Picosecond Dynamics of a Small Molecule in Its Bound State with an Intrinsically Disordered Protein
Gabriella T. Heller, Vaibhav Kumar Shukla, Angelo Miguel Figueiredo, D. Flemming Hansen

TL;DR
The study shows that a small molecule can dynamically interact with a disordered protein from the hepatitis C virus, challenging the idea that such proteins are hard to target with drugs.
Contribution
The novel contribution is the experimental demonstration of a small molecule binding to an intrinsically disordered protein with measurable dynamics.
Findings
5-fluoroindole interacts with the disordered domains of non-structural protein 5A from hepatitis C virus with a Kd of 260 ± 110 μM.
The rotational correlation time increased from 27.0 ± 1.3 ps in the free state to 46 ± 10 ps in the bound state.
Small molecules can bind to intrinsically disordered proteins in dynamic ways, unlike rigid binding to structured proteins.
Abstract
Intrinsically disordered proteins (IDPs) are highly dynamic biomolecules that rapidly interconvert among many structural conformations. These dynamic biomolecules are involved in cancers, neurodegeneration, cardiovascular illnesses, and viral infections. Despite their enormous therapeutic potential, IDPs have generally been considered undruggable because of their lack of classical long-lived binding pockets for small molecules. Currently, only a few instances are known where small molecules have been observed to interact with IDPs, and this situation is further exacerbated by the limited sensitivity of experimental techniques to detect such binding events. Here, using experimental nuclear magnetic resonance (NMR) spectroscopy 19F transverse spin-relaxation measurements, we discovered that a small molecule, 5-fluoroindole, interacts with the disordered domains of non-structural protein…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
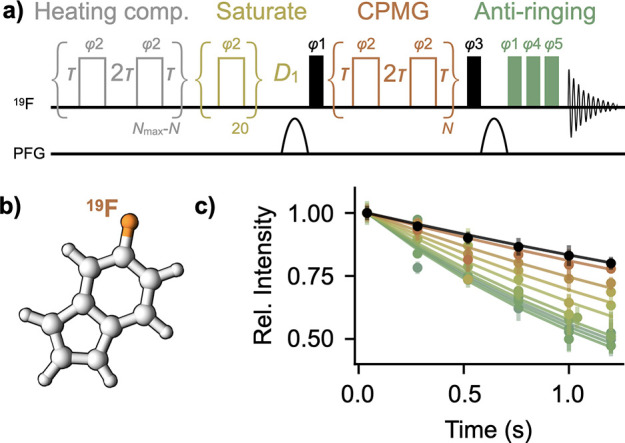 Figure 1
Figure 1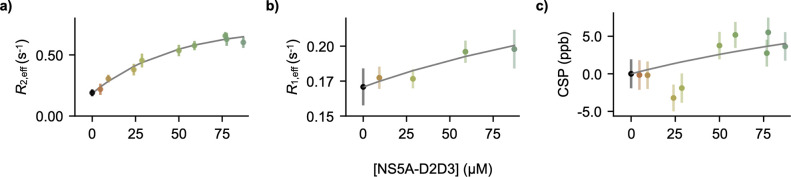 Figure 2
Figure 2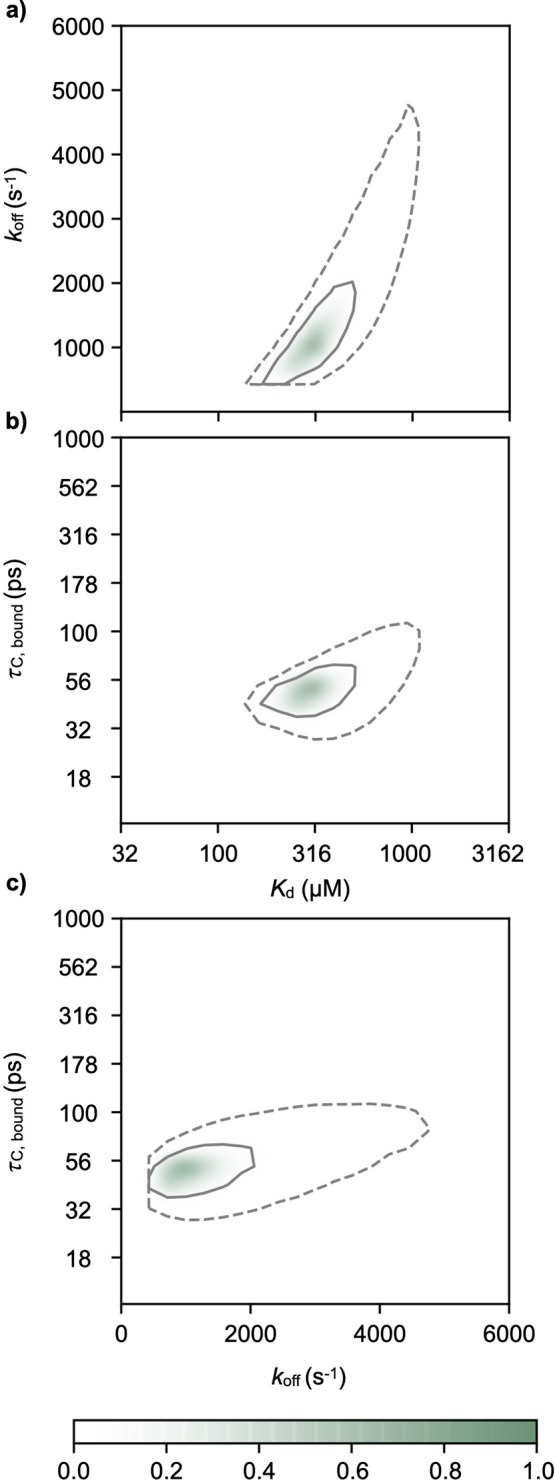 Figure 3
Figure 3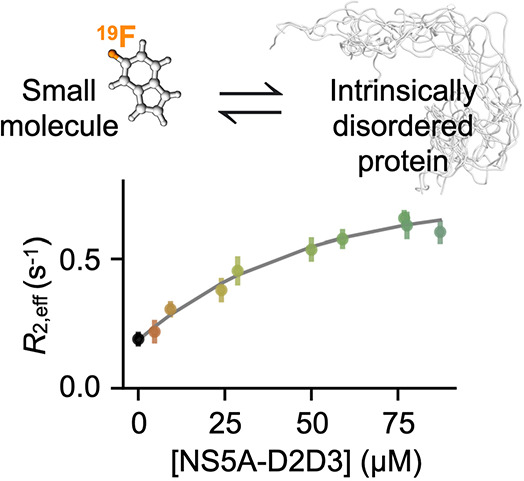 Figure 4
Figure 4- —Engineering and Physical Sciences Research Council10.13039/501100000266
- —Schmidt Science FellowsNA
- —University College London10.13039/501100000765
- —Newnham College, University of Cambridge10.13039/501100000663
