Chloroplast genome data of five Amygdalus species: Clarifying genome structure and phylogenetic relationships
Yixiao Chen, Wenquan Bao, Dun Ao, Yue Bai, Haiguang Huang, Rong Yang, Lin Wang, Ta-na Wuyun

TL;DR
This study sequenced chloroplast genomes of five Amygdalus species to better understand their genetic relationships and structure.
Contribution
The paper provides new chloroplast genome data and clarifies phylogenetic relationships among five Amygdalus species.
Findings
The chloroplast genomes of five Amygdalus species were sequenced and show high structural similarity.
Four highly divergent genomic regions were identified, offering insights into genetic variation.
Phylogenetic analysis grouped the species into distinct branches, aiding in understanding their evolutionary relationships.
Abstract
Amygdalus species have considerable ecological and economic value, however, the phylogenetic relationships among Amygdalus remain controversy. In this study, we sequenced and assembled the chloroplast (cp) genomes of five Amygdalus species: Prunus communis, P. mongolica, P. pedunculata, P. triloba, and P. mira. We then conducted comparative genomic analyses and constructed their phylogenetic relationships. The genome length ranged from 157,870 to 158,451 bp, and 131 genes were annotated (86 protein-coding genes, 37 tRNAs, and 8 rRNAs). Additionally, 49–57 simple sequence repeats were detected, with most in the large single-copy region and with AT base preferences. Comparative genomic analyses revealed high similarities in structure, order, and gene content. However, we identified four highly divergent sequences: trnR-UCU-atpA, nbdhC-trnV-UAC, ycf4-cemA, and rpl32-trnL-UAG. The…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
