A novel synthetic oleanane triterpenoid, 2‐cyano‐3,12‐dioxoolean‐1,9‐dien‐28‐oic acid, regulates mechanical allodynia by rescuing neuronal cell death and glial cell activation in the spinal cord of resiniferatoxin‐treated rats
Ying‐Yi Lu, Chia‐Yang Lin, Hung‐Pei Tsai, Chih‐Lung Lin, Chieh‐Hsin Wu

TL;DR
A new synthetic compound, CDDO, reduces nerve pain in rats by protecting nerve cells and reducing inflammation in the spinal cord.
Contribution
CDDO is shown to alleviate mechanical allodynia by rescuing neuronal death and inhibiting glial activation in a PHN rat model.
Findings
RTX treatment induces mechanical allodynia, neuronal cell death, and glial cell activation in rats.
CDDO administration blocks RTX-induced mechanical allodynia and rescues neuronal cell death.
CDDO inhibits glial cell activation in the spinal cord of RTX-treated rats.
Abstract
Treating postherpetic neuralgia (PHN), which is characterized with a long‐lasting lancinating mechanical allodynia or hyperalgesia, is a big challenge as it is hard to achieve complete resolution. A synthetic triterpenoid, CDDO (2‐cyano‐3,12‐dioxoolean‐1,9‐dien‐28‐oic acid) can exert pleiotropic effects including anti‐inflammation and neuroprotective activities. Nevertheless, the antinociceptive effect of CDDO and its derivatives remains unknown. Resiniferatoxin (RTX) is easily feasible, and an RTX‐treated rodent model can mimic the PHN‐like symptoms. Therefore, RTX‐treated rats were used to serve as a PHN rats’ model in the study to elucidate whether a synthetic triterpenoid, CDDO, can improve mechanical allodynia in RTX‐treated rats. The antinociceptive effects of CDDO were assessed by behavioral tests, western blotting, and immunohistochemistry. Paw withdrawal mechanical threshold…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
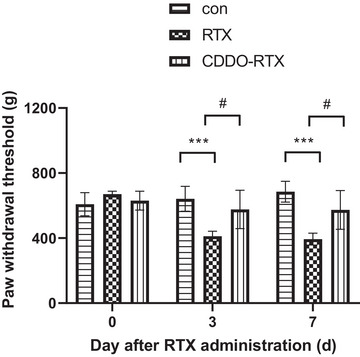 FIGURE 1
FIGURE 1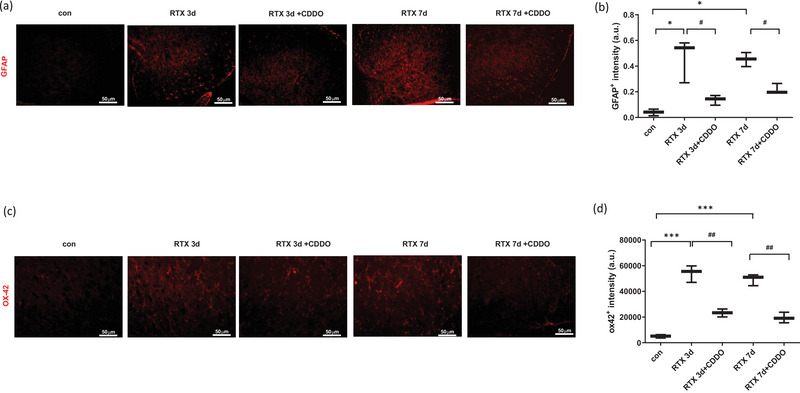 FIGURE 2
FIGURE 2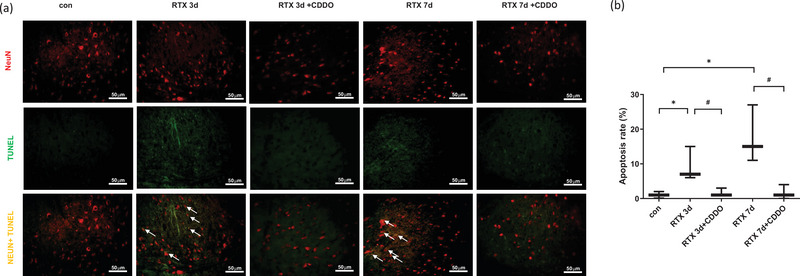 FIGURE 3
FIGURE 3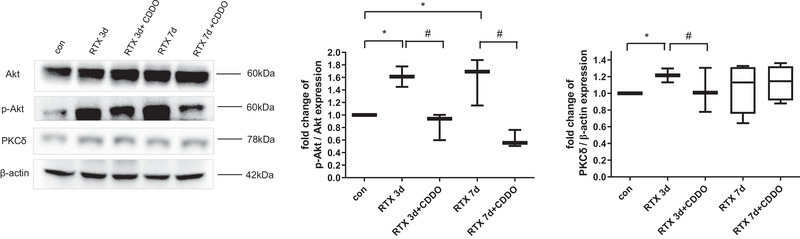 FIGURE 4
FIGURE 4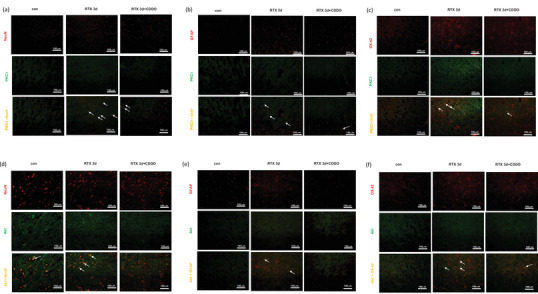 FIGURE 5
FIGURE 5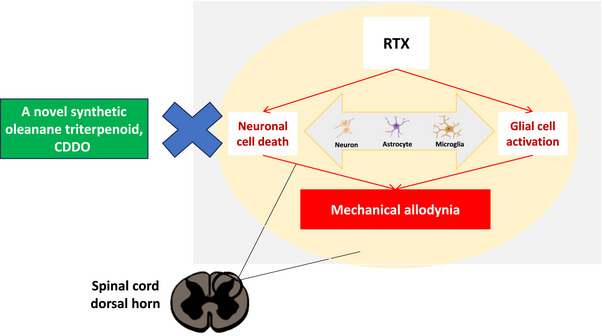 FIGURE 6
FIGURE 6- —Kaohsiung Medical University Hospital
- —Kaohsiung Medical University Research Center Grant
- —Taiwan National Science and Technology Council
- —Kaohsiung Veterans General Hospital 10.13039/501100011913
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsNatural product bioactivities and synthesis · Pharmacological Effects of Natural Compounds · Pain Mechanisms and Treatments