- —Biotechnology and Biological Sciences Research Council10.13039/501100000268
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsFrench Historical and Cultural Studies · European history and politics · Historical Studies and Socio-cultural Analysis
One-third of human proteins are intrinsically disordered proteins (IDPs) that rapidly interconvert among different structures.^3−5^ Some IDPs have been suggested to interact with small molecules,^6−12^ thus opening up an enormous class of potential drug targets, especially for conditions including cancers, cardiovascular diseases, type II diabetes, and viral infections.^13^ Nevertheless, the biophysical mechanisms that underpin the binding between small molecules and IDPs are largely based on theoretical molecular dynamics (MD) simulations that, in turn, have suggested that these interactions are highly dynamic.^6,7,9,14−16^ Here, we employ experimental ^19^F-based nuclear magnetic resonance (NMR) spectroscopy to demonstrate that a small molecule remains extremely dynamic in its bound state with an IDP, in striking contrast to how most small molecules bind to structured proteins.
MD simulations have the potential to offer important insights into interactions between IDPs and small molecules; however, these calculations suffer from force field inaccuracies,^17−19^ and the reliability of these computationally expensive simulations is further hindered by long time scales required to reach convergence, especially for dynamic systems like IDPs, which undergo motions on varying time scales.^18,19^ There is also a lack of available experimental techniques suitable for characterizing interactions between IDPs and small molecules. Most of our high-resolution understanding of small-molecule/drug interactions with proteins comes from experimental X-ray crystallography, which allows scientists to resolve the atomistic details of the binding interactions, primarily enthalpic contributions including electrostatics, hydrogen bonding, π–π stacking, and hydrophobic effects. Due to their highly dynamic nature, IDPs are generally not amenable to crystallography, and given the high entropic costs associated with folding from a disordered state, it is not yet well-understood whether a small molecule could induce a single, stable structure for such experiments.
