The one that takes it all: The essential role of VDAC3 in the redundant control of ABA signaling
José Manuel Ugalde

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
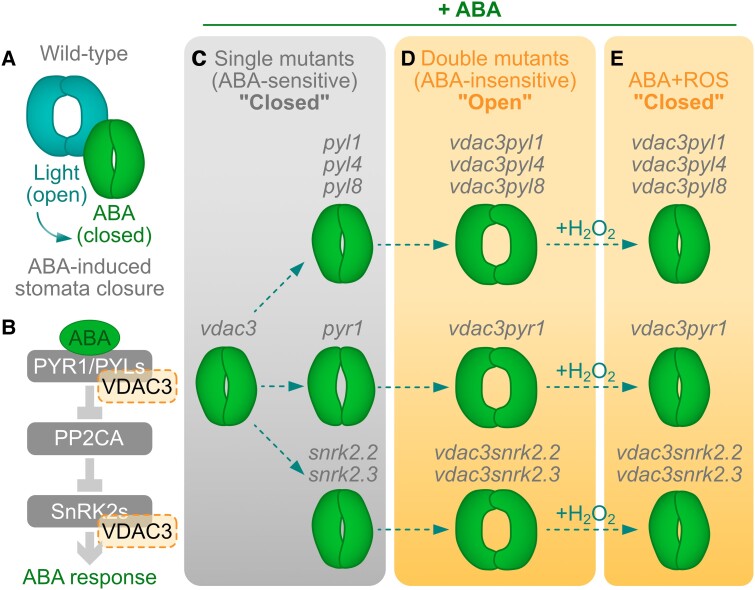 Figure 1
Figure 1- —Deutsche Forschungsgemeinschaft10.13039/501100001659
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsKruppel-like factors research · Caveolin-1 and cellular processes
Gas exchange between plants and their surrounding environment is essential for life on the planet. This gas exchange happens through tiny pores (20–70 µm) on the leaf epidermis called stomata, whose aperture is controlled by a pair of guard cells surrounding the pore (Jezek and Blatt 2017). While stomata opening is crucial for carbon dioxide intake, maintaining an open state exposes the plant to various detrimental effects, such as water loss and a compromised cellular osmotic potential (Ehonen et al. 2019). Controlled stomata closure becomes a necessary adaptation response of plants toward their environment in a process intricately but not exclusively linked to abscisic acid (ABA) and reactive oxygen species (ROS) (Fig. 1A).
In Arabidopsis (Arabidopsis thaliana), the ABA signaling core depends on the ABA-receptors PYRABACTIN RESISTANCE1 (PYR1) receptor and the PYR1-LIKE (PYLs) family. Upon interaction with ABA, the PYR/PYL proteins mediate the inhibition of the PROTEIN PHOSPHATASE 2C A (PP2CA), which, under low ABA levels, acts as a negative regulator of SUCROSE NON-FERMENTING 1 (SNF1)-RELATED PROTEIN KINASES (SnRK2s) (Fig. 1B). The SnRK2s are, in turn, positive regulators of the ABA-dependent signal transduction (Hsu et al. 2021). Notably, these signaling components have a high level of functional redundancy when regulating ABA-dependent stomata closure, where high-order mutants such as the sextuple pyr1pyl12458 are needed to make plants insensitive to ABA-induced stomata closure (Gonzalez-Guzman et al. 2012). Such redundancy is also observed between the kinases SnRK2.2 and SnRK2.3, where only the double mutant between these is partially insensitive to ABA (Fujii et al. 2007).
ABA treatments promote an increment of intracellular ROS levels in guard cells, which have been reported recently to be significant in mitochondria (Postiglione and Muday 2023), an organelle that exhibits important ROS production due to metabolic by-products and fundamental for ABA-mediated stomatal closure (Feitosa-Araujo et al. 2020). In this issue of Plant Physiology, Qin et al. show how VDAC3, a mitochondrial voltage-dependent anion channel protein linked to adaptation responses, is an essential component of the redundant PYR/PYL/SnRK2 signaling core for ABA-mediated stomatal closure (Fig. 1A).