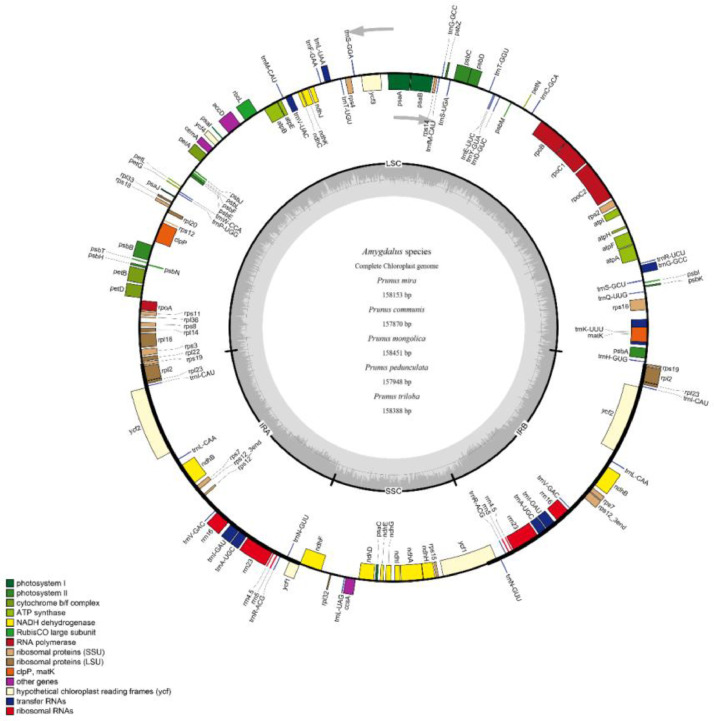 Figure 1
Figure 1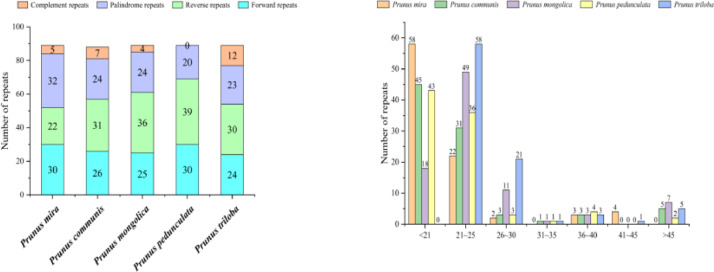 Figure 2
Figure 2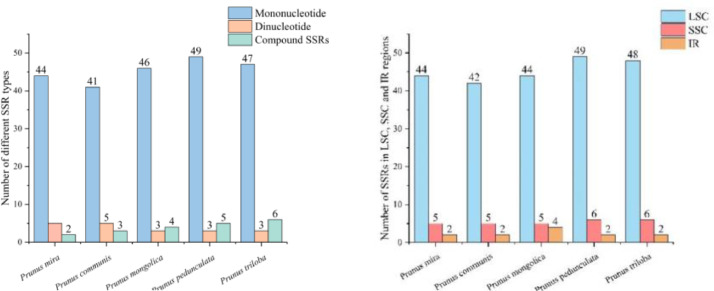 Figure 3
Figure 3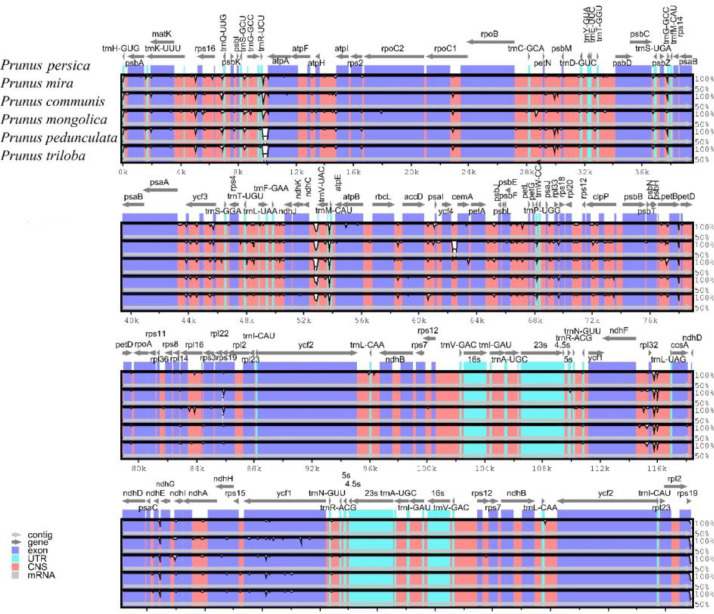 Figure 4
Figure 4 Figure 5
Figure 5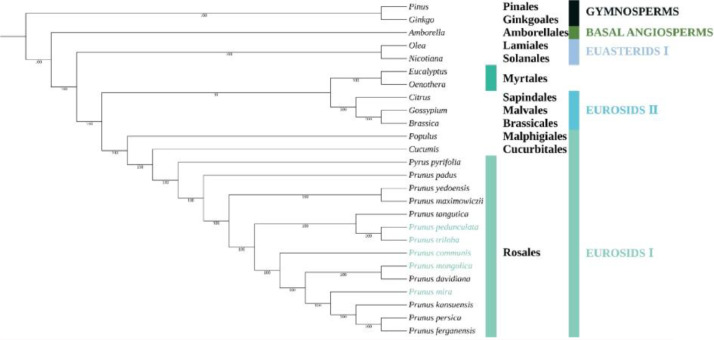 Figure 6
Figure 6Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGenomics and Phylogenetic Studies · Genetic diversity and population structure · Plant Diversity and Evolution
Specifications TableSubjectBiological sciencesSpecific subject areaOmics: GenomicsData formatRawAnalyzedType of dataTableFigureData collectionAn Illumina Hiseq X high-throughput platform (Illumina, San Diego, CA, USA) was used for DNA sequencing. Chloroplast genome fragments were assembled using SOAPdenovo (http://soap.genomics.org.cn/soapdenovo.html) to obtain contigs and optimized assembly results according to the reference cp genome of Prunus persica (GenBank accession number NC_014697.1). Genomes were annotated using DOGMA (http://dogma.ccbb.utexas.edu/). An annotated cp genome map was constructed with Organellar Genome Draw (https://chlorobox.mpimp-golm.mpg.de/OGDraw.htmL).Data source location
- •Institution: Non-timber Forest Research and Development Center, Chinese Academy of Forestry
- •City: Zheng zhou
- •Country: China Data accessibilityRepository name: Mendeley DataData identification number: DOI:10.17632/kh83zb2f9p.1Direct URL to data: https://data.mendeley.com/datasets/kh83zb2f9p/1
Value of the Data
1
- •Chloroplast genome sequences of five Amygdalus species provides an improved understanding of the genetic relationships among the Amygdalus.
