Location, location, location: Nuclear pore complexes tether flowering loci to nuclear envelope to boost gene expression
Carlisle Bascom

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
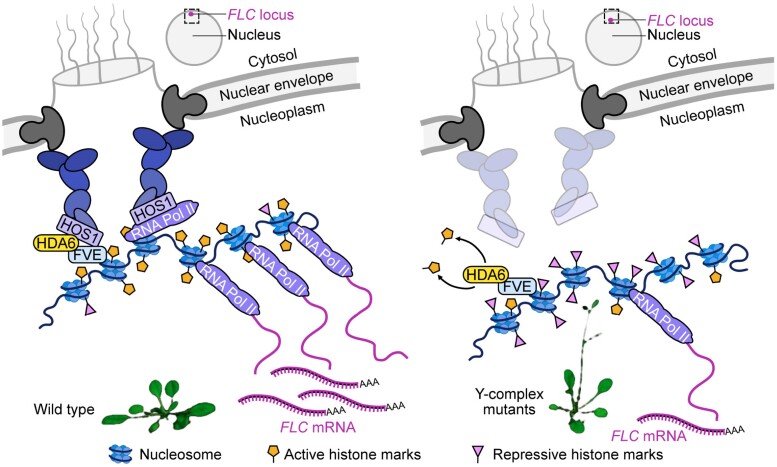 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsNuclear Structure and Function · Plant Molecular Biology Research · Plant Reproductive Biology
In real estate, the three most important factors are location, location, location. But is the same true for a gene locus in the nucleus? Well, research in mammalian cells has discovered that nuclear pore complexes, in addition to regulating transport into and out of the nucleus, affect gene expression by physically tethering the locus to the nuclear envelope (D’Angelo 2018; Brickner et al. 2019). The extent to which plant cells similarly regulate gene expression is understudied. Previous work found that loci of FLOWERING LOCUS C (FLC) shifted to the nuclear periphery in response to temperature, though the exact mechanism was unclear (Rosa et al. 2013). In this issue of The Plant Cell, Penghui Huang and colleagues found that a specific nuclear pore subcomplex, the Y-complex, regulates the expression of FLC in Arabidopsis by physically moving the locus closer to the periphery of the nucleus (Huang et al. 2023) (see Fig. 1).
In Arabidopsis, the Y-complex is made of NUCLEOPORIN43 (Nup43), 85, 96, 107, 133, and 160 proteins, as well as SECRETORY13 (SEC13) and SEC13 HOMOLOG1 (SEH1) (Tamura et al. 2010). Further, HIGH EXPRESSION OF OSMOTICALLY RESPONSIVE GENES1 (HOS1) co-precipitates with Nup43, suggesting HOS1 functions in the Y-complex (Tamura et al. 2010). Curiously, several nup mutants flower earlier than the wild type, including nup85-1, nup96-1, nup107-3, nup160-3, and hos1-3. To unravel the underlying genetic intricacies, the authors conducted RNA-seq experiments with each mutant and found that these mutants displayed remarkably similar expression patterns of differentially expressed genes. Genes in common included the upregulation of the floral activating genes FLOWERING LOCUS T (FT) and SUPPRESSOR OF OVEREXPRESSION OF CO 1 (SOC1) and the downregulation of the floral repressor gene FLC. Histone modification is a well-described mechanism for controlling gene expression. Therefore, the authors used ChIP-qPCR to determine the type of histone modifications within the FLC locus in the background of Y-complex mutants. Overall, the histone modifications lined up with the expression data: there were more repressing modifications on the histones in the FLC locus in the Y-complex mutants than in the wild type. While intriguing, these data did not explain how members of the Y-complex affect histone modifications and thus gene expression.
There are several prerequisites that must be met for a gene to be expressed. Briefly, DNA is wound around histones to form chromatin, and the histones must be modified such that DNA is accessible to RNA polymerase. HISTONE DEACETYLASE 6 (HDA6) is a protein that can deacetylate and methylate histones, thereby augmenting gene expression (Earley et al. 2006). Through a combination of bimolecular fluorescence complementation, co-immunoprecipitation, fluorescent in situ hybridization, and ChIP-qPCR, the authors confirmed that members of the Y-complex not only interact with HDA6 but also with RNA Polymerase II and the histones themselves. With these data, the authors present a compelling model whereby the FLC locus is brought closer to the edge of the nucleus, and this interaction upregulates FLC expression, ensuring that plants flower at the right time. Looking ahead, one cannot help but wonder what factors regulate FLC-Y-complex tethering, such as vernalization, photoperiod, circadian clock control, and stress resilience.
Data availability
This manuscript does not have any data associated with it to make available.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Brickner DG , Randise-Hinchliff C, Corbin ML, Liang JM, Kim S, Sump B, D’Urso A, Kim SH, Satomura A, Schmit H, et al The role of transcription factors and nuclear pore proteins in controlling the spatial organization of the yeast genome. Devel Cell. 2019:49(6):936–947.e 4. 10.1016/j.devcel.2019.05.02331211995 PMC 6613381 · doi ↗ · pubmed ↗
- 2D’Angelo MA . Nuclear pore complexes as hubs for gene regulation. Nucleus. 2018:9(1):142–148. 10.1080/19491034.2017.139554229095096 PMC 5973259 · doi ↗ · pubmed ↗
- 3Earley K , Lawrence RJ, Pontes O, Reuther R, Enciso AJ, Silva M, Neves N, Gross M, Viegas W, Pikaard CS. Erasure of histone acetylation by Arabidopsis HDA 6 mediates large-scale gene silencing in nucleolar dominance. Genes Devel. 2006:20(10):1283–1293. 10.1101/gad.141770616648464 PMC 1472903 · doi ↗ · pubmed ↗
- 4Huang P , Zhang X, Cheng Z, Wang X, Miao Y, Huang G, Fu Y-F, Feng X. The nuclear pore Y-complex functions as a platform for transcriptional regulation of FLOWERING LOCUS C in Arabidopsis. Plant Cell. 2024:36(2):346–366. 10.1093/plcell/koad 271PMC 1082731437877462 · doi ↗ · pubmed ↗
- 5Rosa S , Lucia FD, Mylne JS, Zhu D, Ohmido N, Pendle A, Kato N, Shaw P, Dean C. Physical clustering of FLC alleles during polycomb-mediated epigenetic silencing in vernalization. Genes Devel. 2013:27(17):1845–1850. 10.1101/gad.221713.11324013499 PMC 3778238 · doi ↗ · pubmed ↗
- 6Tamura K , Fukao Y, Iwamoto M, Haraguchi T, Hara-Nishimura I. Identification and characterization of nuclear pore complex components in Arabidopsis thaliana. Plant Cell.2010:22(12):4084–4097. 10.1105/tpc.110.07994721189294 PMC 3027183 · doi ↗ · pubmed ↗