The authors proved that VDAC3 interacts with many ABA receptors, such as PYR1, PYL1, PYL2, PYL4, and PYL8, as well as with the kinases SnRK2.2, SnRK2.3, and 2.6 via yeast 2 hybrid, protein pull down, and bimolecular fluorescence complementation assays. The latter approach also helped them elucidate that all these interactions happen in the plant mitochondria. To test the relevance of VDAC3 in the ABA response, the authors generated double or triple mutants from vdac3 and tested the capacity of these lines to close their stomata upon ABA exposure (Fig. 1A). While the single mutants vdac3, pyl1, pyl4, pyl8, pyr8, snrk2.2, and snrk2.3 did not show any difference in ABA-induced stomata closure, pyr1 and pyl2 lines showed to be partially insensitive to ABA by keeping their stomata not fully closed after treatment (Fig. 1C). Remarkably, crossing each of these single mutant lines with vdac3, every double mutant (vdac3pyr1, vdac3pyl1, vdac3pyl4, vdac3pyl8, vdac3snrk2.2, vdac3snrk2.3) was far less sensitive to ABA, observed as a failure to close their stomata after ABA treatments (Fig. 1D). The ABA-insensitive phenotype in the vdac3 double mutants, also correlated with lower ROS levels after ABA exposure compared to the single mutants. Furthermore, the authors managed to revert the open stomata phenotype by directly treating the double mutants with hydrogen peroxide, indicating that ROS are an essential downstream component of the ABA signaling pathway modulated by VDAC3 (Fig. 1E).
This article unveils VDAC3 as a critical player in the redundancy control of ABA-mediated stomatal closure. The protein interacts with multiple PYLs, PYRs, and SnRK2s, forming a regulatory module crucial for ABA-dependent stomatal closure. The study suggests that VDAC3 acts as a scaffold protein in mitochondria, enhancing the efficiency of ABA-sensing complexes. Yet, future research will establish how conserved is this regulation in other plant species.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ehonen S , Yarmolinsky D, Kollist H, Kangasjärvi J. Reactive oxygen species, photosynthesis, and environment in the regulation of stomata. Antioxid Redox Signal. 2019:30(9):1220–1237. 10.1089/ars.2017.745529237281 · doi ↗ · pubmed ↗
- 2Feitosa-Araujo E , da Fonseca-Pereira P, Pena MM, Medeiros DB, Perez de Souza L, Yoshida T, Weber APM, Araújo WL, Fernie AR, Schwarzländer M, et al Changes in intracellular NAD status affect stomatal development in an abscisic acid-dependent manner. Plant J. 2020:104(5):1149–1168. 10.1111/tpj.1500032996222 · doi ↗ · pubmed ↗
- 3Fujii H , Verslues PE, Zhu J-K. Identification of two protein kinases required for abscisic acid regulation of seed germination, root growth, and gene expression in Arabidopsis. Plant Cell. 2007:19(2):485–494. 10.1105/tpc.106.04853817307925 PMC 1867333 · doi ↗ · pubmed ↗
- 4Gonzalez-Guzman M , Pizzio GA, Antoni R, Vera-Sirera F, Merilo E, Bassel GW, Fernández MA, Holdsworth MJ, Perez-Amador MA, Kollist H, et al Arabidopsis PYR/PYL/RCAR receptors play a major role in quantitative regulation of stomatal aperture and transcriptional response to abscisic acid. Plant Cell. 2012:24(6):2483–2496. 10.1105/tpc.112.09857422739828 PMC 3406898 · doi ↗ · pubmed ↗
- 5Hsu P-K , Dubeaux G, Takahashi Y, Schroeder JI. Signaling mechanisms in abscisic acid-mediated stomatal closure. Plant J. 2021:105(2):307–321. 10.1111/tpj.1506733145840 PMC 7902384 · doi ↗ · pubmed ↗
- 6Jezek M , Blatt MR. The membrane transport system of the guard cell and its integration for stomatal dynamics. Plant Physiol. 2017:174(2):487–519. 10.1104/pp.16.0194928408539 PMC 5462021 · doi ↗ · pubmed ↗
- 7Postiglione AE , Muday GK. Abscisic acid increases hydrogen peroxide in mitochondria to facilitate stomatal closure. Plant Physiol. 2023:192(1):469–487. 10.1093/plphys/kiac 60136573336 PMC 10152677 · doi ↗ · pubmed ↗
- 8Qin H , Yang W, Liu Z, Ouyang Y, Wang X, Duan H, Zhao B, Wang S, Zhang J, Chang Y, et al Arabidopsis mitochondrial protein VDAC 3 ensures redundant control of stomatal closure by ABA signaling components. Plant Physiol. 2023.