NMR spectroscopy uniquely provides atomic-resolution insights into biomolecular interactions in physiological environments,^20−22^ without the need to apply large labels nor localize the molecules on a surface—both of which may alter the structural ensemble and thus the behavior of the IDP. Standard NMR experiments, such as ligand-detected chemical shift perturbations, are commonly employed to screen and assess the binding of small molecules to structured proteins.^23^ However, chemical shifts report on the local environment of the nuclei in question, which, in turn, are averaged over time and over all the molecules in solution. Given the proposed dynamic nature of the interactions between IDPs and small-molecule ligands,^6,7,9,15^ we rationalized that NMR parameters that report on ligand dynamics and exchange might be more sensitive to detect IDP/small-molecule binding than chemical shifts.^12^ To further enhance the sensitivity of the experiment to binding, we employed ^19^F instead of ^1^H as a probe, as ^19^F has the advantage that there are no background signals in the NMR spectra from buffer components nor protein, and these spectra therefore exclusively report on the small molecule in question.
We chose the disordered domains 2 and 3 from the non-structural protein 5A (NS5A-D2D3) from the hepatitis C virus (JFH-1 genotype) as a model system, given the availability of established purification protocols and chemical shift assignments.^24−26^ Using ^19^F transverse spin-relaxation rates R2,eff via a CPMG-based R2 experiment^1,27,28^ (Figure 1a), we identified that 5-fluoroindole (Figure 1b) interacts with NS5A-D2D3. Specifically, as the concentration of NS5A-D2D3 is increased in samples containing 50 μM 5-fluoroindole, the relaxation curves systematically decrease (Figure 1c), corresponding to an increase in the effective transverse relaxation rate of the ^19^F spin in 5-fluoroindole, R2,eff (Figure 2a). Key features of the R2,eff pulse program (Figure 1a) include 1) a heating compensation element to ensure that the sample is exposed to identical RF power independent of the number of CPMG blocks used^1^ and 2) an anti-ringing sequence to remove broad baseline artifacts at the on-resonance ^19^F frequency that result from sub-optimal performance of the ^1^H RF coil that has been detuned for ^19^F experiments.^1,29^ We also measured ^19^F longitudinal relaxation rates R1,eff (Figure 2b) using an inversion recovery experiment^2^ and ^19^F chemical shift perturbations (Figure 2c) of 50 μM 5-fluoroindole in the presence of varying concentrations of NS5A-D2D3 and observed systematic changes.
To gain additional insight into the interaction mechanism of 5-fluoroindole with NS5A-D2D3, a simple one-site binding model was assumed, where 5-fluoroindole can exist in one of two states: a “free” form (F) and a “bound” form (B) interacting with NS5A-D2D3. The binding mechanism is likely dynamic and more complex, but here we simply assume the “bound” form represents an ensemble of states all interacting with NS5A-D2D3. The increase in R1,eff and R2,eff observed with increasing concentrations of NS5A-D2D3 (Figure 2) could arise from either elevated intrinsic relaxation rates of 5-fluoroindole in the bound conformation or an exchange-induced increase in these relaxation rates.^30^ To address this, we took an integrative approach: the experimental ^19^F transverse relaxation, (Figure 2a), ^19^F longitudinal relaxation (Figure 2b), and ^19^F chemical shifts (Figure 2c, Figure S1) were combined with translational diffusion measurements of 50 μM 5-fluoroindole, measured by ^1^H diffusion ordered spectroscopy (DOSY) (Figure S2). ^1^H instead of ^19^F DOSY measurements were employed due to the relatively increased sensitivity of the ^1^H nucleus. ^19^F transverse relaxation, longitudinal relaxation, and chemical shift data involved titration of NS5A-D2D3 up to 90 μM. ^1^H DOSY measurements were performed in the absence and presence of 75 μM NS5A-D2D3. All data were analyzed simultaneously within the one-site binding model. In particular, we related free and bound longitudinal and transverse relaxation rates (R1,F, R1,B, R2,F, and R2,B) with free and bound rotational correlation times (τ_c,F_ and τ_c,B_, respectively) via well-established equations (Supporting Information).^31−34^ The free and bound rotational correlation times, the dissociation rate (koff), the dissociation constant (Kd), and the diffusion coefficient of the bound form (DB) were determined from a least-squares analysis; see the Supporting Information. The diffusion coefficient of the free form of 5-fluoroindole (DF) was determined to be (1.47 ± 0.02) × 10^–9^ m^2^ s^–1^ by fitting the ^1^H DOSY data in the absence of NS5A-D2D3 (Supporting Information and Figure S2). Fits obtained using all data are shown in Figures 2 and 3. The analysis gave a Kd of 260 ± 110 μM (Figure 3a,b), a koff of 800 ± 500 s^–1^ (Figure 3a,c), a τ_c,F_ of 27.0 ± 1.3 ps, a τ_c,B_ of 46 ± 10 ps (Figure 3b,c), and a DB of (1.5 ± 0.6) × 10^–9^ m^2^ s^–1^.