- •Identification of these SSR loci and variations provides candidate molecular markers for research on population diversity and evolutionary research.
- •The chloroplast genome data provides a basis for the development and utilization of Amygdalus resources.
Data Description
2
The genus Amygdalus was classified into subgenus Amygdalus and subgenus Persica [1]. The subgenus Amygdalus is mainly distributed in the Mediterranean region and central-eastern Asia, with the exception of P. triloba, which widely distribution in northwest China [2]. The P. triloba, P. pedunculata, P. mongolica, and P. communis belonged to subgenus Amygdalus, and the kernels were rich in oil and protein, which can be use as high-quality oil and protein resource [3,4]. The shell can also be used as fuel and adsorbent for heavy metals and pigments [5]. P. mira, belonging to subgenus Persica, is mainly distributed in the Yarlung Zangbo Grand Canyon and the tributary basins of Tibetan Plateau [6]. The kernels of P. mira are rich in oleic acids, linoleic acids, and fat-soluble components, which was used for Chinese traditional medicine to treatment and improvement of diseases [7].
The classification of genus Amygdalus has always been controversial [8]. Base on the morphological classification, the P. communis, P. mongolica, P. tangutica, P. triloba, and P. pedunculata were classified into subgenus Amygdalus, whereas, the P. mira, P. davidiana, P. ferganensis, P. kansuensis, and P. persica were classified into subgenus Persica [1]. However, Wang et al. [9] revealed that Prunus tenella, P. pedunculata, and P. triloba should be classified into the genus Prunus L., while, P. communis, P. mongolica, and P. tangutica be divided into the subgenus Amygdalus L. by using cp genomes, and Yazbek et al. [10] investigated the phylogeny of Prunus subg. Amygdalus by plastid and nuclear genes found that the P. triloba, and P. pedunculata should be excluded from subgenus Amygdalus.
Chloroplasts (cp) are semi-autonomous organelles with a conserved, maternally inherited genome separate from the rest of the plant. The genome size of terrestrial plants cp varies from 100 to 200 kb and generally contains 110–130 genes, which are mainly composed of genes involved in photosynthesis, transcription, and translation [11]. Most higher-plant cp genomes form a quadripartite structure, containing one pair of inverted repeats (IR), a small single-copy (SSC) region, and a large single-copy (LSC) region separated by IRs [12]. Therefore, the study of the cp genome plays an important role in species identification, phylogenetic analysis, and molecular marker development [13,14]. The rapid development of next-generation sequencing technologies and phylogenetic genomics has led to the sequencing of cp genomes from multiple Prunoideae species, which are widely used in molecular evolution and phylogenetic research [15], [16], [17]. However, the cp genomes of Amygdalus species were remain insufficient. Here, we sequenced, assembled, and performed phylogenetic analysis on the complete cp genomes of five Amygdalus species (P. communis, P. mongolica, P. pedunculata, P. triloba, and P. mira) aimed to provide an improved understanding of the genetic relationships among the Amygdalus and provide a basis for the development and utilization of Amygdalus resources.