INTRODUCTION
1
Reactivation of latent varicella zoster virus in sensory ganglia leads to grouped cutaneous blisters and neuralgia along the corresponding sensory nerves (Schmader, 2018). After the skin eruption resolved, postherpetic neuralgia (PHN) characterized with a lancinating mechanical allodynia or hyperalgesia could last beyond 3 or more months (Zorzoli et al., 2018). The disproportionate painful sensation without corresponding tissue damage is its characteristic (Oaklander, 2008). Increasing with growing age, the incidence of PHN even rises to 75% in people aged 70 (Delaney et al., 2009). The deficiency of myelin and axon, dorsal horn atrophy, and change of epidermal axon density contribute to the peripheral and central sensitization after receiving nociceptive stimulus (Chen et al., 2017; Watson et al., 1991; Werner et al., 2017). Treating PHN is a big challenge as it is hard to achieve complete resolution even after multimodal therapy and it often impairs life quality due to several complications (Gruver & Guthmiller, 2023; Schutzer‐Weissmann & Farquhar‐Smith, 2017).
As the chemical structure of triterpenoids resembles those of steroids, triterpenoids’ extracts exhibit anti‐inflammatory, antioxidant, antimicrobial, antidiabetic, as well as cardio‐ and hepatoprotective activity (Phillips et al., 2006). To augment the natural triterpenoids’ biological functions, 2‐cyano‐3,12‐dioxoolean‐1,9‐dien‐28‐oic acid (CDDO) and its derivatives were synthesized to amply their pleiotropic capability depending on different doses (Borella et al., 2019; Robles et al., 2016). They exhibit anti‐inflammatory and antioxidative effects at low doses, induce cell differentiation at intermediate doses, lead to anti‐proliferative, cytotoxic, and proapoptotic activities at high doses (Borella et al., 2019). Several studies have documented that CDDO and its derivatives have neuroprotective effects. They can exert cytoprotective properties in neurons, astrocytes, and microglia by activating NADPH quinone oxidoreductase 1 and reducing nitric oxide synthesis (Graber et al., 2011). Activation of glial cells and neuronal cell death contribute to neuropathic pain (Hains & Waxman, 2006; Ohtori et al., 2004). CDDO and a derivative with a methyl ester at C‐28 can reduce microglial activities and preserve neuronal cells survival (Suh et al., 1999; Tran et al., 2008). Nevertheless, the antinociceptive effect of CDDO and its derivatives remains unknown.