The rotational correlation time of the bound state of the small molecule (τ_c,B_) of 46 ps suggests that the small molecule is extremely dynamic within the bound state. This is particularly evident when considered in the context of the rotational correlation time of typical disordered proteins. Tryptophan residues, which, like 5-fluoroindole, also contain an indole motif, within disordered protein sequences have been reported to have fast rotational correlation components between approximately 100 and 260 ps, with longer components on the low nanoseconds time scale.^35^ In this context, the τ_c,B_ that we observe, faster than the rotational correlation time of tryptophan residues in disordered proteins, suggests that the small molecule remains highly dynamic in the bound state. This extreme dynamics of the small molecule in the bound state is consistent with predictions of other small molecules interacting with IDPs.^6,9,15^
This analysis allows not only for a sensitive detection of small-molecule binding to IDPs but also the quantification of the associated dynamics of the interaction, dissociation constant, and off-rates. The value of the derived Kd is particularly of note, since the micromolar interaction observed here is the same order as often observed for lead compounds in initial drug-screening programs.^11^
To better understand the role of specificity in this interaction, we compared R2,eff measurements of 5-fluoroindole for samples containing similar densities of either PEG-20k or 75 μM NS5A-D2D3, given that the two polymers have roughly the same molecular weight. The decrease in the relaxation profile is more significant for NS5A-D2D3 than for PEG-20k (Figure S3), suggesting that the interaction between the IDP and 5-fluoroindole is distinct from simple crowding.
It has previously been reported that, at high concentrations, both NS5A-D2 and NS5A-D3 form transient secondary structures, potentially due to the formation of multimers.^24^ To confirm that this also occurs for NS5A-D2D3, we performed circular dichroism (CD) measurements and observed a concentration-dependent loss of disorder at and above 100 μM, suggesting transient secondary structure formation (Figures S4 and S5). From the CD experiments a saturation of the more structured state could not be achieved, and therefore an equilibrium constant could not be determined for the multimerization.
Knowing that 5-fluoroindole interacts with NS5A-D2D3, we wondered whether this interaction alters the equilibrium of N55A-D2D3 related to a change in the secondary structure propensity. At NS5A-D2D3 concentrations at or below 75 μM, we observed minimal changes in the presence of 50 μM 5-fluoroindole (Figure S5a,b), consistent with the protein-detected NMR experiments (Figures S6–S8, discussed below); that is, 5-fluoroindole generally does not alter the secondary structure propensity of NS5A-D2D3. At 200 μM NS5A-D2D3, very subtle changes were observed, suggesting that 5-fluoroindole may stabilize the less disordered state adopted by NS5A-D2D3 when the protein is at higher concentrations (Figure S5c). Of particular interest is that the NS5A-D2D3 system at high concentrations provides a way to assess the binding of 5-fluoroindole to a less disordered state. Notably, when chemical shift perturbations of 5-fluoroindole were measured in the presence of 200 μM NS5A-D2D3, larger chemical shift perturbations for both ^1^H (Figure S4b) and ^19^F (Figure S4c) were detected as compared to those detected for NS5A-D2D3 in its more dilute (and thus more disordered) state. This observation coincided with an increase in ^19^F longitudinal (R1,eff) and transverse (R2,eff) relaxation rates of 0.24 ± 0.02 and 1.40 ± 0.11 s^–1^, respectively. These data suggest a further increase in the effective correlation time of 5-fluoroindole when it interacts with the less disordered state of NS5A-D2D3.
NMR chemical shift perturbations are the gold-standard technique for screening and characterizing small-molecule binding to structured proteins. While significant protein-detected chemical shift perturbations have been reported for small-molecule interactions with IDPs,^9−11^ these are often small—a fraction of the peak line widths—or nearly undetectable in cases like the one presented here (Figures S6–S8). The largest protein-detected chemical shift perturbations we observe (more than 1 SD across all residues, each less than 12 ppb), which are consistent across two concentrations of 5-fluoroindole, occur at residues A308, W312, A313, T334, S414, and S448. Furthermore, it has recently been reported that heteronuclear single-quantum coherence spectra (HSQCs) of disordered proteins are highly prone to false-positive characterization of ligand interactions due to artifacts arising from mismatched pH.^36^ In contrast, we report here that ligand-detected ^19^F transverse relaxation measurements are sensitive to small-molecule/IDP binding.