Total cp genome lengths for P. mira, P. communis, P. mongolica, P. pedunculata, and P. triloba were 158,153, 157,870, 158,451, 157,948, and 158,388 bp, respectively (Fig. 1). Each cp genome exhibited a typical quadripartite structure, including two IRs (26,373–26,931 bp), one LSC (86,144–86,525 bp), and one SSC (18,966–19,211 bp). The GC content in the cp genome ranged from 36.72 % in P. mongolica to 36.78 % in P. pedunculata and was higher in the IR regions (ranging from 42.55 % in P. mira to 42.60 % in P. communis) than that in the LSC regions (ranging from 34.57 % in P. communis to 34.62 % in P. pedunculata) and the SSC regions(ranging from 30.27 % in P. mongolica to 30.46 % in P. communis), suggesting that the two IR regions were relatively stable.Fig. 1. Assembly, size, and features of the chloroplast genomes from five Amygdalus species. The dark gray area in the inner circle represents genomic GC content, whereas the light gray area indicates AT content.Fig 1
We annotated 131 genes in each of the five Amygdalus cp genomes, including 86 protein-coding genes, 37 transfer RNAs (tRNA), and eight (rRNAs) (Table 1). These genes were in the same order across all five cp genomes.Table 1. Chloroplast genomes of the five Amygdalus species.Table 1. SpeciesGenomeG-C Content(%)GenesCDStRNArRNAGenomeLSCSSCIRPrunus mira158,15336.7434.630.3542.5513186378Prunus communis157,87036.7634.5730.4642.613186378Prunus mongolica158,45136.7234.5830.2742.5813186378Prunus pedunculata157,94836.7834.6230.4542.5913186378Prunus triloba158,38836.7734.6230.4442.5913186378
Functional analysis classified 131 genes into four categories, including photosynthesis-related, self-replication-related, other, and unknown function(Table 2). A total of 18 genes were duplicated in the IR region. Furthermore, 18 contained one intron each, while two genes (clpP and ycf3) contained two introns (Table S1). In addition, the GC content of rRNA (55.5 %) and tRNA (ranging from 53.3 % in P. communis to 53.41 % in P. mira) was higher than that in protein-coding genes (ranging from 37.61 % in P. triloba to 37.65 % in P. communis) (Table S2).Table 2. Genes identified in the chloroplast genomes of the five Amygdalus species.Table 2. Gene categoriesGene groupsGene namesSelf-replicationRibosomal RNAsrrn16^b^, rrn23^b^, rrn4.5^b^, rrn5^b^Transfer RNAstrnA-UGC^a,b^, trnC-GCA, trnD-GUC, trnE-UUC, trnF-GAA, trnfM-CAU, trnG-GCC^a^, trnH-GUG, trnI-CAU^b^, trnI-GAU^a,b^, trnK-UUU^a^, trnL-CAA^b^, trnL-UAA^a^, trnL-UAG, trnM-CAU, trnN-GUU^b^, trnP-UGG, trnQ-UUG, trnR-ACG^b^, trnR-UCU, trnS-GCU, trnS-GGA, trnS-UGA, trnT-GGU, trnT-UGU, trnV-GAC^b^, trnV-UAC^a^, trnW-CCA, trnY-GUAProteins of small ribosomal subunitrps2, rps3, rps4, rps7^b^, rps8, rps11, rps12^a,^^b^, rps14, rps15, rps16^a^, rps18, rps19^b^Proteins of large ribosomal subunitrpl2^a, b^, rpl14, rpl16^a^, rpl20, rpl22, rpl23^b^, rpl32, rpl33, rpl36Subunits of RNA polymeraserpoA, rpoB, rpoC1^a^, rpoC2Genes for photosynthesisSubunits of NADH-dehydrogenasendhA^a^, ndhB^a