Resiniferatoxin (RTX) is a capsaicin ultrapotent analog, which can bind to transient receptor potential vanilloid 1 (TRPV1) irreversibly. It can lead to unmyelinated afferent neurons degeneration, myelinated afferent fibers damage, abnormal nerve sprouting into the lamina II of the spinal dorsal horn, and denervation of skin in the same affected dermatome (Hsieh et al., 2008). It generates paradoxical changes in mechanical and thermal sensitivities, mimicking the distinctive PHN symptoms (Pan et al., 2003; Yuan et al., 2017). Therefore, series studies have showed that systemic RTX acts as a nonviral PHN model to investigate the action of nociceptive afferents (Hong et al., 2018; Lei et al., 2016; Wu et al., 2021). As RTX is easily feasible and an RTX‐treated rodent model can mimic the PHN‐like symptoms, RTX‐treated rats were utilized to serve as a PHN rats’ model in the study. We hypothesized that a synthetic triterpenoid, CDDO, can improve mechanical allodynia in RTX‐treated rats. Later, the life of neurons and activity of glial cells were altered in the spinal cord. This study indicates that a synthetic triterpenoid, CDDO, can interfere neuronal cell death and glial cell activation in the spinal cord of RTX‐treated rats, which offers a potential choice to treat the debilitating PHN.
MATERIALS AND METHODS
2
Animals
2.1
In the experiment, adult, male, Sprague‐Dawley rats (weight of 300–350 g) were used. These rats were purchased from National Animal Center of Taiwan and kept in different cages under a controlled environment (room temperature maintained at 20–24°C) with easily accessible food and water in a 12:12‐h light‐dark cycle. All procedures in the experiment were confirmed and approved by the Animal Care and Use Committee of Kaohsiung Medical University.
Establishment of PHN rat model
2.2
The PHN rats’ model was established following the procedures described by Hsieh et al., and our previous experiment, which a single dose of RTX (50 ug/kg, Sigma), was administrated intraperitoneally (i.p.) (Hsieh et al., 2012, 2008; Lin et al., 2013; Wu et al., 2021). CDDO was dissolved in the DMSO to reach 50 mg/mL at concentration before use. After RTX used, CDDO‐RTX‐rats received CDDO (10 mg/kg, i.p.) every day. Control rats did not receive both RTX and CDDO. The investigators were unaware of the treatment conditions.
Behavioral tests
2.3
The withdrawal thresholds of hind‐paw in response to the force applied by calibrated forceps (Biosebs) were used to estimate mechanical allodynia. An abrupt withdrawal of hind paw was regarded to be a positive response. The measurements were performed before, at the third day and seventh days after treatment with CDDO. The investigators were unaware of the treatment conditions.
Western blotting
2.4
After rats were terminally anesthetized using Zoletil (50 mg/kg), the fourth to fifth lumbar segments of spinal cord were quickly removed and homogenized in lysis buffer containing proteinase inhibitor. To collect the supernatant, the homogenates were centrifuged at 4°C for 30 min at 13,000 rpm. After the protein concentration was estimated by a BCA Protein Assay Kit, proteins were loaded onto 8%–12% SDS‐polyacrylamide gels. The proteins were transferred onto a polyvinylidene fluoride membrane. Membranes were blocked with 5% skim milk for 1 h and incubated at 4°C overnight with following primary antibody: Akt (1: 500; Cell Signaling Technology; #9272; RRID: AB_329827), PKC‐δ (1: 500; BD; 610398; RRID:AB_397781), phospho‐Akt (Ser473) (1: 500; Cell Signaling Technology; #9271; RRID:AB_329825), and β‐actin (1: 10,000; Millipore; MAB1501R; RRID:AB_2223041). After washed with Tween 20 and Tris‐buffered saline, membranes were incubated at room temperature for 1 h with horseradish peroxidase‐conjugated secondary antibody. The immunoreactive bands were visualized by an ECL reagent and measured by MiniChemi chemiluminescent imaging and analysis system (Sage Creation Science). The intensity of bands was normalized to the loading control (β‐actin).