In this case study, we uncovered a micromolar binding affinity between 5-fluoroindole and NS5A-D2D3 in its disordered form, where chemical shift perturbations were minimal. We observe that transverse relaxation (R2) is more sensitive to binding than either longitudinal relaxation (R1) or chemical shift perturbations alone. In the 1D experiments, subtle changes can indeed be observed, but these could have easily been mistaken for mismatched buffers. These include very subtle increases in peak intensities, which can be explained by increases in longitudinal relaxation suggested by our model, combined with short relaxation delays (^1^H: D1 = 1 s, ^19^F: D1 = 0.5 s). We also observe subtle chemical shift perturbations in the 1D experiments. The change in chemical shift upon binding leads to a chemical-exchange contribution to the transverse relaxation but not to longitudinal relaxation, making changes upon binding easier to detect via R2-based methods as compared to R1-based experiments. We do not observe line broadening in the 1D experiments, as they are overshadowed by ^1^H–^19^F scalar couplings. Our integrative analysis, combining transverse and longitudinal relaxation, chemical shifts, and diffusion measurements, suggests that the small molecule remains very dynamic when interacting with NS5A-D2D3 in its disordered state. Our analysis of the NMR data coarsely assumes a singly averaged bound state, whereas MD simulations can frequently resolve multiple conformations within the bound ensemble.^6,7,9,37^ Additional types of NMR measurements could potentially provide further insight into the bound state ensemble.^38^
We anticipate that ^19^F ligand-detected spin-relaxation experiments offer a promising route to characterize the conformational dynamics of small molecules bound to IDPs. Furthermore, this approach could be adapted as a medium-throughput screening strategy to identify small molecules that bind IDPs and other dynamic biomolecules, especially in cases where such interactions may be largely undetectable by other approaches. Using this tool, it is feasible to quantify the binding of numerous small-molecule/IDP interactions, including point mutants. This could facilitate the exploration of crucial inquiries regarding specificity and the nature of these dynamic interactions, ultimately contributing valuable insights into the prospective “druggability” of these dynamic biomolecules.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Overbeck J. H.; Kremer W.; Sprangers R. A suite of 19F based relaxation dispersion experiments to assess biomolecular motions. J. Biomol. NMR 2020, 74 (12), 753–766. 10.1007/s 10858-020-00348-4.32997265 PMC 7701166 · doi ↗ · pubmed ↗
- 2Vold R. L. On the measurement of transverse relaxation rates in complex spin systems. J. Chem. Phys. 1972, 56 (7), 3210–3216. 10.1063/1.1677681. · doi ↗
- 3Dyson H. J.; Wright P. E. Intrinsically unstructured proteins and their functions. Nat. Rev. Mol. Cell Biol. 2005, 6 (3), 197–208. 10.1038/nrm 1589.15738986 · doi ↗ · pubmed ↗
- 4Csizmok V.; Follis A. V.; Kriwacki R. W.; Forman-Kay J. D. Dynamic protein interaction networks and new structural paradigms in signaling. Chem. Rev. 2016, 116 (11), 6424–6462. 10.1021/acs.chemrev.5b 00548.26922996 PMC 5342629 · doi ↗ · pubmed ↗
- 5Tompa P. Intrinsically unstructured proteins. Trends Biochem. Sci. 2002, 27 (10), 527–533. 10.1016/S 0968-0004(02)02169-2.12368089 · doi ↗ · pubmed ↗
- 6Heller G. T.; Aprile F. A.; Michaels T. C.; Limbocker R.; Perni M.; Ruggeri F. S.; Mannini B.; Löhr T.; Bonomi M.; Camilloni C.; et al. Small-molecule sequestration of amyloid-β as a drug discovery strategy for Alzheimer’s disease. Sci. Adv. 2020, 6 (45), eabb 592410.1126/sciadv.abb 5924.33148639 PMC 7673680 · doi ↗ · pubmed ↗
- 7Heller G. T.; Aprile F. A.; Bonomi M.; Camilloni C.; De Simone A.; Vendruscolo M. Sequence specificity in the entropy-driven binding of a small molecule and a disordered peptide. J. Mol. Biol. 2017, 429 (18), 2772–2779. 10.1016/j.jmb.2017.07.016.28743590 · doi ↗ · pubmed ↗
- 8Follis A. V.; Hammoudeh D. I.; Wang H.; Prochownik E. V.; Metallo S. J. Structural rationale for the coupled binding and unfolding of the c-Myc oncoprotein by small molecules. Chem. Biol. 2008, 15 (11), 1149–1155. 10.1016/j.chembiol.2008.09.011.19022175 · doi ↗ · pubmed ↗