b^, ndhC, ndhD, ndhE, ndhF, ndhG, ndhH, ndhI, ndhJ, ndhKSubunits of Photosystem ⅠpsaA, psaB, psaC, psaI, psaJSubunits of Photosystem ⅡpsbA, psbB, psbC, psbD, psbE, psbF, psbH, psbI, psbJ, psbK, psbL, psbM, psbN, psbT, psbZLarge subunit of RuBisCOrbcLSubunits of cytochrome b/f complexpetA, petB^a^, petD^a^, petG, petL, petNSubunits of ATP synthaseatpA, atpB, atpE, atpF^a^, atpH, atpIOther genesAcetyl-CoA carboxylaseaccDCytochrome c biogenesisccsAMaturasematKEnvelope membrane proteincemAUnknownConserved hypothetical chloroplast reading framesycf1^b^, ycf2^b^, ycf3^a^, ycf4
We identified 89 repeat sequences, including forward, reverse, palindromic, and complement repeats in P. mira, P. mongolica, P. triloba, and P. pedunculata, while P. communis had 88 repeat sequences (Fig. 2). The lengths of the repeat sequence in P. mira, P. mongolica, P. communis, and P. pedunculata was 18–25 bp, while that in P. triloba was 21–30 bp. Five, seven, four, and twelve complement repeats were found in P. mira, P. communis, Prunus mongolica, and P. triloba, respectively. Reverse repeats (31, 36, 39, 30) were more abundant than palindromic repeats (24, 24, 20, 23) in P. communis, P. mongolica, P. pedunculata, and P. triloba, respectively. In contrast, P. mira had fewer reverse repeats (22) than palindromic repeats (32).Fig. 2(A.left) Number of dispersed repeats in the five Amygdalus species. (B.right) Number of long repeat sequences, clustered by length, in the five Amygdalus species.Fig 2
Three types of SSRs (mononucleotide, dinucleotide, and compound) were present in the five Amygdalus cp genomes (Fig. 3a), in counts ranging from 49 (P. communis) to 57 (P. pedunculata). Mononucleotide repeats (ranging from 83.67 % in P. communis to 86.79 % in P. mongolica) were the most frequently occurring, followed by dinucleotide (ranging from 5.26 % in P. pedunculata to 10.20 % in P. communis) and compound SSRs (ranging from 3.92 % in P. mira to 10.71 % in P. triloba). The number of A (ranging from 29.27 % in P. communis to 40.91 % in P. mira) and T (ranging from 50.00 % in P. mira to 60.98 % in P. communis) mononucleotide repeats was higher than that of C (4.35 % in P. mongolica to 8.51 % in P. triloba) or G repeats (4.08 % in P. pedunculata to 4.88 % in P. communis) (Table S3). and two dinucleotide repeat motifs were obtained: AT (ranging from 3.51 % in P. pedunculata to 5.88 % in P. mira) and TA (ranging from 1.75 % in P. pedunculata to 6.12 % in P. communis).Fig. 3(A.left) Number and type of short sequence repeats (SSRs). B. right) Number of SSRs in the large single-copy (LSC), small single-copy (SSC), and inverted-repeat (IR) regions.Fig 3
We further analyzed the distribution of SSRs (Fig. 3b) and found that most SSRs were distributed in the LSC regions (83.02–86.27 %); fewer were in the SSC regions (9.43–10.71 %) and IR regions (3.51–7.55 %) regions. Additionally, 79.59–84.21 % SSRs were located in intergenic spacer (IGS) regions, while 15.79–20.41 % SSRs were in coding sequences (CDS).