Immunohistochemistry
2.5
After rats were terminally anesthetized using Zoletil (50 mg/kg), the fourth to fifth lumbar segments of spinal cord were removed, postfixed in 4% paraformaldehyde and cut into 8 μm‐thick sections in a cryostat. After rising with phosphate‐buffered saline, blocked with 1% normal goat serum in 0.1% Triton X‐100 and 0.1% phosphate‐buffered saline at room temperature for 1 h, these sections were stained with following antibodies at 4°C for 24 h: glial fibrillary acidic protein (GFAP) (1:400; Sigma; G3893; RRID: AB_477010), NeuN (1:400; Millipore; MAB377; RRID:AB_2298772), OX‐42 (1:100; BIO‐RAD; MCA275G; RRID:AB_321301), phospho‐Akt (Ser473) (p‐Akt) (1:200; Cell Signaling Technology; #9271; RRID:AB_329825), and PKC‐δ (1:200; BD; 610398; RRID:AB_397781) for immunofluorescence. These stained sections were labeled with a mixture of Alexa Fluor 594‐conjugated anti‐mouse secondary antibody (1:500; Jackson ImmunoResearch; 115‐585‐146; RRID: AB_2338881) or Alexa Fluor 488‐conjugated goat anti‐rabbit secondary antibody (1:500; Jackson ImmunoResearch; 111‐545‐144; RRID: AB_2338052) for 1 h. TUNEL kit (Roche; 11684795910) was used to stain apoptotic cells. After washed with phosphate‐buffered saline, these stained sections were covered onto slides and examined using an Olympus fluorescence microscope. ImageJ software was used to measure the amounts of apoptotic neurons, the GFAP intensity, and the OX‐42 intensity.
Statistically analysis
2.6
All data in the experiment was recorded as the mean ± SEM. The differences were analyzed using Student t test if only two datasets or using ANOVA test followed by a Bonferroni test if multiple datasets. Besides, a p value less than .05 was regarded as statistically significant.
RESULTS
3
Effect of a synthetic triterpenoid, CDDO, on mechanical allodynia in RTX‐treated rats
3.1
To assess the effect of CDDO on mechanical allodynia in RTX‐treated rats, we evaluated the paw withdrawal threshold by using calibrated forceps. The baseline mechanical threshold of every group was similar. RTX significantly decreased the mechanical threshold (p < .001), whereas treatment with CDDO significantly increased the mechanical threshold until 7 days later (p < .05) (Figure 1).
*Effect of a synthetic triterpenoid, 2‐cyano‐3,12‐dioxoolean‐1,9‐dien‐28‐oic acid (CDDO), on mechanical allodynia in resiniferatoxin (RTX)‐treated rats. Mechanical threshold was evaluated through paw withdrawal threshold before, at the third day and seventh days after treatment with CDDO in RTX‐treated rats. CDDO significantly increased the mechanical threshold until 7 days later. (n = 6) Mean ± SEM is shown for data (**p < .001 vs. control (con) group, #p < .05 vs. CDDO‐RTX group).
Effect of a synthetic triterpenoid, CDDO, on glial cell activation in RTX‐treated rats
3.2
To assess the effect of CDDO on glial cell activation in RTX‐treated rats, we evaluated the intensity of GFAP (astrocyte marker) and the intensity of OX‐42 (microglia marker) in the spinal cord dorsal horn by using immunohistochemistry. RTX significantly increased the GFAP intensity (p < .05), whereas treatment with CDDO significantly decreased the GFAP intensity until 7 days later (p < .05) (Figure 2a,b). Besides, RTX significantly increased the OX‐42 intensity (p < .001), whereas treatment with CDDO significantly decreased the OX‐42 intensity until 7 days later (p < .01) (Figure 2c,d).
*Effect of a synthetic triterpenoid, 2‐cyano‐3,12‐dioxoolean‐1,9‐dien‐28‐oic acid (CDDO), on glial cell activation in resiniferatoxin (RTX)‐treated rats. Immunohistochemistry study was undergone to stain the sections of RTX‐treated rats’ spinal cord. (a) Representative images for GFAP intensity. GFAP, red staining. (b) Quantification of GFAP intensity. (c) Representative images for OX‐42 intensity. OX‐42, red staining. (d) Quantification of OX‐42 intensity. Scale bar, 50 μm. Mean ± SEM is shown for data (n = 3) (*p < .05, **p < .01, **p < .001 vs. control (con) group, #p < .05, ##p < .01 vs. CDDO‐RTX group).
