHH. 2021. Restoring global gene regulation through experimental evolution uncovers a NAP-like behaviour of Crp/Cap. m Bio 12:e 0202821. doi:10.1128/m Bio.02028-2134700380 PMC 8546631 · doi ↗ · pubmed ↗
- 5Grainger DC, Hurd D, Harrison M, Holdstock J, Busby SJW. 2005. Studies of the distribution of Escherichia coli c AMP-receptor protein and RNA polymerase along the E. Proc Natl Acad Sci USA 102:17693–17698. doi:10.1073/pnas.050668710216301522 PMC 1308901 · doi ↗ · pubmed ↗
- 6Zubay G, Schwartz D, Beckwith J. 1970. Mechanism of activation of catabolite-sensitive genes: a positive control system. Proc Natl Acad Sci USA 66:104–110. doi:10.1073/pnas.66.1.1044320461 PMC 286094 · doi ↗ · pubmed ↗
- 7Daniel J, Danchin A. 1986. 2-Ketoglutarate as a possible regulatory metabolite involved in cyclic AMP-dependent catabolite repression in Escherichia coli K 12. Biochimie 68:303–310. doi:10.1016/s 0300-9084(86)80027-x 3015255 · doi ↗ · pubmed ↗
- 8Lauritsen I, Frendorf PO, Capucci S, Heyde SAH, Blomquist SD, Wendel S, Fischer EC, Sekowska A, Danchin A, Nørholm MHH. 2021. Temporal evolution of master regulator Crp identifies pyrimidines as catabolite modulator factors. Nat Commun 12:5880. doi:10.1038/s 41467-021-26098-x 34620864 PMC 8497467 · doi ↗ · pubmed ↗