The analysis of cp genome sequence alignment of five Amygdalus species were carried out by using P. persica as a reference, the result showed that the variations in IGS were higher than that in CDS (Fig. 4). We identified four highly divergent sequences in the IGS: trnR-UCU-atpA, nbdhC-trnV-UAC, ycf4-cemA, and rpl32-trnL-UAG. These sequences have potential candidate molecular markers for Amygdalus species.Fig. 4. Sequence alignment of chloroplast genomes from the five Amygdalus species. The y-axis represents percent identity from 50 to 100 %.Fig 4
We compared IR regions among the cp genomes of the five Amygdalus species and two related species (P. persica and P. pyrifolia) (Fig. 5). Although the cp genome structure and gene organization were highly conserved, IR expansions and contractions resulted in slight variations in the LSC/IRb and SSC/IRa borders. The genes of rps19, ycf1, and ndhF were distributed near the boundaries of IR/LSC and IR/SSC. Among them, the ycf1 was detected at the LSC/IRa boundary and the size of ycf1 in the IRa region was 1041–1051 bp, the rps19 was located at the LSC/IRb boundary, with a fragment size of 182–265 bp in the IRb region. The ndhF was located at the IRb/SSC border with a fragment size of 1, 2, 5, and 8 bp in P. triloba, P. pedunculata, P. communis, and Prunus Mira, respectively, however, the ndhF in P. mongolica did not overlap at IRb/SSC boundary.Fig. 5. Comparison of LSC, SSC, and IR border regions across the five Amygdalus species.Fig 5
Our phylogenetic trees based on complete cp genome data had a higher bootstrap value, and 23 out of 24 nodes had 100 % bootstrap values (Fig. 6).Fig. 6. Maximum-likelihood (ML) phylogenetic tree of 26 species based on complete chloroplast sequences. The bootstrap values are marked at the tree node.Fig 6
To further ascertain the phylogenetic position of Amygdalus, a phylogenetic analysis was carried out among 26 species. The result showed that five groups were divided, including eurosids I, eurosids II, gymnosperms, basal angiosperms, and euasterids I (Fig. 6, Table S4). The phylogenomic relationship analysis suggested the Amygdalus species were grouped together. Furthermore, four Persica species*,* including P. mira, Prunus kansuensis, Prunus ferganensis, and P. persica were clustered into a separate branch, and P. pedunculata, P. triloba, and Prunus tangutica were categorized into a branch, while P. mongolica and Prunus davidiana were clustered a branch, these results are consistent with previous reports [2,17].
Experimental Design, Materials and Methods
3
Young and fresh leaves were obtained from all five species. P. mira samples were collected from Bengga, Tibet; P. pedunculata and P. triloba were collected from Inner Mongolia; P. mongolica and P. communis were collected from the Gansu and Henan Provinces, respectively. Samples were stored at −80 °C until analysis.
Total genomic DNA was extracted using a Plant Genomic DNA Kit (Tiangen, Beijing, China). An Illumina Hiseq X high-throughput platform (Illumina, San Diego, CA, USA) was used for DNA sequencing. Library preparation and sequencing were completed by BGI Genomics (Shenzhen, China).
Chloroplast genome fragments were assembled using SOAPdenovo (http://soap.genomics.org.cn/soapdenovo.html) to obtain contigs and optimized assembly results according to the reference cp genome of Prunus persica (GenBank accession ID NC_014697.1). Spaces between long contigs were modified using Gapcloser (https://sourceforge.net/projects/soapdenovo2/files/GapCloser/) to obtain a complete cp genome.
Genomes were annotated using DOGMA (http://dogma.ccbb.utexas.edu/), and the annotation results were manually modified using Geneious (https://www.geneious.com/). An annotated cp genome map was constructed with Organellar Genome Draw (https://chlorobox.mpimp-golm.mpg.de/OGDraw.htmL).
Genomic GC/AT content, as well as lengths of LSC, SSC, IRa, IRb, intergenic regions, exons, introns, tRNA, and rRNA, were counted in DNASTAR (https://www.dnastar.com/).
CpGAVAS (http://124.17.107.12/0506/cpgavas/analyzer/home) was used to visualize the location and length of IR boundaries and junctions.
Long repeat sequences, including forward, palindromic, reverse, and complement repeats, were identified using REPuter (http://bibiserv.techfak.uni-biele.org.de/reputer/); the length and similarity of these sequences were more than 15 bp and 90 %, respectively. The distribution of simple sequence repeats (SSRs) (mono-, di-, tri-, tetra-, penta-, and hexanucleotide repeats) was predicted using MISA (https://webblast.ipk-gatersleben.de/misa/index.php), with a minimum number of six, four, four, three, three and three, respectively. Compound SSRs were those interrupted by a nonrepetitive nucleotide sequence.