Effect of a synthetic triterpenoid, CDDO, on neuronal cell death in RTX‐treated rats
3.3
To assess the effect of CDDO on neuronal cell death in RTX‐treated rats, we evaluated the merged intensity of NeuN (neuron marker) and TUNEL in the spinal cord dorsal horn by using immunohistochemistry. RTX significantly increased the apoptotic rate of neurons (p < .05), whereas treatment with CDDO significantly decreased the apoptotic rate of neurons until 7 days later (p < .05) (Figure 3a,b). In summary, CDDO attenuated mechanical allodynia, reduced glial cell activation and neuronal cell death in the spinal cord of RTX‐treated rats.
*Effect of a synthetic triterpenoid, 2‐cyano‐3,12‐dioxoolean‐1,9‐dien‐28‐oic acid (CDDO), on neuronal cell death in resiniferatoxin (RTX)‐treated rats. Immunohistochemistry and TUNEL studies were undergone to stain the sections of RTX‐treated rats’ spinal cord. (a) Representative images for death of neurons. NeuN, red staining; TUNEL, green staining; merged, yellow staining. (b) Quantification of apoptotic rates. Scale bar, 50 μm. Mean ± SEM is shown for data (n = 3) (p < .05 vs. control (con) group, #p < .05 vs. CDDO‐RTX group).
A synthetic triterpenoid, CDDO, regulated PKC‐δ/p‐Akt level in the spinal cord of RTX‐treated rats
3.4
Next, western blotting was performed to estimate whether PKC‐δ/Akt signaling was involved in regulating mechanical allodynia. RTX significantly increased the protein level of PKC‐δ (p < .05), whereas treatment with CDDO significantly decreased the level until 3 days later (p < .05) (Figure 4). Besides, RTX significantly increased the protein level of p‐Akt (p < .05), whereas treatment with CDDO significantly decreased the level until 7 days later (p < .05) (Figure 4). Furthermore, immunohistochemistry study was utilized to clarify cellular localization regulated by PKC‐δ/Akt signaling. PKC‐δ (Figure 5a–c) and p‐Akt (Figure 5d–f) were expressed on neurons, astrocytes, and microglia of spinal cord. Through PKC‐δ/Akt signaling, CDDO can regulate neuronal cell death and glial cell activation in the spinal cord of RTX‐treated rats.
*A synthetic triterpenoid, 2‐cyano‐3,12‐dioxoolean‐1,9‐dien‐28‐oic acid (CDDO), regulated PKC‐δ/phosphorylated Akt (p‐Akt) level in the spinal cord of resiniferatoxin (RTX)‐treated rats. The PKC‐δ, Akt, and p‐Akt protein levels in the spinal cord of RTX‐treated rats were measured by western blotting. Mean ± SEM is shown for data (n = 3) (p < .05 vs. control (con) group, #p < .05 vs. CDDO‐RTX group).
PKC‐δ and p‐AKT were expressed on neurons, astrocytes, and microglia in the spinal cord of resiniferatoxin (RTX)‐treated rats. Immunohistochemistry study was undergone to stain the sections of RTX‐treated rats’ spinal cord before, at the third day after treatment with 2‐cyano‐3,12‐dioxoolean‐1,9‐dien‐28‐oic acid (CDDO). PKC‐δ was expressed on (a) neurons, (b) astrocytes, and (c) microglia. PKC‐δ, green staining; NeuN, red staining; GFAP, red staining; OX‐42, red staining; merged, yellow staining. Scale bar, 50 μm. p‐AKT were expressed on (d) neurons, (e) astrocytes, and (f) microglia. p‐AKT, green staining; NeuN, red staining; GFAP, red staining; OX‐42, red staining; merged, yellow staining. Scale bar, 50 μm.
DISCUSSION
4
A growing body of evidence showed that a synthetic triterpenoid, CDDO can exert pleiotropic effects including neuroprotective activities (Graber et al., 2011). However, its antinociceptive effect remains unclear. In the present study, we utilized RTX‐treated rats to serve as a PHN rats’ model. Our results demonstrated that CDDO can improve mechanical allodynia by interfering neuronal cell death and glial cell activation in the spinal cord of RTX‐treated rats (Figure 6). Therefore, our study suggested that a novel synthetic oleanane triterpenoid, CDDO, is an alternative therapeutic choice for PHN.
The schematic signaling pathways involved in resiniferatoxin (RTX)‐treated rats after receiving daily 2‐cyano‐3,12‐dioxoolean‐1,9‐dien‐28‐oic acid treatment (CDDO). RTX‐treated rats elicit mechanical allodynia. A novel synthetic triterpenoid, CDDO, can improve mechanical allodynia by interfering neuronal cell death and glial cell activation in the spinal cord of RTX‐treated rats.
After nerve injury, spinal disinhibition activates central sensitization and leads to mechanical allodynia (Cao et al., 2022). The spinal cord dorsal horn is the major center to modulate pain signals, where activated glial cells release extracellular signaling molecules to trigger secondary somatosensory neurons (Chen et al., 2021). Nociception is finally transmitted to the supraspinal brain area to process the pain perception. Being the first relay center to percept noxious stimulus, spinal cord dorsal horn can exhibit excessive metabolism and activate inflammation to regulate PHN and serve as a crucial role (Zhao et al., 2019). In steady circumstance, astrocytes in central nervous system (CNS) provide nutritional and structural support for neurons, whereas microglia behaves as CNS‐resident immune cells to keep homeostasis and maintain neural function (Haydon, 2001; Kierdorf & Prinz, 2017). After nerve injury, astrocytes and microglia undergo glial cell activation with structural and functional transformation (Milligan & Watkins, 2009). Microgliosis in ipsilateral spinal dorsal horn developed rapidly followed by astrogliosis (Kohno et al., 2018). Activated central glia release inflammatory cytokines sensitize nociceptive neurons so that pathological pain persists (Ji et al., 2019). Augmented inflammatory cytokines further activate neuronal cells apoptosis to derive hyperalgesia after loss of inhibitory neurons (Hatch et al., 2018). In RTX‐treated rats, both unmyelinated and myelinated nerve fibers are damaged (Wu et al., 2013). RTX not only activates astrocytes and microglia but also increases TNFα to induce death of neurons, which leads to nociceptive hyperalgesia (Lei et al., 2016; Leo et al., 2017). In RTX‐treated rats, blockade of hepatoma‐derived growth factor (HDGF) attenuates mechanical allodynia by inhibiting spinal cord astrogliosis (Wu et al., 2021). In our study, RTX led to mechanical allodynia by activating microgliosis, astrogliosis as well as neuronal cell death, which is consistent with previous studies. CDDO attenuated mechanical allodynia by reducing glial cell activity and death of neurons in the spinal cord of RTX‐treated rats.
PKC‐δ/Akt signaling is involved in cell death (Basu & Pal, 2010; Jin et al., 2014). Regardless of PI3K, PKC‐δ regulates cancer cells survival by activating Akt (Grossoni et al., 2007) or retinal neuronal cell apoptosis of diabetic rats by activating Akt (Kim et al., 2008). Akt regulates mTOR downstream signals to induce hippocampal neuronal cell death (Liu et al., 2014). In response to noxious stimulus, PKC‐δ/Akt signaling can regulate pain processing in neuropathic pain such as paclitaxel neuropathy and diabetic neuropathy (Chen et al., 2017; He & Wang, 2015; Velazquez et al., 2007). Spared nerve injury activates PKC‐δ/Akt to aggravate neuropahic pain mediating by membrane estrogen receptors (Wright et al., 2019). Through PKC‐δ activation, capsaicin enhances TRPV1 expression to cause thermal hyperalgesia (Cesare et al., 1999). Capsaicin also activates Akt in neuronal cells of spinal cord dorsal horn to induce mechanical allodynia (Sun et al., 2006). After surgery, Akt is upregulated in microglia and neuronal cells of spinal cord in mice to elicit postsurgical pain (Xu et al., 2019). Through decreasing Akt level of spinal cord, blockade of HDGF attenuates mechanical allodynia (Wu et al., 2021). Li et al. (2021) had revealed that RTX caused hyperalgesia by activation of PKC‐δ in microglia. In our study, RTX increased the protein level of PKC‐δ/p‐Akt, whereas treatment with CDDO decreased the level (Figure 4). PKC‐δ and p‐Akt were expressed on neurons, astrocytes, and microglia of spinal cord (Figure 5). Through PKC‐δ/Akt signaling, CDDO can regulate neuronal cell death and glial cell activation in the spinal cord of RTX‐treated rats without PI3‐kinase.