Sequences were aligned using mVISTA (http://genome.lbl.gov/vista/index.shtml) to analyze homology and similarity.
The LSC, SSC, and IR border sequences in Amygdalus cp genomes were compared against those in the cp genomes of P. persica (NC_014697) and Pyrus pyrifolia (NC_015996). The IR-SC boundaries of the cp genomes were visualized in IRscope.
The cp genome sequences of 24 angiosperms and 2 gymnosperms were selected for phylogenetic analysis. Multiple genome sequences were aligned in MAFFT version 7 (https://www.ebi.ac.uk/Tools/msa/mafft/). Pinus thunbergia (NC_001631) and Ginkgo biloba (NC_016986) were chosen as outgroups. The maximum likelihood (ML) phylogenetic tree was constructed based on the complete chloroplast genome sequences using the GTAGAMMA model with a bootstrap test of 1000 replicates in RAxML version 8.2.12 (CIPRES Science Gateway version 3.3, https://www.phylo.org/). Trees were visualized using iTOL (https://itol.embl.de/).
Limitations
None.
Ethics Statement
All authors have read and follow the ethical requirements for publication in Data in Brief and confirming that the current work does not involve human subjects, animal experiments, or any data collected from social media platforms.
CRediT authorship contribution statement
Yixiao Chen: Methodology, Investigation, Writing – original draft. Wenquan Bao: Writing – review & editing. Dun Ao: Formal analysis, Resources. Yue Bai: Formal analysis, Resources. Haiguang Huang: Formal analysis, Resources. Rong Yang: Formal analysis, Resources. Lin Wang: Writing – review & editing. Ta-na Wuyun: Conceptualization.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Yu D.Chinese Academy of Sciences Flora Editorial Committee Flora of China. Volume 381986 Science Press
- 2Duan C.Comparative Analysis and Phylogenomics of Chloroplast Genomes of Amygdalus – Subgenus Amygdalus in China 2020 Northwest University 10.27405/d.cnki.gxbdu.2020.002254 · doi ↗
- 3Li C.Yang J.Yao L.Qin F.Hou G.Chen B.Jin L.Deng J.Shen Y.Characterisation, physicochemical and functional properties of protein isolates from Amygdalus pedunculata Pall seeds Food Chem 311202012588810.1016/j.foodchem.2019.12588831771911 · doi ↗ · pubmed ↗
- 4Jing Z.Cheng J.Guo C.Wang X.Seed traits, nutrient elements and assessment of genetic diversity for almond (Amygdalus spp.) endangered to China as revealed using SRAP markers Biochem. Syst. Ecol.492013515710.1016/j.bse.2013.03.015 · doi ↗
- 5Esfahlan A.J.Jamei R.Esfahlan R.J.The importance of almond (Prunus amygdalus L.) and its by-products Food Chem.120201034936010.1016/j.foodchem.2009.09.063 · doi ↗
- 6Guan F.Wang S.Li R.Peng M.Meng F.Genetic diversity of wild peach (Prunus mira Koehne kov et. kpst) from different altitudes in the tibetan plateau by pollen morphous and RAPD markers Hort Science Horts 49820141017102210.21273/HORTSCI.49.8.1017 Retrieved Dec 28, 2023, from · doi ↗
- 7Zhang J.Chen W.Sun W.Zhou Y.Li X.Zhang J.Fan G.Yin H.Qin J.Yuan Y.Xu W.Prunus mira Koehne in Sichuan, China: recorded history as a medicine and food, modern applications, distribution, and ethnobotanical investigations Front. Pharmacol.13202282671210.3389/fphar.2022.826712 PMC 895967035355707 · doi ↗ · pubmed ↗
- 8Shishkin B.K.Yuzepchuk S.V.Flora of the U.S.S.R. V.101971 Israel Program for Scientific Translations Ltd.Jerusalem