CONCLUSIONS
5
In summary, our results show a novel finding that a synthetic triterpenoid, CDDO, is an alternative therapeutic choice for PHN. CDDO can interfere neuronal cell death and glial cell activation in the spinal cord of RTX‐treated rats.
AUTHOR CONTRIBUTIONS
YingYi Lu: Conceptualization; formal analysis; funding acquisition; investigation; project administration; writing—original draft. Chia‐Yang Lin: Data curation; formal analysis. Hung‐Pei Tsai: Investigation; methodology. Chih‐Lung Lin: Formal analysis. Chieh‐Hsin Wu: Conceptualization; funding acquisition; project administration; supervision; validation; writing—review and editing.
CONFLICT OF INTEREST STATEMENT
The authors declared no conflicts of interest.
CONSENT TO PARTICIPATE
All authors approved to participate.
CONSENT FOR PUBLICATION
All authors approved publication.
PEER REVIEW
The peer review history for this article is available at https://publons.com/publon/10.1002/brb3.3398.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Basu, A. , & Pal, D. (2010). Two faces of protein kinase Cdelta: The contrasting roles of PK Cdelta in cell survival and cell death. The Scientific World Journal [Electronic Resource], 10, 2272–2284. 10.1100/tsw.2010.214 21103796 PMC 5763792 · doi ↗ · pubmed ↗
- 2Borella, R. , Forti, L. , Gibellini, L. , De Gaetano, A. , De Biasi, S. , Nasi, M. , & Pinti, M. (2019). Synthesis and anticancer activity of CDDO and CDDO‐Me, two derivatives of natural triterpenoids. Molecules (Basel, Switzerland), 24(22), 4097. 10.3390/molecules 24224097 31766211 PMC 6891335 · doi ↗ · pubmed ↗
- 3Cao, B. , Scherrer, G. , & Chen, L. (2022). Spinal cord retinoic acid receptor signaling gates mechanical hypersensitivity in neuropathic pain. Neuron, 110(24), 4108–4124. e 6. 10.1016/j.neuron.2022.09.027 36223767 PMC 9789181 · doi ↗ · pubmed ↗
- 4Cesare, P. , Dekker, L. V. , Sardini, A. , Parker, P. J. , & Mc Naughton, P. A. (1999). Specific involvement of PKC‐epsilon in sensitization of the neuronal response to painful heat. Neuron, 23(3), 617–624. 10.1016/s 0896-6273(00)80813-2 10433272 · doi ↗ · pubmed ↗
- 5Chen, F. , Chen, F. , Shang, Z. , Shui, Y. , Wu, G. , Liu, C. , & Li, Y. (2017). White matter microstructure degenerates in patients with postherpetic neuralgia. Neuroscience Letters, 656, 152–157. 10.1016/j.neulet.2017.07.023 28729077 · doi ↗ · pubmed ↗
- 6Chen, P. , Wang, C. , Ren, Y. N. , Ye, Z. J. , Jiang, C. , & Wu, Z. B. (2021). Alterations in the gut microbiota and metabolite profiles in the context of neuropathic pain. Molecular Brain, 14(1), 50. 10.1186/s 13041-021-00765-y 33750430 PMC 7941960 · doi ↗ · pubmed ↗
- 7Chen, S. P. , Zhou, Y. Q. , Liu, D. Q. , Zhang, W. , Manyande, A. , Guan, X. H. , & Omar, D. M. (2017). PI 3K/Akt pathway: A potential therapeutic target for chronic pain. Current Pharmaceutical Design, 23(12), 1860–1868. 10.2174/1381612823666170210150147 28190392 · doi ↗ · pubmed ↗
- 8Delaney, A. , Colvin, L. A. , Fallon, M. T. , Dalziel, R. G. , Mitchell, R. , & Fleetwood‐Walker, S. M. (2009). Postherpetic neuralgia: From preclinical models to the clinic. Neurotherapeutics, 6(4), 630–637. 10.1016/j.nurt.2009.07.005 19789068 PMC 5084285 · doi ↗ · pubmed ↗